
美丽异木棉复合体的花色变化规律研究
2021-05-17 05:56吴欣黄颂谊黄桂莲
广东园林 2021年2期
吴欣 黄颂谊 黄桂莲
吉贝属Ceiba隶属于锦葵科(APGIV 系统),原分布于中南美洲和非洲西部,包括17 个物种[1]。该属大部分物种因观赏价值较高而被广泛引种栽培于世界各地[2~3]。据记载,我国于20 世纪60 年代开始引种“美丽异木棉”于海南尖峰岭,现今在两广地区、福建、海南和香港等地广泛栽培[2~3]。国内引进的“美丽异木棉”作为园林常用树种,不仅花期长、花色美、观赏价值高,也具有易繁殖、生长快和抗性强的特点[4~6]。根据近期的文献报道,基于形态和分子数据分析表明,目前我国华南地区栽培的“美丽异木棉”包括了吉贝属内至少3 个物种以及它们的杂种后代,其中涉及的物种有美丽异木棉C. speciosa、C. insignis和C.crispiflora[7~8]。由于国内园林常用的“美丽异木棉”并非为一个物种,因此本研究将吉贝属多个物种及其杂种称为“美丽异木棉复合体”。
美丽异木棉复合体的花瓣颜色组成较为复杂,花瓣先端颜色分为白色(浅灰色)、浅红色、红色、深红色和紫红色,花瓣基部颜色分为白色、黄白色、浅黄色和黄色[9]。目前,美丽异木棉复合体在园林应用上个体颜色参差不齐,难以形成具有群体效应的景观,对花色变化研究有利于选育出观赏价值高、花色统一的美丽异木棉复合体优良种质,形成具有群体效应的景观。有文献将美丽异木棉复合体的花瓣先端颜色和基部颜色进行分类统计[7~9],但并没有进行颜色变化相关的研究。而国内有关美丽异木棉复合体的文献报道,主要涉及形态特征[9]、光合特性[10~11]、栽培繁殖[12~18]、病理学[19]、种质分析[7~8,20]等方面。本研究对美丽异木棉复合体的花色变化规律进行观察分析,以期为美丽异木棉复合体的品种选育提供参考。
1 材料与方法
1.1 试验地点及对象
试验地点在广州市白云区的陈田花园,观测对象为来自8 株不同花色的美丽异木棉复合体个体的108 朵花,编号1~108,花色类型涵盖了粉白、粉红和紫红色3 种。为减少环境因素如温差、降雨、气流等变化的影响,观测对象均为在同一天(2019 年12月11 日)开放的花朵。观测时段为2019 年12 月11—20 日(截止时间为最后一个观测对象脱落),具体的观测时间为上午8:00—12:00。
1.2 试验方法
观察观测对象从花苞开裂至花苞脱落的过程,记录其开花状态、花瓣先端颜色、基部颜色、瓣爪颜色等,具体方法如下:
1)开花状态:根据花瓣的质感和花朵的状态,将开花的阶段划分为初绽、盛放、软化、下垂和脱落。记录数据并拍照。
2)花瓣先端颜色:使用英国皇家园林比色卡(Royal Horticultural Society, Sixth Edition 2015)记录花瓣先端颜色,比色的位置为花瓣先端的顶部1/3 位置处。记录数据并拍照。
3)花瓣基部:肉眼观察花瓣基部(花瓣底部1/3 位置处)是否出现褐化,拍照并记录数据。
4)瓣爪颜色:将花瓣中部位置的黄/白色区域定义为瓣爪区域,使用英国皇家园林比色卡记录瓣爪颜色。黄色与白色的划分以比色卡的颜色描述为准。最后记录数据并拍照。
5)数据分析:本研究将对盛放状态下的花朵的花瓣先端颜色进行聚类分析,因此需将比色卡RHS 颜色转换成RGB 数值,在SPSS v.19 中进行系统聚类分析。RHS与RGB 的颜色转换标准参照美国杜鹃花协会官方网站(https://www.azaleas.org/)提供的数据。其他数据使用excel 进行分析或图表绘制。
2 结果与分析
2.1 花期分布情况
根据统计,超过80%的观测对象花期为7~8 d,约95%的观测对象花期为6~9 d(表1)。
2.2 开花阶段变化规律
根据统计,95.4%的观测对象均经历了初绽、盛放、软化、下垂、和脱落5 个阶段(图1)。所有花朵均在初绽当天即进入盛放阶段,均在花期最后一天发生脱落。不同的开花阶段情况如表2 所示。
2.3 花瓣颜色变化规律
108 个观测对象的花瓣颜色均随着开花时间的推移而有所变化。花瓣主要分为先端、瓣爪和基部3个颜色区域,三者的颜色均发生变化(图2~3)。
2.3.1 花瓣先端颜色变化规律
以花朵盛放时的花瓣先端颜色为准,通过对观测对象的花瓣先端颜色的聚类分析,可将主要花色划分为粉白、粉红和紫红三大类别(图4)。根据观察,随着开花时间推移,观测对象均显示出花瓣先端颜色发生变化的情况,变化趋势均为逐渐弱化、淡化,如粉白色花的变化趋势为粉白色逐渐淡化为白色;粉红色花的颜色慢慢转变为更浅的粉色;深紫红色花则变化较广,最终颜色为淡紫红色或粉红色。
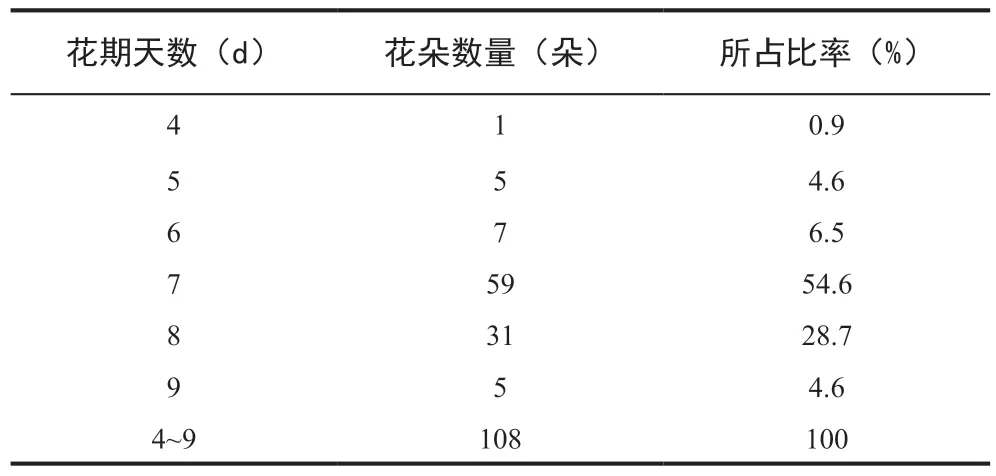
表1 观测对象的花期情况

图1 同一观测对象在完整花期内不同开花阶段的表现
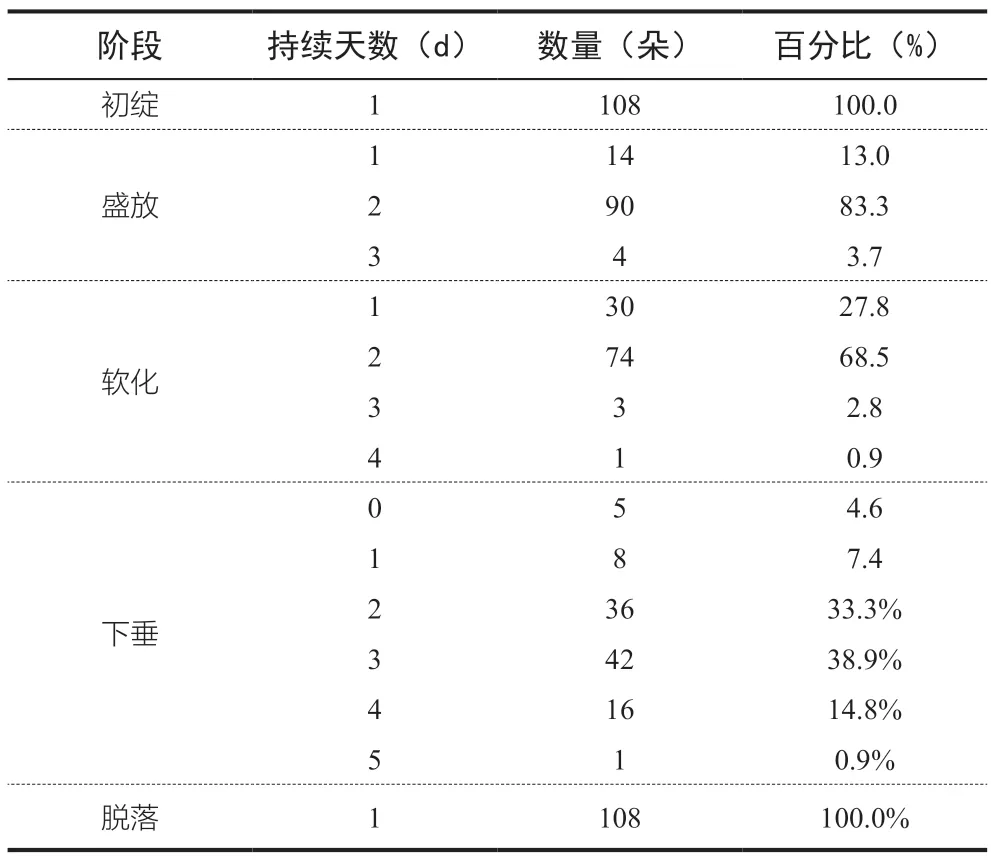
表2 观测对象开花的各阶段情况

图2 开花前期的花瓣
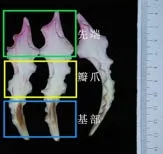
图3 开花后期的花瓣
2.3.2 瓣爪颜色变化规律
据观测,瓣爪颜色主要有黄色和白色2 种,所有观测对象的瓣爪初始颜色均为黄色,随着时间推移而逐渐淡化为白色。根据记录,观测对象的黄瓣爪时期均为开花后的1~3 d,较为集中;而白瓣爪时期的持续时间差异较大,为0~7 d(表3)。据统计,大部分观测对象的黄瓣爪时期占其花期的20%~40%,平均黄瓣爪时期占比为30%;白瓣爪时期的占比则为50%~70%,平均白瓣爪时期占比为56%(图5)。
2.3.3 花瓣基部颜色变化规律
在瓣爪颜色发生变化的同时,花瓣基部的颜色也发生改变。据观察,观测对象的花瓣基部均出现褐化现象,但花瓣基部开始出现褐化的时间不一。据统计,花瓣基部褐化通常在开花后的2~4 d 开始出现,出现的时间主要为第3 d(图6)。结合瓣爪由黄色转变为白色的时间节点,发现71%的观测对象瓣爪颜色变为白色和花瓣基部开始褐化的时间节点重叠。当花瓣基部开始出现褐化之后,褐化(颜色)程度逐渐加深,同时褐化范围逐渐由花瓣基部向瓣爪部位延伸,直至花瓣脱落。
2.3.4 不同开花阶段的花瓣颜色变化
研究显示,美丽异木棉复合体花瓣瓣爪和花瓣基部在开花阶段中出现明显的颜色变化,并且颜色变化与开花阶段有较明显的相关性,而花瓣顶端颜色变化相对不明显。所有观测对象的花瓣瓣爪在开花阶段早期(初绽和盛放)呈现黄色,开花阶段中期(软化)颜色逐渐淡化,开花后期(下垂)完全变成白色。因此,观测对象处于初绽和盛放阶段时,花瓣瓣爪的颜色为黄色,软化阶段可能为黄色或白色,下垂阶段则为白色。此外,所有观测对象的花瓣基部在开花阶段早期(初绽和盛放)没有褐化,开花阶段中期(软化)逐渐褐化,开花后期(下垂)完全褐化。因此,观测对象处于初绽和盛放阶段时,花瓣基部没有褐化,软化阶段为逐渐褐化,下垂阶段则为完全褐化。

图4 美丽异木棉复合体(盛放时)花瓣先端颜色的聚类分析

表3 观测对象的瓣爪颜色变化情况
3 结论与讨论
3.1 花期与开花阶段
本研究涉及同一天开花的108 朵美丽异木棉复合体花朵在其完整花期内的动态变化,包括花期、开花状态、花瓣先端颜色、瓣爪颜色、基部颜色等变化过程。研究结果表明,大部分美丽异木棉复合体花朵开花时间为6~9 d。在整个花期内,开花状态是动态变化的,主要经历了初绽、盛放、软化、下垂和脱落5 个阶段。少部分花朵缺失了部分状态,如下垂阶段,且其花期往往相对较短,因此推断这些花朵很可能是在其他时间段(如当天下午或夜间)处于这些阶段,但没有被观测到,在未来的进一步研究中需要增加下午和晚上的观测时间。
3.2 花瓣颜色变化规律
本研究显示,美丽异木棉复合体花瓣颜色存在差异,主要有2 个因素:1)花瓣固有的颜色差异,如盛放时期花瓣先端有粉白色、粉红色、紫红色三大类。2)花瓣自身演变,如花瓣先端颜色可由粉白色淡化为白色(浅灰色),深紫红色逐渐转为紫红色,深红色慢慢转化为浅红色等;花瓣基部颜色在花朵初绽时为黄色,随着花期逐渐淡化为浅黄色,最终变为白色;花瓣基部逐渐褐化,并且褐色不断加深,直至花朵脱落。
本次研究发现部分观测对象(例如2、3、10 号)不仅花期长(9 d),而且瓣爪颜色变化时间长(黄瓣爪持续4 d),褐化时间晚(第4 天开始出现褐化)。此类型的个体更具有园林观赏价值,可作为进一步的园林应用或育种材料。
3.3 花瓣颜色与开花阶段的联系
开花阶段可由花瓣的瓣爪颜色和花瓣基部是否褐化进行预判:1)如瓣爪为黄色和花瓣基部没有褐化,则观测对象处于开花阶段的初期—初绽和盛放阶段;2)瓣爪为白色和花瓣基部已经褐化,表明观测对象处于开花阶段的后期—下垂阶段;3)观测对象处于软化阶段时,花瓣基部和瓣爪颜色处于上述描述的过渡状态。
本研究发现,美丽异木棉复合体花朵颜色组成的复杂性不仅来源于花瓣先端固有的颜色差异,还有来源于花朵在花期内的动态变化。为挖掘美丽异木棉复合体的花朵状态及颜色变化的本质及影响因素,后续可对该类群进行分子层面的分析如qPCR、转录组测序、基因组重测序等,以探讨其变化的内在分子机制。
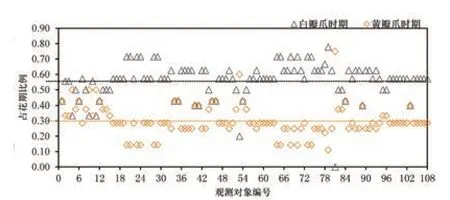
图5 黄/白瓣爪时期占花期的比例情况

图6 观测对象花瓣基部褐化与瓣爪颜色变化时间的比较情况
猜你喜欢
农业工程学报(2022年13期)2022-10-09
大气科学学报(2022年2期)2022-05-12
聊城大学学报(自然科学版)(2022年3期)2022-02-14
作物学报(2022年3期)2022-01-22
参花(下)(2021年3期)2021-03-24
歌海(2021年6期)2021-02-01
海峡姐妹(2019年5期)2019-06-18
热带农业科学(2017年9期)2017-10-23
海峡姐妹(2017年3期)2017-04-16
农村农业农民·B版(2016年7期)2016-10-21
