
基于转录组的葡萄花翅小卷蛾三类解毒酶基因家族分析
2021-03-16 13:35白姣洋赵俊凝陈茂华
生物安全学报 2021年1期
白姣洋, 赵俊凝, 王 康, 陈茂华
西北农林科技大学植物保护学院/旱区作物逆境生物学国家重点实验室/农业农村部西北黄土高原作物有害生物综合治理重点实验室,陕西 杨凌712100
葡萄花翅小卷蛾是影响世界葡萄产量和质量的一种重要经济害虫,2012年国家检验检疫中心将其列入检疫性有害生物名录(牛春敬等,2013)。该虫原产于意大利,后在地中海和欧洲地区迅速蔓延,现已扩散至美洲阿根廷、智利、加利福尼亚等地区(Gilliganetal.,2011)。目前,我国尚未出现该虫入侵的相关报道(李俊峰等,2017)。
葡萄花翅小卷蛾隶属鳞翅目Lepidoptera卷蛾科Tortricinae,其幼虫可取食多种经济作物的花和果实,寄主包括葡萄VitisviniferaL.、油橄榄OleaeuropeaL.、迷迭香RosmarinusoficinalisL.、铁线莲ClematisvitalbaL.、山茱萸Cornusspp.、忍冬LoniceraxylosteumL.、女贞LigustrumvulgareL.等在内的40多种植物(Iroiattietal.,2011),对不同寄主植物营养物质和次生代谢物具有广泛适应性。目前葡萄花翅小卷蛾的控制主要依赖化学农药(Akyol & Aslan,2010; Ifoulis & Savopoulou,2004),而杀虫剂的长期使用导致该虫对有机磷、氨基甲酸酯、拟除虫菊酯等多种杀虫剂产生了抗药性,造成该虫防治困难(苏莎等,2020; Pasquinietal.,2018)。
当昆虫遭遇植物防御性次生代谢物和农药等有毒外源化合物刺激时,其体内的解毒酶会发生一系列变化,从而协助昆虫消除这些化合物对其生长发育的不利影响(陈澄宇等,2015; 段辛乐等,2015; 吴有刚等,2019)。Hatipogluetal.(2015)研究发现,采自土耳其的葡萄花翅小卷蛾产生抗药性时,抗性品系与敏感品系虫体内的解毒酶表达量存在差异。Navarro-Roldánetal.(2020)的研究也证实了不同性别葡萄花翅小卷蛾种群体内解毒酶活性不同。昆虫的解毒酶基因家族主要包括细胞色素P450(Cytochrome P450s,P450s)、羧酸酯酶(Carboxylesterases,CarE)和谷胱甘肽S-转移酶(Glutathione S-transferase,GSTs)三大酶系。不同解毒酶对杀虫剂的代谢方式不同:CarE可使有机物的酯键发生断裂,达到解毒效果;GST能促使体内各种有毒物质以非酶方式排出体外。抗药性相关的P450基因主要为CYP4和CYP6家族的成员(张红英等,2002),这些基因在昆虫抗性种群中表达水平显著上升。一般来说,三大解毒酶可通过高表达、基因扩增或基因突变等方式提高昆虫对杀虫剂的抗性。
迄今为止,对葡萄花翅小卷蛾抗性的研究主要集中在比较其对不同杀虫剂的抗性水平,而对抗药性的分子机制知之甚少(Navarro-Roldánetal.,2020),解毒酶相关的研究对该虫的抗药性监测和抗药性治理具有重要的意义。
本研究对葡萄花翅小卷蛾的转录组进行测序,并通过注释信息获取解毒酶基因,与棉铃虫Helicoverpaarmigera(Hübner)和苹果蠹蛾CydiapomonellaL.等鳞翅目昆虫的同源基因对比,进行系统发育树分析,探究三大解毒酶家族基因的分类和进化特点,挖掘该虫的解毒酶基因序列,为进一步研究葡萄花翅小卷蛾对杀虫剂和寄主植物的适应性机制奠定基础。
1 材料与方法
1.1 RNA样本
葡萄花翅小卷蛾样本由阿根廷门多萨省农业卫生与质量研究所Gustavo Taret先生提供,成虫样本保存于RNA保存液中;RNA的提取参照Direct-zol RNA Miniprep试剂盒(Zymo research, Irvine, CA)说明书,DNase I处理去除DNA残留。利用Nano Photometer测定仪和凝胶电泳检测RNA浓度和质量。
1.2 转录组测序、组装及功能注释
将满足测序要求的高质量葡萄花翅小卷蛾总RNA送至深圳华大基因科技有限公司,通过BGISEQ-500测序平台进行转录组测序获得原始数据;使用trimmomatic进行过滤(去除包含接头、未知碱基含量>5%和低质量的reads);利用Trinity对clean reads进行de novo组装,然后使用Tgicl对组装的转录本进行聚类去冗余,获得unigene;通过Blast将组装得到的unigene进行七大功能数据库Nr(non-redundant proteins reference sequences)、Nt (nucleotide sequence database)、KEGG (kyoto encyclopedia of genes and genomes) 、Swiss-Prot、GO (gene ontology )、Pfam和KOG注释。
1.3 三大解毒酶基因家族的系统发育分析
以葡萄花翅小卷蛾3种解毒酶(P450、CarE和GST)基因的蛋白序列为基础,在NCBI中下载鳞翅目物种昆虫3种解毒酶基因的蛋白序列,使用MEGA 7程序中的MUSCLE对不同物种的P450、GST和CarE氨基酸序列进行比对(对于CarE和P450基因,选取>1000 bp的基因序列用于建树分析)。3种解毒酶基因均采用MEGA 7软件中的neighbor join (NJ)方法构建系统树,Bootstrap值设为1000。
2 结果与分析
2.1 转录组组装
为获得葡萄花翅小卷蛾的转录组数据,利用BGISEQ-500技术平台对其进行RNA测序。测序样品数据为6.4 Gb,经Trinity和Tgicl软件拼接共获得44360个unigene,其总长度、平均长度、N50及GC含量分别为49864535、1124、2078 bp和42.81%,通过BUSCO软件评估组装结果为95%,表明转录组的组装具有较高的完整性。
2.2 基因功能注释
将组装获得的44360条unigene与7个数据库进行比对。结果显示,共有28065条unigene被注释,其中通过Nr数据库成功注释的unigene数量最多,共有26388条,在总unigene数中占比59.49%(表1)。Unigene注释到Nr数据库的物种主要为棉铃虫和柑橘卷叶蛾Amyeloistransitella(Walker),分别占24.11%和16.94%(图1)。此外,根据Nr注释结果,本研究共注释了144条解毒酶基因,其中包括91条P450酶系基因、22条GST酶系基因和31条CarE酶系基因。

表1 葡萄花翅小卷蛾转录组unigene BLAST注释结果

图1 葡萄花翅小卷蛾转录组unigene在Nr数据库中的分布
2.3 P450注释分析及其与几种鳞翅目昆虫P450的进化关系
葡萄花翅小卷蛾转录组中有91个CYP基因,在解毒酶基因中所占比例最大,选取核苷酸序列长度大于1000 bp的33个P450基因进行发育进化分析。结果显示,33个P450基因可注释到CYP4、CYP6、CYP9等家族,分属于clade 2(2个基因)、clade 3(17个基因)、clade 4(8个基因)和线粒体clade(6个基因)(图2)。葡萄花翅小卷蛾P450基因均能够与鳞翅目的P450聚在一起,其中葡萄花翅小卷蛾CYP4、CYP6和CYP9家族基因与鳞翅目其他物种的同源数较多。
2.4 GST注释分析及其与几种鳞翅目昆虫GST的进化关系
注释结果显示,在葡萄花翅小卷蛾中有22个GST基因,核苷酸序列长度在590~1450 bp之间。将这些GST基因与其他鳞翅目昆虫的同源基因进行氨基酸序列比对并构建系统发育树,结果显示,葡萄花翅小卷蛾与苹果蠹蛾、云杉芽卷蛾Choristoneurafumiferana(Clem.)和棉铃虫等昆虫的相应GST蛋白可聚为一类。注释的葡萄花翅小卷蛾表达的GST分别属于Microsomal(3个)、Epsilon(7个)、Delta(5个)、Omega(3个)、Sigma(2个)、Zeta(1个)和Theta(1个)等GST亚家族,其中主要以Epsilon和Delta亚家族GST数量最多(图3)。
2.5 CarE注释分析及其与鳞翅目近缘物种的CarE进化关系
羧酸酯酶又称脂族酯酶,是酯酶超基因家族中重要的一类酶,根据其功能通常可分为消化解毒酶、激素与信息素代谢酶以及神经发育相关蛋白。在葡萄花翅小卷蛾转录组中有31个CarE基因的转录本,选取基因长度大于1000 bp的12个CarE基因构建系统发育树,发现这些CarE基因根据功能大致可以分为3类:一类承担脂质转运和代谢;另一类参与外源化合物消化和解毒,与棉铃虫CarE聚为一类;还有一类是与神经发育相关的蛋白,在葡萄花翅小卷蛾中主要为乙酰胆碱酯酶(图4)。
3 结论与讨论
葡萄花翅小卷蛾是葡萄等果树上的重要害虫,作为重要的进境检疫性有害生物,该虫在我国尚未有入侵和发生的报道。开展葡萄花翅小卷蛾代谢酶的完整分析,对于该虫入侵风险评估以及防治措施制定具有重要意义。本研究通过对葡萄花翅小卷蛾转录组测序,发现该虫表达144个解毒酶基因,包含91个细胞色素P450基因、31个CarE基因和22个GST基因。这些解毒酶基因与其他鳞翅目昆虫的同源基因存在较高的相似性。已有研究发现,不同鳞翅目昆虫基因组中的解毒酶基因存在许多同源基因对和基因群,存在较高的保守性(艾均文等,2015)。葡萄花翅小卷蛾与其他鳞翅目昆虫的解毒酶同源基因可能具有相似的代谢机制。

图2 葡萄花翅小卷蛾与其几种鳞翅目昆虫CYP的系统发育关系
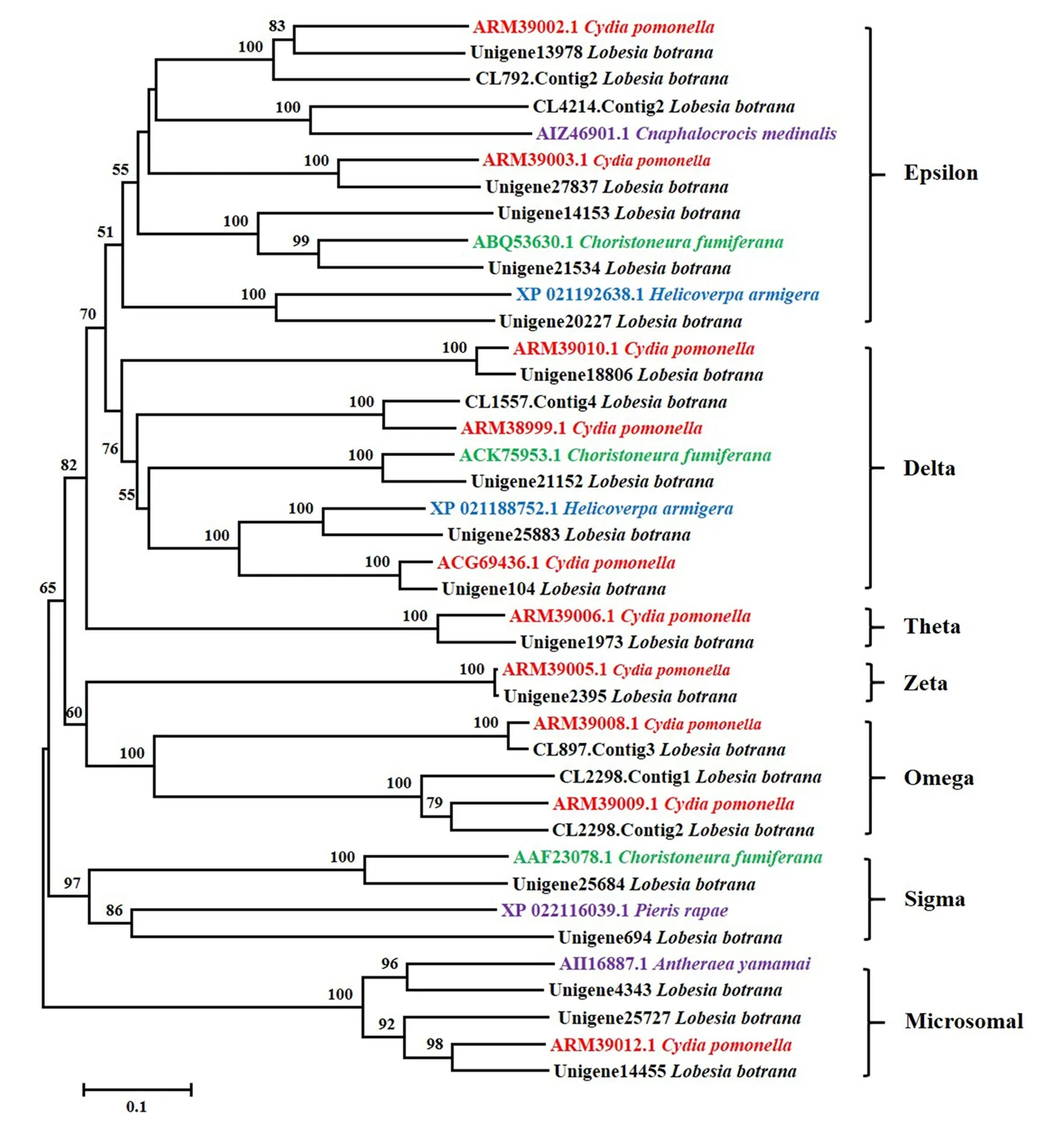
图3 葡萄花翅小卷蛾与几种鳞翅目昆虫GST的系统发育关系

图4 葡萄花翅小卷蛾与几种鳞翅目昆虫CarE的系统发育关系
细胞色素P450是昆虫体内一类重要的解毒酶,其在生物转化、内源底物的催化以及外源物质的代谢和解毒过程中具有重要作用(Guengerich,2001; Lietal.,2007; Feyereisen,1999; Rewitz,2004)。P450通过基因转录表达变化、基因突变等多种机制影响昆虫对不同寄主适应性和杀虫剂抗性(Bassetal.,2013; Wangetal.,2018)。葡萄花翅小卷蛾幼虫在不同寄主上的生长速度、交配成功率、繁殖力都存在差异,这可能与其体内P450解毒酶对植物次生代谢物质的解毒能力有关(Denis & Jerme,2005)。本研究发现,葡萄花翅小卷蛾的P450基因主要属于CYP4和CYP6家族,这2个家族的P450基因被证实与多种杀虫剂的抗性相关(郭亭亭等,2009; 杨帆和王进军,2008; Kimetal.,2017)。本研究组发现1个葡萄花翅小卷蛾的P450CYP6亚家族基因和苹果蠹蛾的CYP6B2基因在系统树上聚为一支,而CYP6B2被证实在苹果蠹蛾对溴氰菊酯和谷硫磷的抗药性中具有重要作用(Wanetal. 2019);而葡萄花翅小卷蛾的1个CYP4亚家族基因和小菜蛾CYP4g15基因聚为一支,据报道,CYP4g15参与柑橘木虱DiaphorinacitriKuwayama对农药的代谢(Tianetal.,2019);葡萄花翅小卷蛾另外1个CYP4亚家族基因和斜纹夜蛾SpodopteralitturaFabriciusCYP4C1基因聚为一支,CYP4C1被证实在多种昆虫对植物次生代谢物质中具有重要作用(Huangetal.,2019)。
CarE在昆虫对有机磷、氨基甲酸酯以及拟除虫菊酯等多种杀虫剂的抗性中起重要作用(刘红霞等,2012; 张柯等,2002; Wangetal.,2018)。CarE基因扩增和基因突变可能介导昆虫对杀虫剂产生抗性(Wangetal.,2018; Zhangetal.,2013)。羧酸酯酶CarE001A和CarE001H在棉铃虫对拟除虫菊酯类农药的代谢中具有重要的作用(Lietal.,2020)。羧酸酯酶RpCarE基因表达量的变化介导了禾谷缢管蚜RhopalosiphumpadiL.对异丙威和高效氯氟氰菊酯2种农药的抗性(Wangetal.,2018)。Navarro-Roldánetal.(2020)研究葡萄花翅小卷蛾对毒死蜱、高效氯氟氰菊酯和噻虫啉3种杀虫剂的代谢解毒机制,发现酯酶活性显著升高,羧酸酯酶活性增强可能介导该抗药性。本研究共发现31个CarE基因,这些酯酶可能参与葡萄花翅小卷蛾体内脂质转运和代谢、内源与外源化合物的消化和解毒,其功能有待进一步研究。
GST是昆虫体内三大解毒酶之一,可以通过促进杀虫剂的还原性脱氯化氢或与还原性谷胱甘肽发生偶联反应来代谢杀虫剂(Enayatietal.,2010)。GST还可以清除昆虫体内杀虫剂作用过程中产生的有毒氧自由基。大量研究表明,GST与昆虫对有机磷、有机氯、拟除虫菊酯类杀虫剂的抗性有关(尤春燕等,2013; Fournieretal.,1992),与抗性相关的昆虫GST基因分属于Delta和Epsion 2个亚族(Caoetal.,2018; Vontasetal.,2001)。此外,相关研究发现,棉铃虫GST可被植物次生代谢物质诱导,并对杀虫剂敏感性产生影响(陈凤菊等,2003; 高希武等,1997)。本研究基于葡萄花翅小卷蛾转录组数据发现22个GST基因,其中有12个GST基因属于昆虫特有的Delta和Epsion亚族,这2个亚家族的基因在昆虫对外源物质的代谢中发挥非常重要的作用(Chen & Zhang,2015),本研究鉴定的葡萄花翅小卷蛾各个GST基因的具体功能有待进一步分析。
猜你喜欢
疯狂英语·新读写(2022年1期)2022-11-23
当代水产(2022年7期)2022-09-20
河北科技师范学院学报(2022年2期)2022-08-26
轻工学报(2022年3期)2022-06-22
食品科学(2022年6期)2022-03-30
中国饲料(2021年17期)2021-11-02
空间科学学报(2021年1期)2021-05-22
世界农药(2019年3期)2019-09-10
今日农业(2019年14期)2019-01-04
分析化学(2017年12期)2017-12-25
