
蛋氨酸限制提高内源性H2S 活性延缓衰老的机制研究进展#
2021-03-08 02:38李欣吴海燕
四川生理科学杂志 2021年1期
李欣 吴海燕
(1.四川大学华西基础医学与法医学院基础医学专业2018级,四川 成都 610041;2.四川大学华西基础医学与法医学院药理学教研室,四川 成都 610041)
衰老,以机体生理完整性的逐渐丧失为特征,造成机体器官功能逐渐受损,导致相关疾病包括癌症、糖尿病、心血管疾病及神经退行性疾病等发生。
在不造成营养不良前提下的饮食限制(Dietary restriction,DR)被证实能够延长多种真核生物包括从酵母、线虫、啮齿类动物到灵长类动物的生命[1]。其中,蛋氨酸限制(Methionine restriction,MetR)是近年来研究较多的DR方案之一。MetR对延缓衰老和延长寿命的作用机制目前存在多种看法,见图1,其一为营养感知信号的下调[2],主要包括沉默信息调节因子2相关酶(Silent information regulator2-related enzymes, SIRT )、 腺苷一磷酸激活的蛋白激酶(AMP-activated protein kinase,AMPK)通路的上调、胰岛素/胰岛素样生长因子1(Insulin-like growth factor 1,IGF-1)及哺乳动物雷帕霉素靶蛋白(Mammalian target of rapamycin,mTOR)通路的下调等[3]。
最近,有学者提出内源性硫化氢(Endogenous hydrogen sulfide, H2S)活性的提高可能是MetR的 主要机制,并且通过上述通路影响衰老进程。
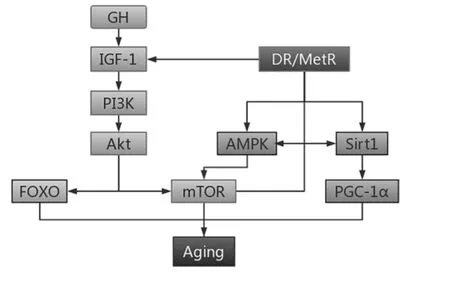
图1 饮食限制通过生长营养轴调节衰老
1 MetR延缓衰老进程
MetR作为含硫氨基酸饮食限制的主要方法,其益处在无脊椎动物实验上已经得到了充分的证实。Ruckenstuhl等人发现,MetR可以通过上游TOR途径或直接作用来增强自噬,从而延长酵母细胞的寿命,而自噬必需基因ATG5、ATG7或ATG8的缺失可以消除MetR的正面作用[4]。另有研究表明,二甲双胍改变了大肠杆菌蛋氨酸的代谢,导致大肠杆菌中蛋氨酸的限制,从而延长了宿主秀丽隐杆线虫(C.elegans.)的寿命,这种作用可以被线虫蛋氨酸合成酶(metr-1)和S-腺苷蛋氨酸合成酶(sams-1)的突变所改变[5]。
MetR的作用在啮齿类动物上也得到了充分的体现。Orentreich等人首次发现如果将饮食中蛋氨酸的含量从0.86%降低到0.17%可以将雄性Fischer 344大鼠的寿命延长30%[6]。在小鼠实验中,减少饮食中80%的蛋氨酸可逆转炎症以及DNA损伤反应基因的转录组改变从而延长早衰综合征(Hutchinson-Gilford ProgeriaSyndrome,HGPS)小鼠模型的寿命[7]。在体外实验中,MetR可通过下调线粒体蛋白合成和呼吸链的组装,抑制细胞衰老延长人二倍体成纤维细胞的寿命[8]
2 转硫通路通过促进蛋氨酸代谢提高内源性H2S 含量
2015年,Hine等人首先发现含硫氨基酸限制可提高转硫通路(Transsulfuration pathway,TSP)中胱硫醚-γ-裂解酶(Cystathionine-γ-lyase,CGL)活性,导致内源性H2S含量升高,从而减缓肝脏缺血再灌注损伤[9]。研究发现长期MetR、隔天禁食及20%-30%DR饲养的成年小鼠的肝脏和肾脏相比对照组产生H2S的能力增强,因而能够抵抗损伤延长寿命。
哺乳动物体内源性H2S主要来源于蛋氨酸和半胱氨酸,由TSP 中的酶胱硫醚-β-合成酶(Cystathionine-β-lyase,CBS)、胱硫醚-γ-裂解酶(Cystathionine-γ-lyase,CGL)和3-巯基丙酮酸硫基转移酶(3-Mercaptopyruvate Sulfurtransferase,3-MST)以磷酸吡哆醛(Pyridoxal phosphate,PLP)为辅因子及半胱氨酸为底物催化产生[10],见图2。在蛋氨酸转化为同型半胱氨酸后,CBS催化同型半胱氨酸和丝氨酸生成胱硫醚,随后胱硫醚在CGL的作用下转化为半胱氨酸,这是H2S生成的关键底物。
半胱氨酸在CBS和CGL的作用下生成H2S,此外,也可通过半胱氨酸转氨酶(Cysteine aminotransferase,CAT)生成3-巯基丙酮酸(3-mercaptopyruvate,3-MP)再经过3-巯基丙酮酸硫基转移酶(3-Mercaptopyruvate Sulfurtransferase,3-MSP)的作用生成H2S。
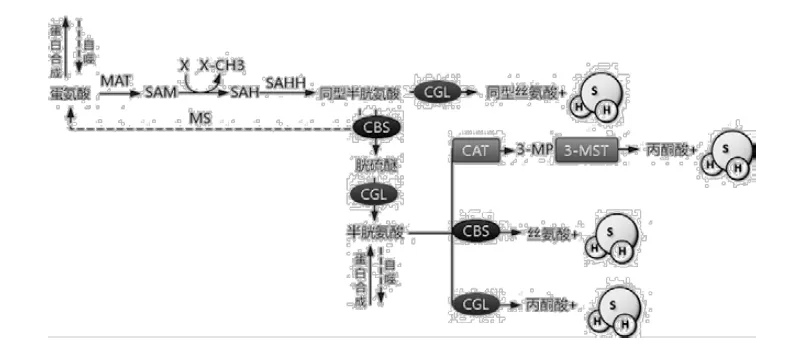
图2 蛋氨酸在哺乳动物体内的代谢
3 MetR 上调H2S 及H2S 的作用机制
在哺乳动物中,限制含硫氨基酸摄入可以从转录水平上调TSP基因。缺乏单一氨基酸可以激活氨基酸饥饿反应激酶——一般性调控阻遏蛋白2激酶(General control non-derepressible 2 kinase,GCN2),导致其靶点eIF2α的磷酸化以及转录因子如激活转录因子4(Activating transcription factor 4,ATF4)的稳定[11]。MetR激活小鼠肝脏中氨基酸饥饿反应。ATF4参与CGL的调控,它不仅在热量限制(Calorie restriction,CR)和MetR的小鼠肝脏中增加,在其他与饮食限制无关的慢性衰老模型中也升高[12]。
TSP中的关键产物是半胱氨酸,当体内半胱氨酸含量较低的时候,机体以蛋氨酸为原料通过转甲基途径(Transmethylation pathway,TMP)和TSP逐步生产半胱氨酸,见图2。当限制H2S生成底物半胱氨酸时,TSP被激活,其中关键酶CGL和CBS的活性上调,从而使H2S产量增加。当限制反应底物(半胱氨酸)时,内源性硫化氢(Endogenous hydrogen sulfide,H2S)的产量反而增高,这似乎有些矛盾。然而,在MetR情况下,由CGL和CBS介导衍生的H2S的具体来源尚未十分清楚。其中,自噬释放的游离半胱氨酸可能是H2S的重要来源,这在酵母身上得到了很好的诠释,酵母MetR的寿命延长作用依赖于自噬的存在[4]。而在小鼠中,当自噬蛋白5(Autophagy protein 5,ATG5)或7(Autophagy protein 7, ATG7)的敲除亦可导致H2S产量的下调[13]。
H2S一直被认为是毒素及环境污染物,高浓度的H2S通过抑制细胞色素C氧化酶和干扰铁依赖的生化反应来抑制呼吸。近年来科学家们发现到H2S可以在生物体内合成,并确定H2S为继一氧化碳(Carbon monoxide,CO)和一氧化氮(Nitric oxide,NO)的之后的第三个气体信号转导分子[14]。在2007年,H2S首次被发现可以延长秀丽隐杆线虫的寿命并且调节其耐热性,50 ppm的H2S使秀丽隐杆线虫的平均寿命增加了70%,在高温(35°C)下秀丽隐杆线虫的存活率也提高了约8倍[15]。在秀丽隐杆线虫中,3-巯基丙酮酸硫基转移酶(3-mercaptopyruvate sulfurtransferase,3-MST)是主要的生成H2S的酶,在3-MST敲除后,野生秀丽隐杆线虫的寿命降低了约36%。
H2S被认为是MetR中发挥作用的主要分子,可以从不同方面延缓衰老。主要分子机制是其对蛋白质表面暴露的半胱氨酸和硫醇残基的硫化作用(Sulfhydration),使得-SH基团转化为-SSH或过硫化物基团。这种翻译后的修饰对靶蛋白功能有着重要作用。由于暴露的半胱氨酸残基易受亚硝基化的影响,H2S可与这些残基竞争从而影响蛋白质的功能。如前所述,衰老通常伴随着营养感知信号失调,而IIS是其中的主要信号通路。H2S的合成酶CGL和CBS可在胰岛β细胞表达,并通过刺激KATP离子通道,抑制胰腺β细胞和胰岛素分泌从而下调IIS通路[16]。此外,最近一项研究显示H2S可以显著下调IGF-1R的表达刺激IGF-1R的硫化作用,减弱IGF-1与IGF-1R的结合从而抑制血管平滑肌细胞的增殖[17],这一研究为增生性心血管疾病(如动脉粥样硬化)提供了创新治疗策略。
4 H2S 供体药物的使用
如前所述,内源性H2S是MetR延缓衰老及相关疾病的关键分子,因而研究者通过化合物模拟H2S在体内的生物学效应。但是因H2S是气体小分子物质,单纯将其用于药物试验的可控性较差,所以现在多使用H2S供体,通过在体内转换成H2S来治疗衰老相关疾病。H2S供体可分为快速释放H2S的药物(如NaHS和Na2S)和缓慢释放H2S的药物(如GYY4137)。目前针对H2S供体的动物研究显示,NaHS通过大鼠心房的KATP通道和PI3K/Akt/NOS/sGC通路刺激心房钠尿肽(Atrial natriuretic peptide,ANP)分泌[18],可能是H2S潜在的保护心血管系统的机制。此外,研究发现GYY4137可以通过激活PI3K/Akt信号,以及部分依赖NO信号和抑制糖原合成酶-3β(Glycogen synthase kinase -3β,GSK-3β)保护心脏从而免受致命的再灌注损伤,因此可作为急性心肌梗死再灌注治疗的辅助手段[19]。最近,一种新型的线粒体靶向的H2S供体药物AP39被报道可通过保护线粒体功能对神经元及阿尔兹海默症小鼠起保护作用[20],为将来神经退行性疾病的治疗提供了治疗策略。体外实验中,AP123和AP39也被证实对微血管内皮细胞的高血糖损伤具有保护作用[21]。
5 总结与展望
MetR和DR通过相同的机制延缓衰老,主要包括IIS,mTOR、AMPK以及sirtuin等通路,这些信号通路之间彼此关联互相影响,MetR通过这些通路以及它们的相互作用下调营养感知信号从而延缓衰老进程,缓解衰老相关疾病。
Hine等人的研究显示,MetR主要通过激活TSP关键酶CGL的活性上调内源性H2S的含量。H2S可以通过多种机制来延缓衰老相关疾病,它同样与营养感知信号通路的关系密切。PTEN是IIS通路中PI3K-Akt轴的主要拮抗剂,H2S通过对PTEN的硫化作用来调节其活性,从而被看作是PTEN的内源性调节剂[22]。除此之外,H2S还可通过对肾小管上皮细胞中AMPK信号通路的激活以及抑制mTOR复合物1活性来延缓肾脏衰老[23]。H2S通过几乎与MetR相同的通路影响营养感知信号的调节,也能从另一个角度说明MetR的主要作用机制在于内源性H2S含量的升高。
已经有许多研究证实H2S在心血管疾病、糖尿病、肿瘤以及神经退行性疾病等衰老相关疾病中的重要作用。因此,针对H2S供体药物作为饮食限制模拟物的开发应用在预防、延缓衰老及其相关疾病中具有重要意义。
目前,H2S供体药物的研究尚处于动物实验阶段,在哺乳动物身上对抗衰老相关疾病取得了瞩目的效果。但是将其正式作为药物应用于人体还需通过谨慎而严密的临床试验。
猜你喜欢
生物信息学(2022年3期)2022-11-12
全科护理(2022年14期)2022-05-16
中国饲料(2022年5期)2022-04-26
中国医药导报(2021年35期)2022-01-20
西南农业学报(2021年10期)2021-12-14
昆明医科大学学报(2021年8期)2021-08-13
猪业科学(2020年12期)2021-01-09
科学(2020年3期)2020-11-26
实用器官移植电子杂志(2020年4期)2020-08-31
心理学报(2020年7期)2020-07-13
