
华北落叶松根际土壤氮素细菌类群和氮素代谢研究
2021-01-02 07:04杨波于志会
江苏农业科学 2021年23期
杨波 于志会




摘要:采用经典的统计学方法和相关性及排序分析,连续3年(2017—2019年)研究不同林龄华北落叶松(幼林、中林、成熟林和过熟林)根际土壤氮素细菌类群和氮素代谢,并探讨了土壤氮素细菌类群和土壤养分含量之间的关系。结果表明:(1)华北落叶松根际土壤pH值显著低于非根际土壤(P<0.05),根际土壤有机碳、全氮和有效磷的含量均显著高于非根际土壤(过熟林除外),根际和非根际土壤有机碳、全氮和有效磷的含量呈一致的变化趋势,随着华北落叶松林龄的增长表现出先增加后降低的趋势,在成熟林阶段达到最大值。(2)华北落叶松根际和非根际物种丰富度指数、均匀度指数和碳源利用指数随着林龄增加呈先增加后降低趋势,在成熟林阶段达到最大值;幼林、中林和成熟林根际丰富度指数、均匀度指数和碳源利用指数显著高于非根际土壤,而过熟林根际与非根际土壤差异不显著。不同林龄根际与非根际土壤微生物优势度指数差异不显著。(3)参与氮素循环的细菌共4类11属,其中固氮细菌有拜叶林克氏菌属、慢生根瘤菌属、根瘤菌属和弗兰克氏菌属,硝化细菌有硝化杆菌属和亚硝化螺菌属,反硝化细菌有假单胞菌属、罗尔斯通菌属、伯克氏菌属、芽孢杆菌属和链霉菌属,氨化细菌有芽孢杆菌属和假单胞菌属。(4)土壤养分含量和微生物多样性指数与固氮细菌相对丰度密切相关,其中土壤pH值对土壤固氮细菌相对丰度贡献为负,土壤养分含量对土壤固氮细菌相对丰度贡献为正,是土壤固氮细菌相对丰度的重要影响因素。(5)冗余分析结果显示,土壤环境因子分别解释了根际和非根际土壤固氮細菌相对丰度的83.38%和80.93%的总特征值,由此表明,土壤环境因子对土壤固氮细菌相对丰度有显著影响。其中对土壤固氮细菌相对丰度影响较大的有有机碳含量和全氮含量,而pH值对土壤固氮细菌相对丰度影响为负。
关键词:华北落叶松;土壤养分;土壤固氮细菌;土壤微生物;多样性指数;冗余分析
中图分类号: Q157 文献标志码: A
文章编号:1002-1302(2021)23-0225-07
收稿日期:2021-06-09
基金项目:吉林省教育厅科学技术项目(编号:JJKH20200388KJ)。
作者简介:杨 波(1979—),男,河北保定人,硕士,副教授,从事生态环境系统理论与技术研究。E-mail:yangboo7904@126.com。
陆地生态的研究视角,不仅具有重要的生态特性,更为关键的是具有特殊性,主要原因在于它是人类生产生活的直接承载者,此外,更是大量动植物生长发育的空间载体[1-2]。毫无疑问其重要性不言而喻,在人类生产生活过程中扮演着极其关键的角色。而其中具有多种生态子系统,其土壤生态等重要性尤为突出,究其原因主要在于大量植被在其生长过程中需要必备的养分及水分[3-4]。而土壤成为了其有效的供应载体,此外,人类粮食作物及蔬菜种植等过程中也离不开土壤的支持[5]。无论是水分还是养分,都受制于土壤状况,尤其是活性和肥力方面,其起着决定性作用[6-8]。正因为如此,居民生活状况受此直接影响较为突出,否则作物难以维持生长;从土壤和植被生长的视角而言,其成为陆地生态系统中尤为关键的环节,其突出效应难以替代,但是对于贫瘠土壤来讲,其在植被生长过程中难以发挥其应有的作用[8-9]。对于土壤质量而言,其影响因素呈现多样性、复杂性并行的特点,其中尤为关键的两大角色是微生物和酶,在微生物的介入之下,大量的有机质能够得以有效降解,而在酶这一关键角色的参与作用下,形成高效的养分转化,进而维持土壤肥力及活性[9-10],为植被生长提供所需能量。大量学者经过实证分析得知,微生物多样性在土壤生态系统保持过程中发挥着关键的作用,如果微生物难以发挥其多样性效应,难以进行高效及时的有机质降解,这将大大抑制土壤活性的提升,对土壤质量产生较大影响。对于土壤而言,根据与根系的位置关系,还可以分为根际土壤与非根际土壤,其中尤为特殊的是根际土壤,主要在于它与植被根系具有更为接近的关系,因此二者之间的关系更为紧密,不仅发挥着土壤的养分及水分供给载体作用,同时深受根系分泌物的影响,且二者具有相辅相成的效应。在根系的作用下,土壤的理化特性发生了明显变化,此外,根系对于水分及养分的吸收状况直接与土壤息息相关,根际与非根际土壤的差异性也较为突出。由于根系区域的特殊性,这不仅是养分聚集区,同时也是微生物的聚集区,在根际微生物的参与之下,养分物质能够高效地转化为植被可吸收利用的养分,加速了植被生长发育,增强了其生长适应性;此外,植被状况也会对微生物生长造成直接的影响效应,对群落分布施加关键作用,温度、湿度等理化特性对群落分布也至关重要。
土壤质量的影响因素不仅具有多样性,更具有复杂性,多种营养元素的组合构成了土壤肥力和活性的关键,其中氮循环能够显著制约土壤肥力,成为其中关键环节[11],且能够以有机结合态的形式广泛存在于土壤之中,且能够在微生物的参与之下进行有效的转化,进而以无机态的形式被植被吸收利用,从而增加植被所需养分[9-10]。大量学者通过实证研究分析得知,当植被分布区域不同的情况下,其土壤养分的时空分布差异也尤为突出,植被和土壤之间的作用机制呈现出明显的复杂性[12-13]。氮循环的有效发挥离不开微生物的参与,无论是硝化反应,还是反硝化反应,均发挥着关键效应,此外氨化作用效果也较为突出[14]。对于森林来说,其中遍布大量的植被以及一系列的生物资源,其中数量庞大的微生物作为土壤生态的关键构成,在有机质分解方面发挥着关键作用,加之酶的参与,对整个土壤养分循环和能量交换发挥着关键影响效应,进而对植被生长发育产生关键影响[10-12]。不同的森林分布使得植被的多样性分布差异尤为突出。对于木兰围场国有林场而言,其中不仅分布着大量的针叶、阔叶林,还包括大量的天然次生林,为了森林资源保护,还进行了大量的人工林建设。为了探究林木资源的植被群落分布,本研究将华北落叶松作为研究对象,立足于华北落叶松人工林的研究视角,从植被及土壤的角度探究其群落分布,并从时间、空间的角度加以分析,以期为森林资源的合理开发利用提供有益参考和借鉴。
1 材料与方法
1.1 研究区概况
本研究将木兰围场国有林场作为研究对象,该林场属河北省围场县所辖,因其处于京津和内蒙古接壤处,因此地势和气候更为复杂多样,山地气候特点较为突出,一方面呈现中温带向寒温带过渡的特点,另一方面半干旱向半湿润过渡的特点也较为突出,但是雨热同期的特点比较明显,根据近年来气象资料得知,研究区的年均温仅为5 ℃左右,最高均温在7月,最高温度在39 ℃左右;无论是降水量还是气温,其季节差异尤为突出,在夏季,具有近600 mm的较高降水量,且主要集中在4—6月,同时具有较高的温度,在冬季,气温较低,且天气干燥,最低温集中在1月,达到了-40 ℃。对于该区域而言,土层一般在50 cm左右,pH值低值一般为6,整体来说具有突出的微碱性或者中性,其肥力并不高,甚至不少区域出现了明显的岩石裸露,其主要的土质不仅包括棕壤、草甸土,还有褐土、风砂土等,整体而言,土层并不厚。该区域不仅分布着大量的落叶林、云杉等乔木资源,还包括大量的山杏等灌木,以及羊胡子草等草本植被,为了森林资源保护,还进行了大量的人工林建设。
1.2 试验设计
1.2.1 样地设置
本研究开始于2017年,为了尽可能地减小试验误差,进行了连续3年的观测研究,从多个角度探究植被与土壤之间的关系。首先选取样地6块,要求长、宽均统一为30 m,在开始指标分析之前对于林木的株高等长势情况进行测量记录。对于土样的采集本研究利用“S”形取样法,要求取样深度达到20 cm,各样地设置样点5个,为降低误差进行5次重复取样,从而共获取土样150个,将其密封后带回实验室,待后续进行指标测量。将土样去除杂质后进行风干处理,之后过筛密封保存,后续开展相关指标测定。
1.2.2 土壤样品的测定
无论是根际土壤还是非根际土壤,均采用四分法进行采样,要求采样深度为40 cm,对各样地采集1 kg土样后均匀混合,置于实验室条件下进行风干处理20 d,之后用元素分析仪开展氮、碳等多种养分指标测定。对速效磷含量、速效钾含量分别利用比色法、光度计法测定;对有机碳含量、pH值、有效磷含量和有效氮含量等指标的测定参考《土壤农化分析》[15]。
1.2.3 土壤微生物群落功能多样性测定
为全面准确测定微生物多样性,首先须要对所用土样进行温育处理,要求处理时间不低于72 h,并借助Biolog-Eco进行测定,然后在NaCl溶液中加入土样,要求其等同于10 g烘干土样,且浓度为0.145 mol/L,并进行振荡处理,10 min后进行读数处理,在恒温培养过程中要求读数频率为每12 h进行1次。待进行稀释处理后再次进行培养,对其吸光度做好核算及记录。本研究过程中借助于平均颜色变化率(AWCD)对其碳源的利用指数(S)加以探究,计算公式[16]如下:
AWCD=∑[(Ci-R)/31];
H=-∑Pi(lnPi);
Pi=(Ci-Ri)/∑(Ci-Ri);
Ds=1-∑Pi;
E=Ds-εPi。
式中:Ci为第i孔的吸光度;R为对照孔的吸光度。对(Ci-R)<0的孔,计算中记为0,即(Ci-R)≥0。Pi为第i孔的相对吸光度与所有微平板的相对吸光度总和的比值;H表示物种丰富度指数;Ds表示优势度指数。
本研究过程中为进一步深入分析微生物,还对其进行了DNA检测,借助于MO BIO试剂盒,对于合格样品进行随机破碎处理,本研究借助于超声波破碎仪,从而形成了一系列的300 bp片段,经过末端修复等一系列处理后形成文库。之后再进行双末端测序,本研究借助于Illumina HiSeq 2000 测序平台,从而获取一系列的有效数据,这也是生物信息分析的基础。所有数据为2017—2019年的平均值。
2 结果与分析
2.1 不同林龄华北落叶松根际与非根际土壤理化性质
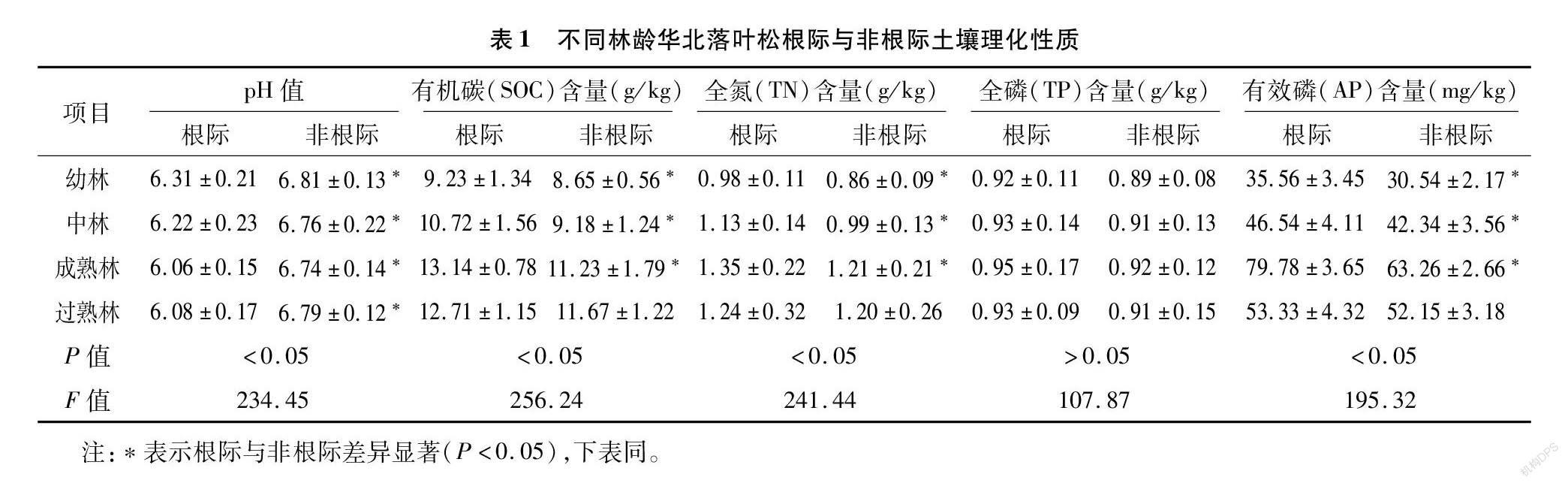
从表1可以看出,华北落叶松根际和非根际土壤pH值随着林龄的增加而先降低后升高,成熟林阶段达到最小值;不同林龄华北落叶松根际土壤pH值显著低于非根际土壤(P<0.05),是因为根在土壤中长期生长,过程中不断向根际分泌出有机化合物,华北落叶松林的土壤pH值处于弱酸性,属较强酸性土壤。华北落叶松根际和非根际土壤有机碳含量随着林龄增加而先增加后降低,在成熟林阶段达到最大值;幼林、中林和成熟林根际土壤有机碳含量显著高于非根际土壤,而过熟林根际土壤有机碳含量与非根际土壤差异不显著。华北落叶松根际和非根际土壤全氮含量随着林龄增加而先增加后降低,在成熟林阶段达到最大值;幼林、中林和成熟林根际土壤全氮含量显著高于非根际土壤,而过熟林根际土壤全氮含量与非根际土壤差异不显著。华北落叶松根际和非根际土壤全磷含量随着林龄增加而先增加后降低,在成熟林阶段达到最大值;不同林龄华北落叶松根际土壤全磷与非根际差异均不显著。华北落叶松根际和非根际土壤有效磷含量随着林龄增加而先增加后降低,在成熟林阶段达到最大值;幼林、中林和成熟林根际土壤有效磷含量显著高于非根际土壤,而过熟林根际土壤有效磷含量与非根际土壤差异不显著。
2.2 华北落叶松人工林土壤微生物群落多样性
由表2可知,华北落叶松根际和非根际物种丰富度指数、均匀度指数和碳源利用指数随着林龄的增加而先增加后降低,在成熟林阶段达到最大值;幼林、中林和成熟林根际土壤的丰富度指数、均匀度指数和碳源利用指数显著高于非根际土壤,而过熟林根际土壤与非根际土壤差异不显著。不同林龄根际与非根际土壤的优势度指数差异不显著。
2.3 华北落叶松人工林土壤氮素微生物类群变化
从表3可以看出,不同林龄华北落叶松人工林土壤中共得到4 类11属氮素循环功能细菌。其中固氮细菌包括拜叶林克氏菌属(Beijerinckia)、慢生根瘤菌属(Bradyrhizobium)、根瘤菌属(Rhizobium)、弗兰克氏菌属(Frankia)共4个属;硝化细菌包括硝化杆菌属(Nitrobacter)、亚硝化螺菌屬(Nitrosospira)共2个属;反硝化细菌包括假单胞菌属(Pseudomonas)、罗尔斯通菌属(Ralstonia)、伯克氏菌属(Burkholderia)、芽孢杆菌属(Bacillus)、链霉菌属(Streptomyces)共5个属;氨化细菌包括芽孢杆菌属、假单胞菌属共2个属。4种功能细菌的相对丰度总体表现为反硝化细菌>氨化细菌>固氮细菌>硝化细菌,其中芽孢杆菌属是优势菌群,占氮素代谢细菌总数的76%以上,亚硝化螺菌属和罗尔斯通菌属所占比例较小,均不到0.1%。固氮细菌和硝化细菌、反硝化细菌和氨化细菌相对丰度随着林龄的增加呈先增加后降低的趋势,在成熟林达到最大值,过熟林阶段有所降低。
2.4 土壤养分与微生物多样性之间相关性
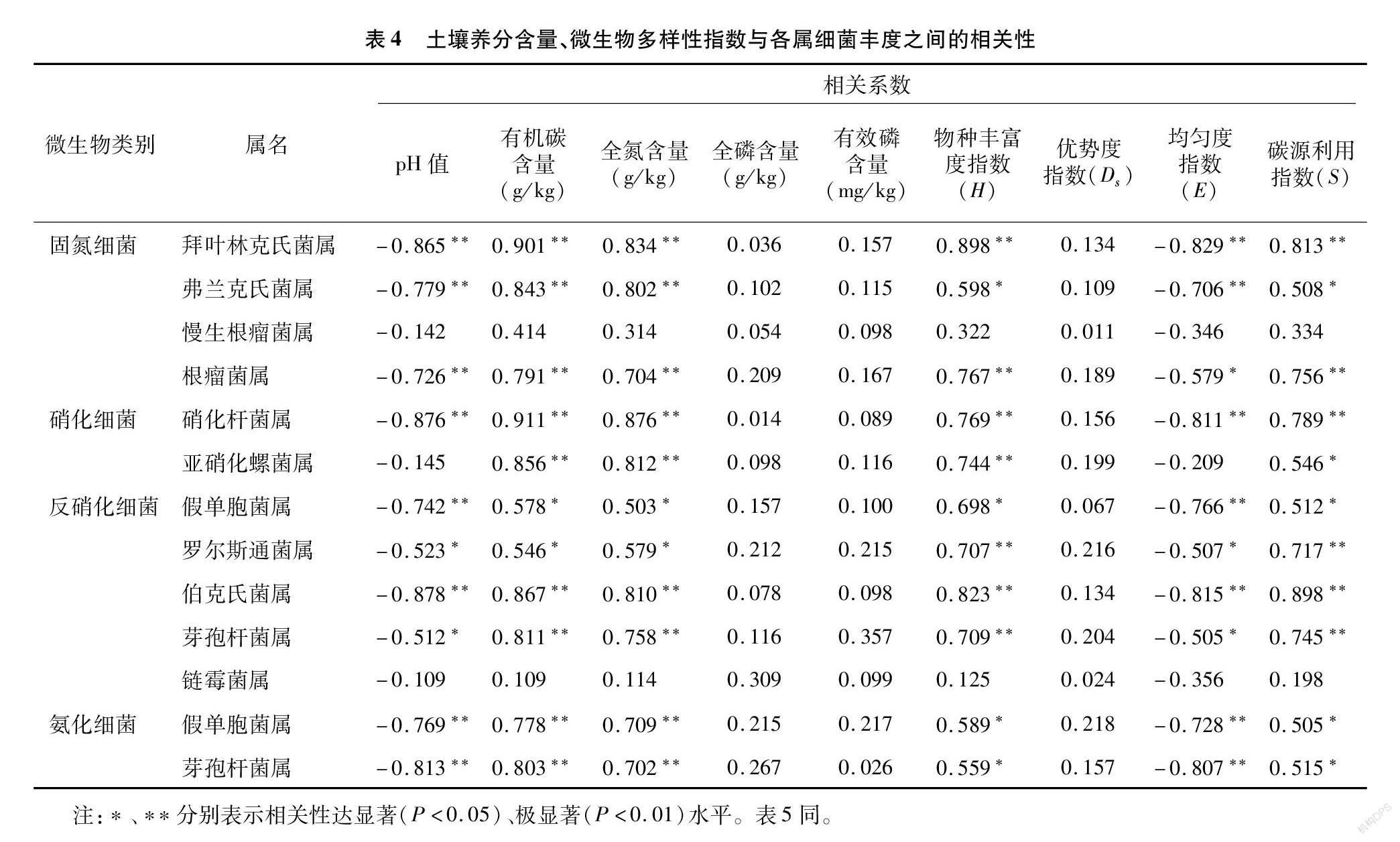
相关性分析结果(表4)表明,土壤pH值与固氮细菌(慢生根瘤菌属除外)、硝化细菌(亚硝化螺菌属除外)、反硝化细菌(链霉菌属除外)和氨化细菌丰度呈显著或极显著负相关关系;土壤有机碳含量、全氮含量与固氮细菌(慢生根瘤菌属除外)、硝化細菌、反硝化细菌(链霉菌属除外)和氨化细菌丰度呈显著或极显著正相关关系;物种丰富度指数(H)、碳源利用指数(S)与固氮细菌(慢生根瘤菌属除外)、硝化细菌、反硝化细菌(链霉菌属除外)和氨化细菌丰度呈显著或极显著正相关关系;均匀度指数(E)与固氮细菌(慢生根瘤菌属除外)、硝化细菌(亚硝化螺菌属除外)、反硝化细菌(链霉菌属除外)和氨化细菌丰度呈显著或极显著负相关;全磷、有效磷含量和优势度指数(Ds)与固氮细菌、硝化细菌、反硝化细菌和氨化细菌丰度没有显著相关性。由此可知,土壤养分含量和微生物多样性指数与固氮细菌相对丰度密切相关,其中土壤pH值对土壤固氮细菌相对丰度贡献为负,土壤养分对土壤固氮细菌相对丰度贡献为正,是土壤固氮细菌相对丰度的重要影响因素。
2.5 土壤微生物群落的冗余分析(RDA)
蒙特卡罗置换检验结果显示,第一典范轴P<0.05,表明该排序模型的解释变量(土壤环境因子)与响应变量(土壤固氮细菌相对丰度)的变化关系较为显著。华北落叶松根际土壤固氮细菌相对丰度与土壤环境因子的冗余分析结果见表5。由图1可知,对于根际,RDA前2个排序轴的特征值分别为0.652 3和0.181 5,分别解释了65.23%和18.15%的固氮细菌相对丰度变化。本研究所选的土壤环境因子共解释了83.38%的总特征值,对华北落叶松根际土壤固氮细菌相对丰度有显著影响。其中对土壤细菌多样性影响较大的有SOC含量和TN含量,而pH值对土壤固氮细菌相对丰度影响为负。对于非根际土壤,RDA前2个排序轴的特征值分别为0.585 6和0.223 7,分别解释了58.56%和22.37%的固氮细菌相对丰度变化。本研究所选的土壤环境因子共解释了80.93%的总特征值,对华北落叶松固氮细菌相对丰度有显著影响。其中对土壤固氮细菌相对丰度影响较大的有SOC含量和TN含量,而pH值对土壤固氮细菌相对丰度影响为负。
3 讨论与结论
微生物分布受制于土壤、气候等一系列影响因素的制约,因此其群落分布也具有突出的差异性,也就形成了不同的多样性分布,无论是均匀度还是丰度等方面均呈现出较大差异。不同的微生物分布状况[17-18]对有机质的分解速率等具有突出差异,形成了突出差异的碳源利用水平,为探究其与土壤等方面的关系,常对其进行多样性分布分析[9,19]。从Shannon指数的角度来说,其侧重于微生物的碳源利用状况,也就是其丰富性方面,该指数越高的情况下,说明其能够越充分地利用不同碳源;对于Mc-Intosh指数而言,更多是侧重于分布均衡性方面,主要受2个方面效应制约,一是微生物种类,二是分布的均匀程度[20]。对于土壤质量而言,影响因素呈现多样性、复杂性并行的特点,其中尤为关键的两大角色是微生物和酶,在微生物的介入之下,大量有机质得以有效降解,而在酶这一关键角色的参与作用下,形成高效的养分转化,进而维持土壤肥力及活性[9-10],为植被生长提供所需能量。通过对华北落叶松连续3年的观测得知,对根际土壤物种而言,无论是丰富度指数、均匀度指数,还是优势度指数、碳源利用指数方面,虽然林木分布具有一定的差异,但是整体而言,其具有较为接近的变化特点,即在林木不断生长的过程中,其多样性指数呈现突出的先升后降变化态势,峰值出现在成熟林;对于非根际土壤来说亦是如此,但是无论是幼林,还是中林甚至是成熟林,非根际土壤的指数均低于根际土壤,且多数差异达到了显著水平。而从过熟林的角度来看,二者差异并不突出,未达到显著水平,主要原因在于该时期林木具有旺盛的根系,能够产生大量的分泌物,在生物量提升方面具有突出效果,而养分又具有明显的富集现象,因此根系区域呈现更高的养分水平[21]。由于林木生长过程中难免产生枯枝落叶,从而形成大量的腐殖质,并在表层土壤形成较为明显的沉积,在微生物的作用下,大量的有机质分解转化为养分,在林木生长发育过程中具有突出的作用[22]。从pH值的角度来讲,无论是根际土壤还是非根际土壤,虽然林木分布具有一定的差异,但是整体而言,其呈现较为接近的变化规律,即随着林木的生长,pH值呈现出突出的先降后升变化态势,低值出现在成熟林;就同一林龄而言,非根际土壤具有更高的pH值,且二者之间的差异尤为突出,达到了0.05水平的显著性差异;主要原因在于根系生长过程中产生了大量的有机化合物,整体来说,该区域土壤呈现突出的弱酸性。
对于微生物而言,在氮循环过程中涉及的细菌共有4类11属,其中不仅有拜叶林克氏菌属等固氮类菌群,还有硝化类菌群,此外,还分布着罗尔斯通菌属等反硝化菌群。从相对丰度来讲,指数最高的是反硝化细菌,其次是氨化细菌,而硝化细菌最低;芽孢杆菌属是其中的优势菌群。虽然菌群分布有突出的差异,但是在林木不断生长的过程中,其丰度呈现出较为突出的先升后降的发展态势,峰值出现在成熟林。
微生物多样性在土壤生态系统保持过程中发挥着关键的作用,如果微生物难以发挥其多样性效应,就难以进行高效及时的有机质降解,这将大大抑制土壤活性的提升,对于其土壤质量产生较大影响。对于土壤而言,根据其与根系的位置关系,还可以分为根际土壤与非根际土壤,其中尤为特殊的是根际土壤,主要在于其与植被根系具有更为接近的关系[22-23]。通过连续3年的观测分析得知,无论是土壤养分、微生物,还是酶活性,其与微生物群落分布的关系较为密切,尤其是土壤养分。为探究多因素交互效应,本研究进行了冗余分析,通过分析得知,除了pH值外,土壤养分含量对微生物分布具有突出的正向影响效果,促进微生物生长发育,尤其是全碳含量及全氮含量成为最主要的影响因子,且相关性最强;综合来看,微生物分布受到一系列因子制约,无论是环境因子还是林木作用,均具有突出影响。
参考文献:
[1]王继伟. 小陇山国家级自然保护区外来引进物种日本落叶松土壤微生物特征研究[D]. 兰州:西北师范大学,2009.
[2]宋妮娜. 大老岭外来种日本落叶松土壤微生物功能多样性及凋落物动态研究[D]. 长春:东北师范大学,2014.
[3]丁 爽. 移栽自不同纬度兴安落叶松土壤微生物量研究[D]. 哈尔滨:东北林业大学,2009.
[4]朱元金,孙 楠. 造林密度对杂种落叶松土壤微生物的影响[J]. 林业科技,2013,38(4):18-21.
[5]闫德仁,刘永军,王晶莹,等. 落叶松人工林土壤肥力与微生物含量的研究[J]. 东北林业大学学报,1996,24(3):46-50.
[6]赵妍丽,孙婧珏,贾淑霞. 施肥对落叶松和水曲柳人工林土壤微生物生物量碳和氮季节变化的影响[C]//第九届中国林业青年学术年会论文集.成都,2010:129.
[7]邵元元,邹 莉,王志英,等. 落叶松人工林土壤养分与微生物群落的变化动态[J]. 东北林业大学学报,2011,39(1):82-84,98.
[8]方 成,岳明灿,王东升,等. 化肥减施配施微生物菌剂对鲜食玉米生长和土壤肥力的影响[J]. 土壤,2020,52(4):743-749.
[9]刘云霞. 氮添加对落叶松人工林土壤微生物活性及其群落结构的影响[D]. 北京:中国科学院研究生院,2010.
[10]贾淑霞,赵妍丽,孙 玥,等. 施肥对落叶松和水曲柳人工林土壤微生物生物量碳和氮季节变化的影响[J]. 应用生态学报,2009,20(9):2063-2071.
[11]王丽红,郭晓冬,谭雪莲,等. 不同轮作方式对马铃薯土壤酶活性及微生物数量的影响[J]. 干旱地区农业研究,2016,34(5):109-113.
[12]陈立新. 落叶松人工林施肥对土壤酶和微生物的影响[J]. 应用生态学报,2004,15(6):1000-1004.
[13]刘延滨,王庆成,王承义,等. 退化落叶松人工林近自然化改造对土壤微生物及养分的影响[J]. 生态学杂志,2012,31(11):2716-2722.
[14]邹 莉,李国江,郑国强. 水曲柳落叶松纯林与混交林土壤微生物主要生理类群[J]. 东北林业大学学报,2000,28(2):49-51.
[15]鲍士旦. 土壤农化分析[M]. 3版. 北京:中国农业出版社,2019.
[16]郭乾坤,梁国庆,周 卫,等. 长期有机培肥提高红壤性水稻土生物学特性及水稻产量的微生物学机制[J]. 植物营养与肥料学报,2020,26(3):492-501.
[17]賈淑霞. 落叶松和水曲柳人工林土壤、根系和土壤微生物呼吸研究[D]. 哈尔滨:东北林业大学,2009.
[18]杨 凯,朱教君,张金鑫,等. 不同林龄落叶松人工林土壤微生物生物量碳氮的季节变化[J]. 生态学报,2009,29(10):5500-5507.
[19]徐 辉. 落叶松与白桦林地土壤微生物边缘效应研究[D]. 哈尔滨:东北林业大学,2017.
[20]耿莹莹. 大兴安岭典型林分土壤微生物生物量动态变化[D]. 哈尔滨:东北林业大学,2012.
[21]管 铭,潘小翠,张崇邦,等. 外来植物入侵的微生物生态学研究进展[J]. 浙江大学学报(理学版),2015,42(4):445-452.
[22]毛 璐,宋 春,徐 敏,等. 栽培模式及施肥对玉米和大豆根际土壤磷素有效性的影响[J]. 中国生态农业学报,2015,23(12):1502-1510.
[23]王桔红,张丽娜,陈学林,等. 入侵植物对根际土壤微生物群落影响的研究进展[J]. 生态科学,2016,35(6):204-210.
猜你喜欢
俪人·教师版(2016年18期)2017-03-02
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
河北渔业(2015年5期)2016-03-01
天津农业科学(2015年12期)2015-12-03
天津农业科学(2015年12期)2015-12-03
湖北农业科学(2015年16期)2015-10-28
安徽农学通报(2015年15期)2015-08-19
