
黄海北部滤食性贝类供饵力区划研究
2021-05-24 01:52刘苏萱
水产科学 2021年3期
张 雪,宋 伦,付 杰,王 昆,刘 印,刘苏萱
( 1.辽宁省自然资源事务服务中心,辽宁 沈阳 110001; 2.辽宁省海洋水产科学研究院,辽宁 大连 116023; 3.大连海洋大学,辽宁 大连 116023 )
黄海北部海域是菲律宾蛤仔(Ruditapesphilippinarum)、虾夷扇贝(Patinopectenyessoensis)、长牡蛎(Crassostreagigas)等滤食性贝类主要增养殖区,近些年增养殖贝类普遍出现滞长、肥满度下降、死亡率升高等现象,严重制约海洋渔业的可持续发展[1-2]。相关研究发现,滤食性贝类对饵料微藻具有选择性策略,大粒级微藻更利于滤食性贝类的营养储备和健康生长[3-4]。然而受技术局限,微藻粒级结构研究进展缓慢,目前常用的叶绿素a分级法由于大孔径滤膜较高的截留率严重低估了小粒径微藻的生物量[5-7]。以往研究贝类饵料生物量大多利用叶绿素a含量表征饵料微藻生物量[8-10],可能会高估滤食性贝类的有效饵料量,进而可能会高估贝类养殖容量。因此,研究滤食性贝类饵料微藻供应能力(即供饵力)对揭示贝类营养能量及免疫水平至关重要。对于滤食性贝类,有效饵料微藻(大粒级藻类)的比例和总量及存在周期可反映增养殖区滤食性贝类的供饵力水平,笔者采用高通量测序—分子鉴定分级技术对黄海北部海域进行饵料微藻供饵力区划,并测算贝类养殖容量,以期为黄海北部海域滤食性贝类增养殖提供数据参考。
1 材料与方法
1.1 试验海域与采样站位
2019年,在黄海北部海域设置53个站位(图1),分别于春季(5月)、夏季(8月)、秋季(10月)采

图1 黄海北部海域采样站位示意Fig.1 Sampling site innorthern Yellow Sea
集真核微藻分子鉴定样品和叶绿素a样品。每个站位采集表层海水1 L,现场用0.22 μm微孔滤膜收集全部微藻,然后将滤膜转移至1.5 mL无菌离心管中,置于-20 ℃或-80 ℃冷冻保存、运输,用于真核微藻分子鉴定。
1.2 分析方法
1.2.1 饵料微藻分子鉴定
饵料微藻DNA提取采用CTAB法(十六烷基三甲基溴化铵),PCR(聚合酶链式反应)扩增引物利用自行开发的真核微藻18S rDNA V4区基因扩增引物,具体参考文献[2]。
1.2.2 叶绿素a含量检测
叶绿素a样品调查、检测方法参照GB/T 12763—2007《海洋调查规范》[11]和GB 17378—2007《海洋监测规范》[12]执行。
1.2.3 数据分析
分子测序利用解析软件(http:∥drive5.com/uparse/)对有效数据进行运算分类单元(OTUs)聚类和物种分类,采用朴素贝叶斯分类方法与数据库(https:∥www. arb-silva.de/,Version 108)对运算分类单元代表序列进行物种注释[13]。根据相关研究,不同粒径的真核微藻序列数比例更接近于生物量比例[14-16],将各个站位所获得的优势度超过0.1%的种类均作为整体优势种参与粒级生物量统计,参考文献[14-20]将筛查出的优势种粒径进行筛检分级,即将优势种按粒径大小划分为大粒级(>10 μm)和小粒级(<10 μm)两组,球状藻类以直径统计粒径大小,杆状藻类以长宽换算为球径统计粒径大小,根据各粒径微藻序列占比统计其粒级结构,同时利用序列数占比和总叶绿素a含量测算某种粒级微藻的叶绿素a含量,相关公式[5]:
式中,ρBi为某粒级微藻的叶绿素a质量浓度(μg/L);ρi为总叶绿素a质量浓度(μg/L);DC为某粒级微藻的序列数;NC为所有微藻的序列数。
物种生物量优势度(Y)表示微藻群落中某一物种质量所占的优势程度[5]:
式中,nx为第x种微藻种类的运算分类单元数,N为运算分类单元总数,fx为第x种微藻种类在各样品中出现的频率。
由于大部分滤食性贝类对大粒级(>10 μm)饵料微藻截留效率较高[4],根据供饵力含义,即有效饵料微藻(大粒级藻类)供应能力,本试验供饵力指数(Ai)专指大粒级微藻生物量与其占比乘积:
Ai=Bi×Ci%
式中,Bi为第i站位大粒级微藻生物量,Ci%为第i站位大粒级微藻占比。
贝类养殖容量参照Parsons-Takahashii营养动态模型,根据海域初级生产者微藻的生产量和生态效率,估算各滤食性贝类的现存生物量[21-22]。
上述公式计算、数据分析、分布饼图绘制均通过WPS Office、SPSS 19.0、ARGIS 10.2等软件完成。
2 结 果
2.1 大粒级微藻优势种
黄海北部海域主要以小粒级(<10 μm)微藻为主,春季、夏季、秋季大粒级(>10 μm)微藻生物量占比分别为(39±10)%、(35±14)%、(38±11)%(图2)。大粒级微藻优势种在各季节演替明显,分别由不同种类占据主要生态位。但大部分微藻都是有毒甲藻,对海域贝类增养殖贝毒累积风险较高。细小微胞藻(Micromonaspusilla)和金牛微球藻(Ostreococcustauri)在小粒级微藻群落中占有绝对优势,春季、夏季、秋季演替规律由细小微胞藻占优势转为金牛微球藻控制生态位(表1)。还有部分未获注释的类群优势度也较高,如共甲藻目。

表1 春、夏、秋季各粒级生物量优势种Tab.1 Dominant species of biomass at various grain grades in spring, summer and autumn

图2 大粒级微藻生物量占比Fig.2 Proportion of biomass of large microalgae
2.2 贝类供饵力区划
黄海北部海域各站位微藻平均供饵力指数分布见图3,将53个站位按供饵力指数高低大致平均分为3组,其中高供饵力组平均指数>0.51,中供饵力组平均指数为0.23~0.51,低供饵力组平均指数<0.23。春季供饵力最高,平均为0.86;秋季次之,平均为0.41;夏季最少,平均为0.30。其中,20号站位最高,14号站位次之。
黄海北部海域春、夏、秋季及平均供饵力区划见图4,春季高供饵力主要集中在庄河近岸、鸭绿江口海域,夏季高供饵力主要集中在旅顺、庄河、东港近岸海域,秋季高供饵力主要集中在旅顺、庄河近岸海域,总体而言旅顺、庄河、鸭绿江口海域滤食性贝类供饵力相对较高,长海县及外海相对较低。
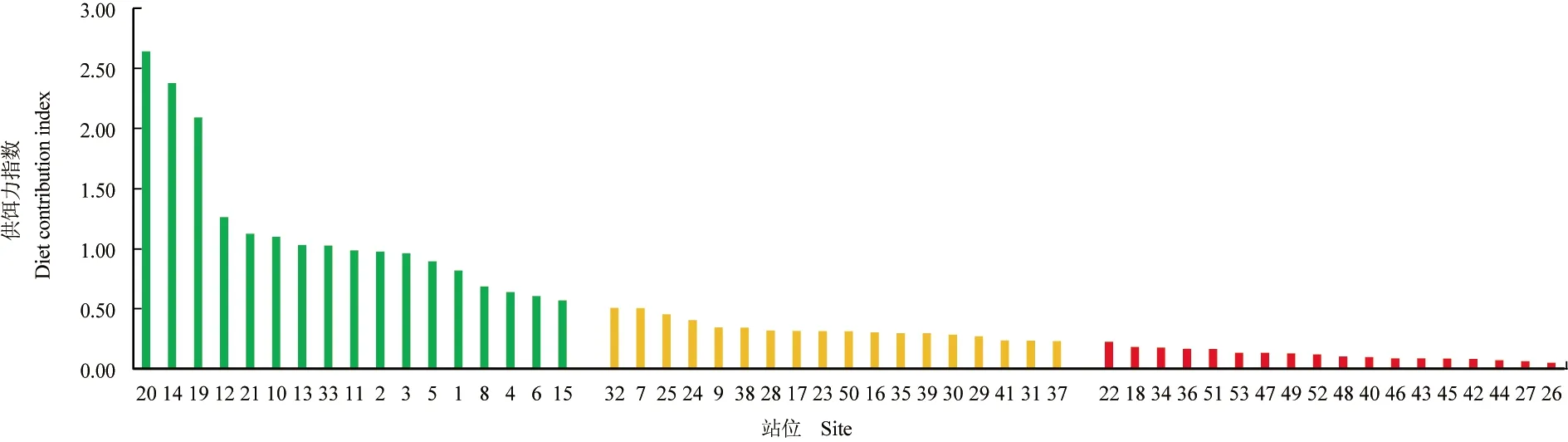
图3 各站位平均供饵力指数Fig.3 Average diet contribution index at each station

图4 黄海北部海域供饵力区划Fig.4 Diet contribution zoning map in northern Yellow Sea
2.3 贝类养殖容量估算
由于大部分滤食性贝类对10 μm以上饵料微藻截留效率较高[4,23],参照Parsons-Takahashii营养动态模型,分别利用总叶绿素a和10 μm以上粒级微藻生物量估算总贝类和菲律宾蛤仔养殖容量。对比结果(图5、图6)显示,目前利用总叶绿素a作为滤食性贝类饵料基础会高估养殖容量,总贝类养殖容量高估约4倍,菲律宾蛤仔养殖容量平均高估2.6倍,黄海北部全海域实际可承载贝类容量春季为151 g/m2、夏季为51 g/m2、秋季为96 g/m2,可承载菲律宾蛤仔养殖容量春季为100 g/m2、夏季为34 g/m2、秋季为63 g/m2,黄海北部海域贝类养殖容量主要为春、秋季贡献。

图5 黄海北部海域贝类养殖容量估算Fig.5 Estimation of shellfish culture capacity in northern Yellow Sea 测算.利用总叶绿素a估算养殖容量;修正.利用大粒级微藻生物量估算养殖容量;下同. Measure.total chl-a is used to estimate culture capacity; correction. the biomass of microalgae of large grain size is used to estimate the culture capacity; et sequentia.
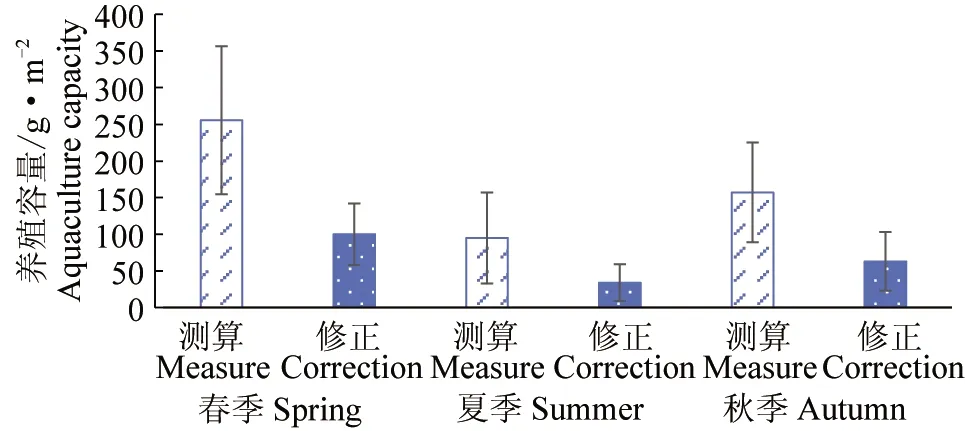
图6 黄海北部海域菲律宾蛤仔养殖容量估算Fig.6 Estimation of culture capacity of Manila clam R. philippinarum in northern Yellow Sea
各站位基于大粒级微藻生物量估算的总贝类和菲律宾蛤仔养殖容量见图7。黄海北部贝类养殖容量分布基本与供饵力站位分布一致,个别站位由于透明度较低,饵料总供应量较少,导致贝类养殖容量估算较低。
3 讨 论
3.1 黄海北部海域微藻粒级特点
滤食性贝类摄食机制有两种:一是靠黏液纤毛作用主动筛选适口饵料,二是靠水动力作用被动吸收适口饵料。研究发现,贻贝等主要依靠黏液纤毛作用,扇贝主要依靠水动力作用,牡蛎两种摄食机制同时利用;滤食性贝类的摄食效率随着饵料粒径的减小而降低,大部分贝类对10 μm以上饵料微藻截留效率较高,对3 μm以下饵料微藻截留效率最低[4,23]。
黄海北部海域主要增养殖对象为菲律宾蛤仔、虾夷扇贝、长牡蛎等滤食性贝类,对天然饵料微藻需求量较大。2019年黄海北部海域菲律宾蛤仔产量7.3×105t、扇贝3.3×105t、牡蛎2.7×105t(数据来源于辽宁省2019年渔业统计年报)。虽然鸭绿江、大洋河等每年携带陆源氮磷等营养盐入海,但随着入海河流污染整治,氮、磷消减力度加大,海域无机营养盐已呈下降趋势,对微藻生物量及粒级结构影响较大[7,24-25]。调查发现,黄海北部海域微藻群落主要以小粒级微藻(<10 μm)为主,对贝类增养殖业潜在影响较大[4]。2018年大长山岛临近海域小粒径(<3 μm)真核微藻生物量组成春季为86%、夏季为52%、秋季为20%,春季优势种抑食金球藻生物量占41%,夏季优势种金牛微球藻生物量占21%,这两种微微型藻类会严重影响滤食性贝类正常摄食生长,养殖风险较高[25-26]。因此应高度关注小粒级微藻对贝类增养殖业的不利影响,加强养殖海域供饵力研究。
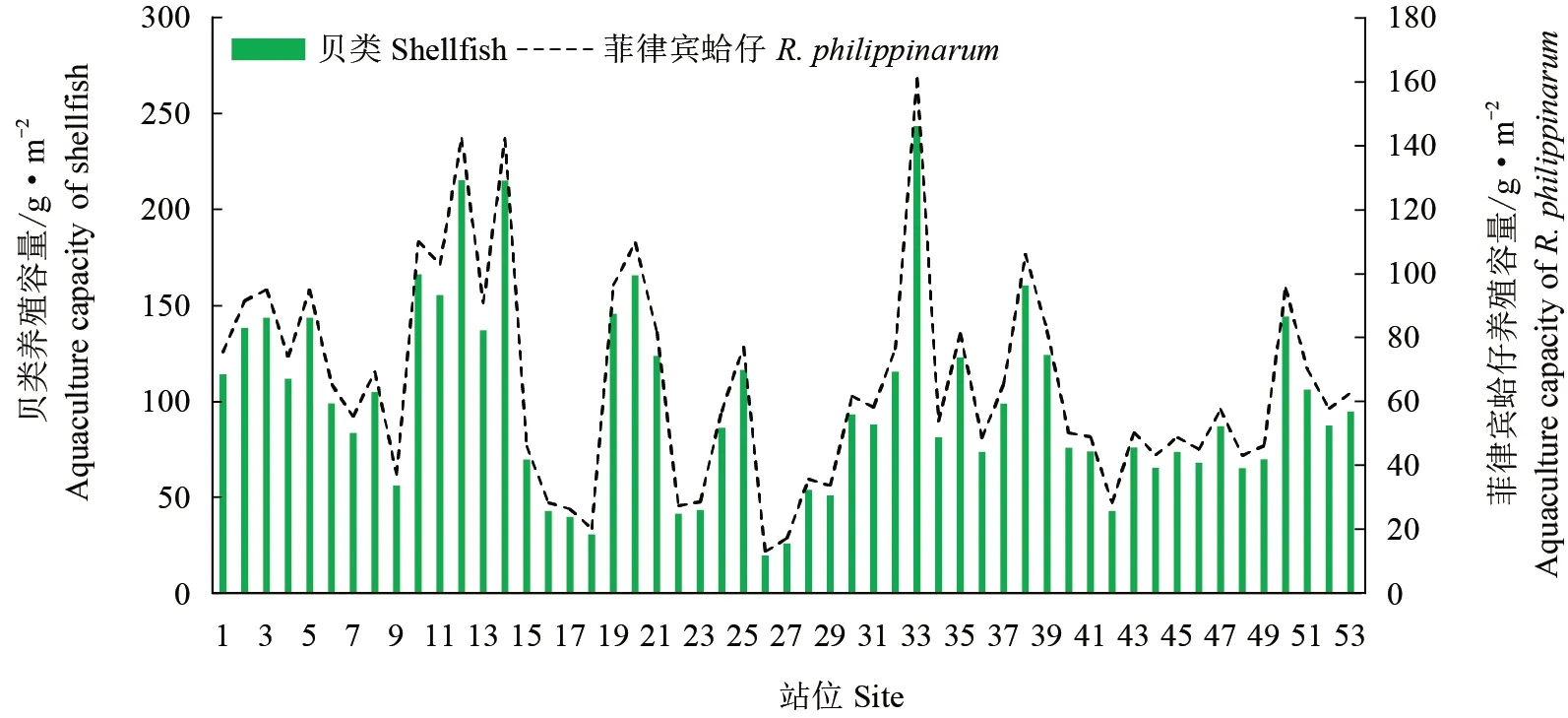
图7 基于大粒级微藻生物量估算的贝类养殖容量Fig.7 Estimation of shellfish culture capacity based on microalgae biomass of large grain size
3.2 黄海北部贝类饵料微藻供饵力区划
有关贝类饵料微藻供饵力研究进展缓慢,主要受限于微藻粒级测定技术。随着分子生物学的快速发展,突破了微微型藻类的鉴定壁垒,然而不同真核生物中rDNA的拷贝数差异较大,因此扩增序列数并不能表征微藻的丰度,但微藻rDNA的拷贝数可表征其生物量,解决了微藻粒级结构研究的理论基础[14-16]。本试验结果显示,黄海北部贝类饵料供饵力和养殖容量主要由春、秋季贡献,旅顺、庄河、鸭绿江口海域相对较高,长海县及外海相对较低。以往研究贝类养殖容量常用总叶绿素a含量表征海区的供饵能力,但滤食性贝类对小粒级微藻利用率极低,笔者发现高估养殖容量约4倍。当然,滤食性贝类也并不单纯摄食微藻,也会被动滤食有机碎屑、鞭毛虫、纤毛虫等,随着研究的深入,供饵力指数也需要进一步优化。饵料微藻的生物量和种类是决定滤食性贝类养殖容量的主要因素,笔者测算的养殖容量偏低,主要由于测算初级生产力时剔除了小粒级微藻所致,更能反映该海域养殖容量实际情况。
4 结 论
(1)黄海北部海域春、夏、秋季主要以小粒级(<10 μm)微藻为主,细小微胞藻和金牛微球藻在小粒级微藻群落中占有绝对优势,春、夏、秋季演替规律由细小微胞藻占优势转为金牛微球藻控制生态位。
(2)黄海北部海域春季高供饵力主要集中在庄河近岸、鸭绿江口海域,夏季高供饵力主要集中在旅顺、庄河、东港近岸海域,秋季高供饵力主要集中在旅顺、庄河近岸海域,总体而言,旅顺、庄河、鸭绿江口海域滤食性贝类供饵力相对较高,长海县及外海相对较低。
(3)黄海北部全海域实际可承载贝类容量春季为151 g/m2、夏季为51 g/m2、秋季为96 g/m2,可承载菲律宾蛤仔养殖容量春季为100 g/m2、夏季为34 g/m2、秋季为63 g/m2,黄海北部贝类养殖容量主要为春季、秋季贡献。
猜你喜欢
水产科学(2022年5期)2022-09-21
矿产保护与利用(2022年3期)2022-08-12
红领巾·探索(2021年8期)2021-08-26
知识就是力量(2021年6期)2021-07-09
矿冶(2021年1期)2021-04-22
皮革制作与环保科技(2021年24期)2021-03-07
贵州大学学报(自然科学版)(2020年1期)2020-02-06
食品与生活(2019年8期)2019-10-30
食品与生活(2017年11期)2017-12-18
科学与财富(2017年6期)2017-03-19
