
喷施外源茉莉酸类化合物对红松防御酶及蛋白酶抑制剂的影响1)
2021-05-27 08:44王琪杨浩伦乌恩泰严善春
东北林业大学学报 2021年5期
王琪 杨浩伦 乌恩泰 严善春
(黑龙江省森林保护研究所,哈尔滨,150040)(林木遗传育种与生物技术教育部重点实验室(东北林业大学))
冷杉梢斑螟(Dioryctriaabietella)和赤松梢斑螟(D.sylvestrella)是红松(Pinuskoraiensis)球果和枝梢的重要害虫,大发生年份球果被害率达80%以上,受其危害后红松种子的质量和产量大幅下跌;树干分叉,木材材质和出材率明显下降,严重影响林区的经济效益和生态建设[1]。2种梢斑螟成虫羽化期较长,由每年6月持续至8月[2],且幼虫生活于球果或枝条虫包内,传统药剂防治效果欠佳。植物的诱导抗性可以增强植物自身的抗虫能力,从而抵御害虫对寄主植物的危害,适用于防御营隐蔽性生活的害虫[3]。
茉莉酸甲酯(MJA)和茉莉酮(ZJ)是茉莉酸防御反应途径中重要的中间产物,在植物防御、生长及代谢过程中发挥重要作用[4]。茉莉酸甲酯或茉莉酮可作为外源诱导剂[4-6],诱导植物产生抗性反应。研究认为2种物质诱导反应途径不同,诱导的防御策略也不同[5-8]。茶树(Camelliasinensis)[9]、云杉(Piceaabies)[10]、落叶松(Larixgmelinii)和杨树(Populusspp.)[11]等许多植物在经茉莉酸甲酯处理后,均可诱导其产生系统性的诱导抗性,可通过直接或间接的防御反应抵御害虫[12]。茉莉酮诱导的防御研究相对较少,早期的研究认为与茉莉酸途径的其他茉莉酸异构体相比,茉莉酮更主要是通过刺激植物特异性挥发物引诱天敌来控制害虫,既采用间接防御策略减少害虫危害[13]。随着研究深入人们发现,用茉莉酮处理也能够诱导小麦(Triticumaestirum)自身多种防御物质质量分数增加[14]。课题组先前的研究结果同样显示利用外源茉莉酸甲酯、茉莉酮均可诱导红松酚酸质量分数增加,且茉莉酮诱导的酚酸种类更多[15]。但具体的机制尚不十分明确。过氧化氢酶(CAT)、超氧化物歧化酶(SOD)、过氧化物酶(POD)、多酚氧化酶(PPO)和苯丙氨酸解氨酶(PAL)是植物诱导防御反应过程的关键酶[3,12-16];蛋白酶抑制剂(PPI)可与害虫消化道蛋白酶相互作用,使昆虫产生厌食反应[12]。本研究利用茉莉酸甲酯、茉莉酮的系统及局部喷施,研究诱导处理后红松主梢和球果防御酶及蛋白酶抑制剂的活性变化,讨论应用不同种类茉莉酸类化合物诱导的红松防御反应的差异,从而为探索利用植物化学防御控制红松梢斑螟危害奠定理论基础。
1 材料与方法
1.1 供试材料
于2016年4月末,在牡丹江林口林业局刁翎红松人工林内,选择40年生红松为试验植株。
1.2 样品采集及相关酶活性测定
将健康红松样树随机分成16组,用不同浓度的茉莉酸类化合物进行原位喷雾诱导和系统喷雾诱导处理。原位诱导即喷施部位与样品采集部位相同,系统诱导即喷雾部位与样品采集部位不同,具体处理方式见表1。每处理组120棵样树,分别在5个时间段,即处理后的1、30、60、90、120 d摘取红松球果(2年生)及主梢,每次取样24株。赤松梢斑螟的第1次转移期,是初孵幼虫由主梢转移到球果,此转移期相应于本研究的1~30 d;其第2个转移期,是5龄老熟幼虫由球果转移回主梢取食越冬,此转移期相应于本研究的90~120 d。采集未喷施任何化合物的健康球果和主梢,作为健康对照;采集梢斑螟危害的红松球果和主梢,作为虫害对照。
将采集到的球果及主梢装入夹链袋中,迅速放入冰盒内带回室内,于-20 ℃保存、备测。CAT、SOD、POD、PPO和PAL,蛋白酶抑制剂的测定方法参考文献[16]。
1.3 数据分析
采用SPSS 11.5软件计算防御酶、蛋白酶抑制剂活性的平均值和标准差。采用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD,α=0.05)比较不同处理、不同时间段防御酶、蛋白酶抑制剂活性的差异显著性。
2 结果与分析
2.1 健康、虫害及不同茉莉酸类化合物处理的红松球果和主梢防御酶活性
健康、虫害及不同浓度茉莉酸类化合物原位喷雾或系统喷雾处理,诱导红松球果内各防御酶随时间的变化趋势见表2。由表2可见,10 mmol·L-1茉莉酮原位及系统喷施茉莉酮诱导的球果SOD活性,只在处理60 d时略低于虫害取食诱导,其他处理时间段高于(处理90 d)或与虫害取食无显著差异。健康球果CAT活性随其生长不断增加,但虫害诱导的CAT活性在检测的1~120 d均显著高于健康球果,以不同浓度茉莉酸类化合物诱导球果CAT,发现尤以10 mmol·L-1茉莉酮原位及系统喷施球果诱导的CAT活性与虫害诱导最为接近。不同浓度茉莉酸类化合物处理诱导的球果POD活性在处理1 d时显著增加,与茉莉酸甲酯相比,茉莉酮诱导的POD更接近虫害取食诱导。不同浓度茉莉酸类化合物处理诱导的球果PPO活性在处理1 d后显著增加,10 mmol·L-1茉莉酸甲酯诱导的PPO活性与虫害诱导无显著差异,100 mmol·L-1茉莉酸甲酯诱导的PPO活性显著高于虫害诱导,10 mmol·L-1茉莉酮原位喷施及100 mmol·L-1茉莉酮原位、系统喷施,诱导的PPO活性均显著高于100 mmol·L-1茉莉酸甲酯诱导。方差分析结果表明,在检测的30~120 d,10 mmol·L-1茉莉酮原位及系统喷施诱导的球果PPO活性与虫害诱导无显著差异。分析不同茉莉酸类化合物诱导的球果PAL活性,发现10 mmol·L-1茉莉酮原位诱导的PAL,在处理的1~120 d与虫害诱导无显著差异。

表2 健康、虫害及不同浓度茉莉酸类化合物处理的红松球果防御酶活性
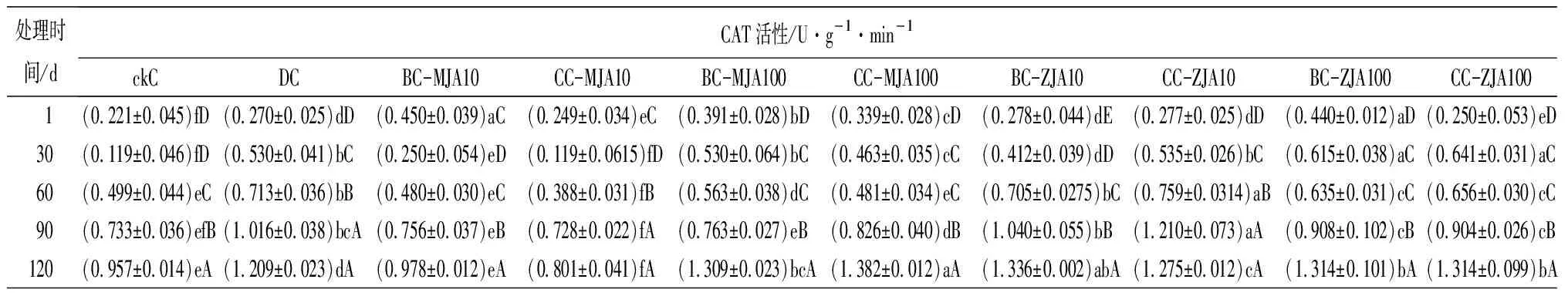
处理时间/dCAT活性/U·g-1·min-1ckCDCBC-MJA10CC-MJA10BC-MJA100CC-MJA100BC-ZJA10CC-ZJA10BC-ZJA100CC-ZJA100 1(0.221±0.045)fD(0.270±0.025)dD(0.450±0.039)aC(0.249±0.034)eC(0.391±0.028)bD(0.339±0.028)cD(0.278±0.044)dE(0.277±0.025)dD(0.440±0.012)aD(0.250±0.053)eD30(0.119±0.046)fD(0.530±0.041)bC(0.250±0.054)eD(0.119±0.0615)fD(0.530±0.064)bC(0.463±0.035)cC(0.412±0.039)dD(0.535±0.026)bC(0.615±0.038)aC(0.641±0.031)aC60(0.499±0.044)eC(0.713±0.036)bB(0.480±0.030)eC(0.388±0.031)fB(0.563±0.038)dC(0.481±0.034)eC(0.705±0.0275)bC(0.759±0.0314)aB(0.635±0.031)cC(0.656±0.030)cC90(0.733±0.036)efB(1.016±0.038)bcA(0.756±0.037)eB(0.728±0.022)fA(0.763±0.027)eB(0.826±0.040)dB(1.040±0.055)bB(1.210±0.073)aA(0.908±0.102)cB(0.904±0.026)cB120(0.957±0.014)eA(1.209±0.023)dA(0.978±0.012)eA(0.801±0.041)fA(1.309±0.023)bcA(1.382±0.012)aA(1.336±0.002)abA(1.275±0.012)cA(1.314±0.101)bA(1.314±0.099)bA
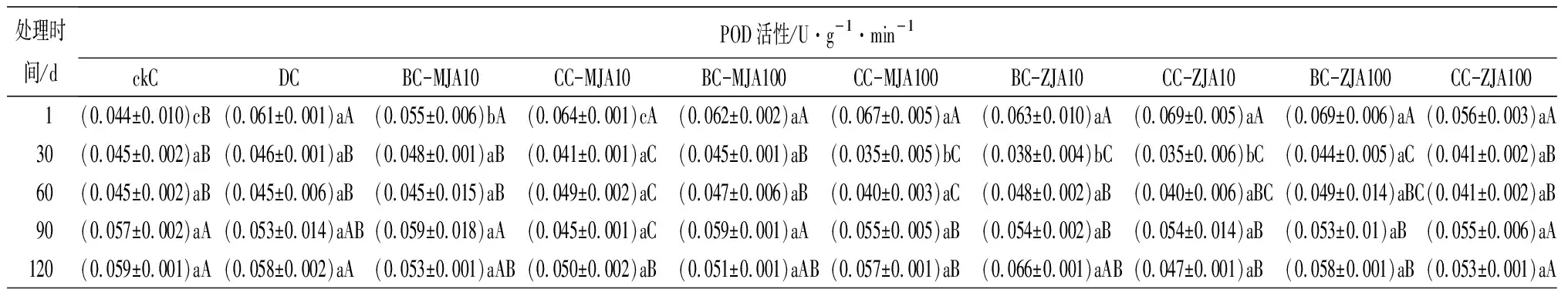
处理时间/dPOD活性/U·g-1·min-1ckCDCBC-MJA10CC-MJA10BC-MJA100CC-MJA100BC-ZJA10CC-ZJA10BC-ZJA100CC-ZJA100 1(0.044±0.010)cB(0.061±0.001)aA(0.055±0.006)bA(0.064±0.001)cA(0.062±0.002)aA(0.067±0.005)aA(0.063±0.010)aA(0.069±0.005)aA(0.069±0.006)aA(0.056±0.003)aA30(0.045±0.002)aB(0.046±0.001)aB(0.048±0.001)aB(0.041±0.001)aC(0.045±0.001)aB(0.035±0.005)bC(0.038±0.004)bC(0.035±0.006)bC(0.044±0.005)aC(0.041±0.002)aB60(0.045±0.002)aB(0.045±0.006)aB(0.045±0.015)aB(0.049±0.002)aC(0.047±0.006)aB(0.040±0.003)aC(0.048±0.002)aB(0.040±0.006)aBC(0.049±0.014)aBC(0.041±0.002)aB90(0.057±0.002)aA(0.053±0.014)aAB(0.059±0.018)aA(0.045±0.001)aC(0.059±0.001)aA(0.055±0.005)aB(0.054±0.002)aB(0.054±0.014)aB(0.053±0.01)aB(0.055±0.006)aA120(0.059±0.001)aA(0.058±0.002)aA(0.053±0.001)aAB(0.050±0.002)aB(0.051±0.001)aAB(0.057±0.001)aB(0.066±0.001)aAB(0.047±0.001)aB(0.058±0.001)aB(0.053±0.001)aA
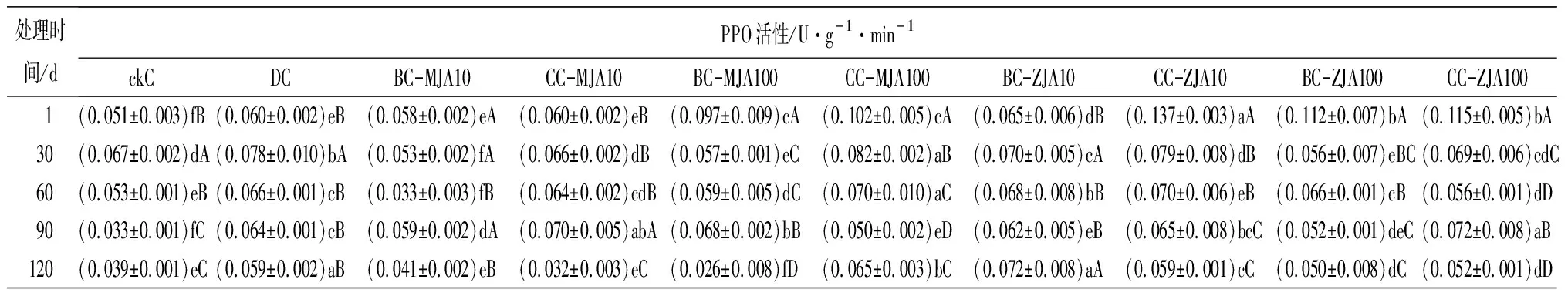
处理时间/dPPO活性/U·g-1·min-1ckCDCBC-MJA10CC-MJA10BC-MJA100CC-MJA100BC-ZJA10CC-ZJA10BC-ZJA100CC-ZJA100 1(0.051±0.003)fB(0.060±0.002)eB(0.058±0.002)eA(0.060±0.002)eB(0.097±0.009)cA(0.102±0.005)cA(0.065±0.006)dB(0.137±0.003)aA(0.112±0.007)bA(0.115±0.005)bA30(0.067±0.002)dA(0.078±0.010)bA(0.053±0.002)fA(0.066±0.002)dB(0.057±0.001)eC(0.082±0.002)aB(0.070±0.005)cA(0.079±0.008)dB(0.056±0.007)eBC(0.069±0.006)cdC60(0.053±0.001)eB(0.066±0.001)cB(0.033±0.003)fB(0.064±0.002)cdB(0.059±0.005)dC(0.070±0.010)aC(0.068±0.008)bB(0.070±0.006)eB(0.066±0.001)cB(0.056±0.001)dD90(0.033±0.001)fC(0.064±0.001)cB(0.059±0.002)dA(0.070±0.005)abA(0.068±0.002)bB(0.050±0.002)eD(0.062±0.005)eB(0.065±0.008)bcC(0.052±0.001)deC(0.072±0.008)aB120(0.039±0.001)eC(0.059±0.002)aB(0.041±0.002)eB(0.032±0.003)eC(0.026±0.008)fD(0.065±0.003)bC(0.072±0.008)aA(0.059±0.001)cC(0.050±0.008)dC(0.052±0.001)dD
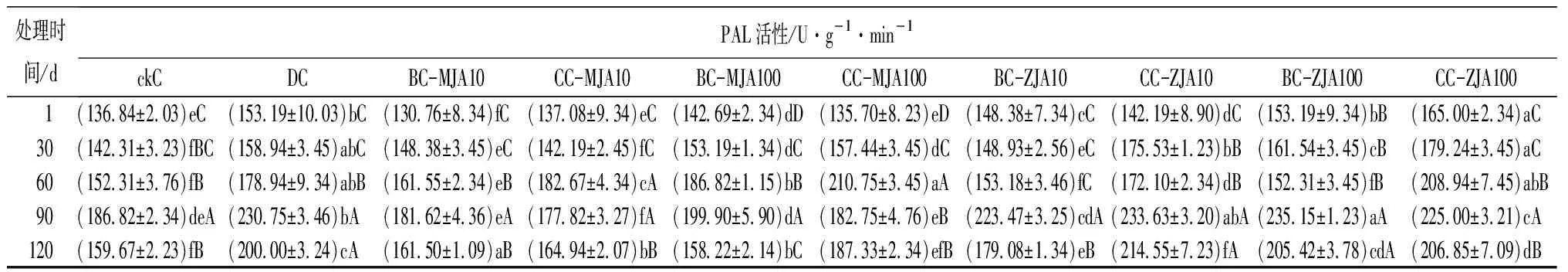
处理时间/dPAL活性/U·g-1·min-1ckCDCBC-MJA10CC-MJA10BC-MJA100CC-MJA100BC-ZJA10CC-ZJA10BC-ZJA100CC-ZJA100 1(136.84±2.03)eC (153.19±10.03)bC(130.76±8.34)fC(137.08±9.34)eC(142.69±2.34)dD(135.70±8.23)eD(148.38±7.34)cC(142.19±8.90)dC(153.19±9.34)bB(165.00±2.34)aC30(142.31±3.23)fBC(158.94±3.45)abC(148.38±3.45)eC(142.19±2.45)fC(153.19±1.34)dC(157.44±3.45)dC(148.93±2.56)eC(175.53±1.23)bB(161.54±3.45)cB(179.24±3.45)aC60(152.31±3.76)fB(178.94±9.34)abB(161.55±2.34)eB(182.67±4.34)cA(186.82±1.15)bB(210.75±3.45)aA(153.18±3.46)fC(172.10±2.34)dB(152.31±3.45)fB(208.94±7.45)abB90(186.82±2.34)deA(230.75±3.46)bA(181.62±4.36)eA(177.82±3.27)fA(199.90±5.90)dA(182.75±4.76)eB(223.47±3.25)cdA(233.63±3.20)abA(235.15±1.23)aA(225.00±3.21)cA120(159.67±2.23)fB(200.00±3.24)cA(161.50±1.09)aB(164.94±2.07)bB(158.22±2.14)bC(187.33±2.34)efB(179.08±1.34)eB(214.55±7.23)fA(205.42±3.78)cdA(206.85±7.09)dB
表3虫害诱导主梢POD活性在取食的30 d时略低于健康主梢,但差异不显著,而此时恰好为赤松梢斑螟在球果内危害的时期,其他时间段均显著高于健康主梢。不同浓度茉莉酸类化合物原位及系统喷施诱导的主梢POD活性在处理1 d时,10 mmol·L-1原位喷施茉莉酮诱导的酶活性最高,10 mmol·L-1与虫害无显著差异,并在之后的4个时间段2种处理均与虫害无显著差异。虫害诱导主梢PPO活性在处理的1~30 d与健康主梢无显著差异,其他时间段均显著高于健康主梢。10 mmol·L-1茉莉酸甲酯系统喷施及不同浓度茉莉酮系统和原位喷施诱导的PPO在处理后的1~120 d均显著高于或与虫害取食无差异。虫害诱导的主梢SOD只有在取食后的30 d与健康主梢无显著差异,其他时间段均显著高于健康主梢。测定喷施不同茉莉酸化合物诱导的主梢SOD活性,发现1 d时,茉莉酮诱导的酶活性显著高于茉莉酸甲酯诱导,并在处理的30和60 d一直保持较高水平,虽然在处理的90~120 d酶的活性有所下降,但10 mmol·L-1茉莉酮原位和系统喷施及100 mmol·L-1系统喷施诱导的酶活性仍高于虫害取食。虫害诱导的PAL活性在取食的1~120 d均显著高于健康主梢。不同浓度茉莉酸化合物诱导的PAL活性在处理后1 d显著低于虫害,但从处理的30 d开始,不同浓度茉莉酮处理诱导主梢PAL活性均高于或与虫害无显著差异。

表3 健康、虫害及不同浓度茉莉酸类化合物处理的红松主梢防御酶活性
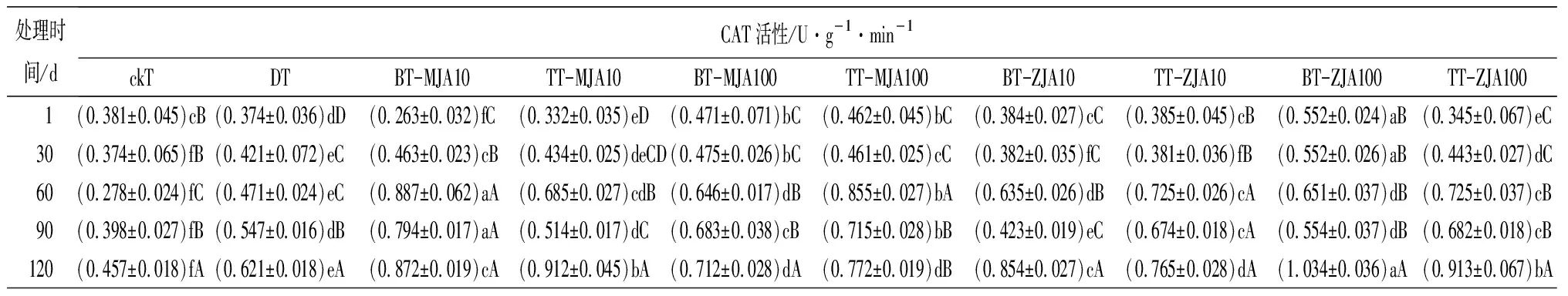
处理时间/dCAT活性/U·g-1·min-1ckTDTBT-MJA10TT-MJA10BT-MJA100TT-MJA100BT-ZJA10TT-ZJA10BT-ZJA100TT-ZJA100 1(0.381±0.045)cB(0.374±0.036)dD(0.263±0.032)fC(0.332±0.035)eD(0.471±0.071)bC(0.462±0.045)bC(0.384±0.027)cC(0.385±0.045)cB(0.552±0.024)aB(0.345±0.067)eC30(0.374±0.065)fB(0.421±0.072)eC(0.463±0.023)cB(0.434±0.025)deCD(0.475±0.026)bC(0.461±0.025)cC(0.382±0.035)fC(0.381±0.036)fB(0.552±0.026)aB(0.443±0.027)dC60(0.278±0.024)fC(0.471±0.024)eC(0.887±0.062)aA(0.685±0.027)cdB(0.646±0.017)dB(0.855±0.027)bA(0.635±0.026)dB(0.725±0.026)cA(0.651±0.037)dB(0.725±0.037)cB90(0.398±0.027)fB(0.547±0.016)dB(0.794±0.017)aA(0.514±0.017)dC(0.683±0.038)cB(0.715±0.028)bB(0.423±0.019)eC(0.674±0.018)cA(0.554±0.037)dB(0.682±0.018)cB120(0.457±0.018)fA(0.621±0.018)eA(0.872±0.019)cA(0.912±0.045)bA(0.712±0.028)dA(0.772±0.019)dB(0.854±0.027)cA(0.765±0.028)dA(1.034±0.036)aA(0.913±0.067)bA
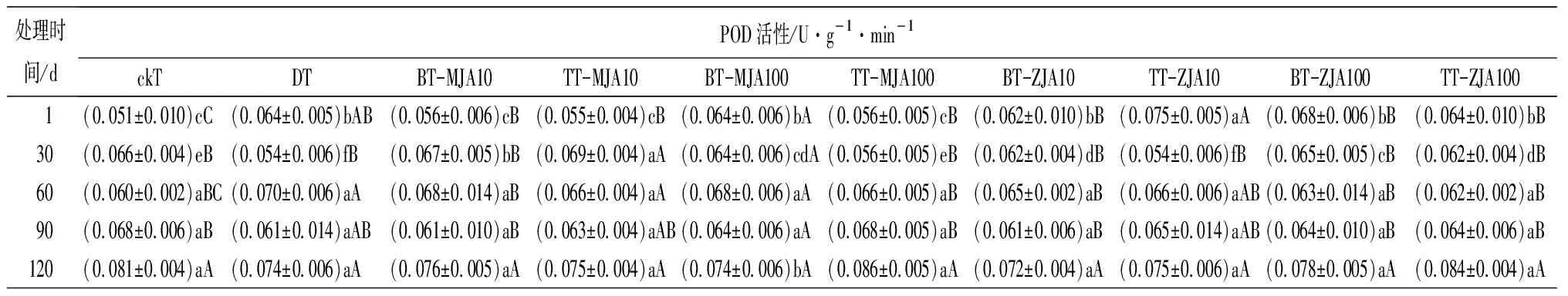
处理时间/dPOD活性/U·g-1·min-1ckTDTBT-MJA10TT-MJA10BT-MJA100TT-MJA100BT-ZJA10TT-ZJA10BT-ZJA100TT-ZJA100 1(0.051±0.010)cC(0.064±0.005)bAB(0.056±0.006)cB(0.055±0.004)cB(0.064±0.006)bA(0.056±0.005)cB(0.062±0.010)bB(0.075±0.005)aA(0.068±0.006)bB(0.064±0.010)bB30(0.066±0.004)eB(0.054±0.006)fB(0.067±0.005)bB(0.069±0.004)aA(0.064±0.006)cdA(0.056±0.005)eB(0.062±0.004)dB(0.054±0.006)fB(0.065±0.005)cB(0.062±0.004)dB60(0.060±0.002)aBC(0.070±0.006)aA(0.068±0.014)aB(0.066±0.004)aA(0.068±0.006)aA(0.066±0.005)aB(0.065±0.002)aB(0.066±0.006)aAB(0.063±0.014)aB(0.062±0.002)aB90(0.068±0.006)aB(0.061±0.014)aAB(0.061±0.010)aB(0.063±0.004)aAB(0.064±0.006)aA(0.068±0.005)aB(0.061±0.006)aB(0.065±0.014)aAB(0.064±0.010)aB(0.064±0.006)aB120(0.081±0.004)aA(0.074±0.006)aA(0.076±0.005)aA(0.075±0.004)aA(0.074±0.006)bA(0.086±0.005)aA(0.072±0.004)aA(0.075±0.006)aA(0.078±0.005)aA(0.084±0.004)aA
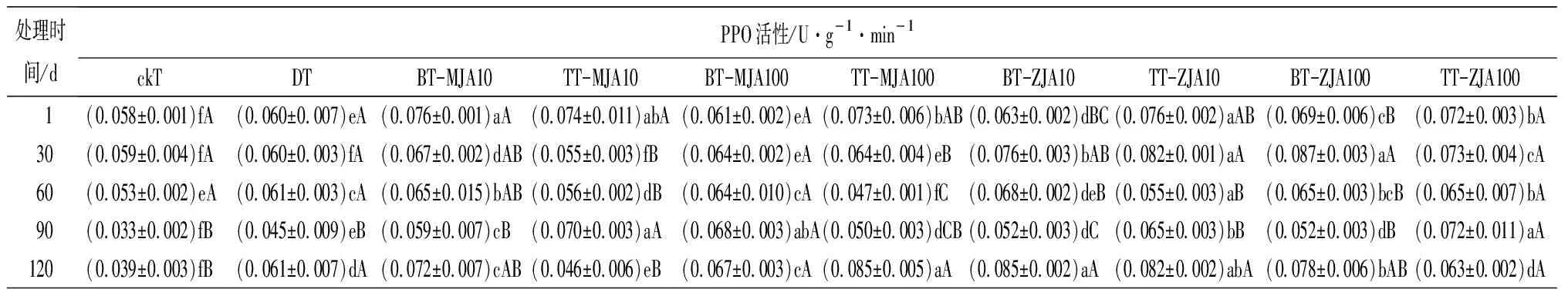
处理时间/dPPO活性/U·g-1·min-1ckTDTBT-MJA10TT-MJA10BT-MJA100TT-MJA100BT-ZJA10TT-ZJA10BT-ZJA100TT-ZJA100 1(0.058±0.001)fA(0.060±0.007)eA(0.076±0.001)aA (0.074±0.011)abA(0.061±0.002)eA(0.073±0.006)bAB(0.063±0.002)dBC(0.076±0.002)aAB(0.069±0.006)cB(0.072±0.003)bA30(0.059±0.004)fA(0.060±0.003)fA(0.067±0.002)dAB(0.055±0.003)fB(0.064±0.002)eA(0.064±0.004)eB(0.076±0.003)bAB(0.082±0.001)aA(0.087±0.003)aA(0.073±0.004)cA60(0.053±0.002)eA(0.061±0.003)cA(0.065±0.015)bAB(0.056±0.002)dB(0.064±0.010)cA(0.047±0.001)fC(0.068±0.002)deB(0.055±0.003)aB(0.065±0.003)bcB(0.065±0.007)bA90(0.033±0.002)fB(0.045±0.009)eB(0.059±0.007)cB(0.070±0.003)aA(0.068±0.003)abA(0.050±0.003)dCB(0.052±0.003)dC(0.065±0.003)bB(0.052±0.003)dB(0.072±0.011)aA120(0.039±0.003)fB(0.061±0.007)dA(0.072±0.007)cAB(0.046±0.006)eB(0.067±0.003)cA(0.085±0.005)aA(0.085±0.002)aA(0.082±0.002)abA(0.078±0.006)bAB(0.063±0.002)dA
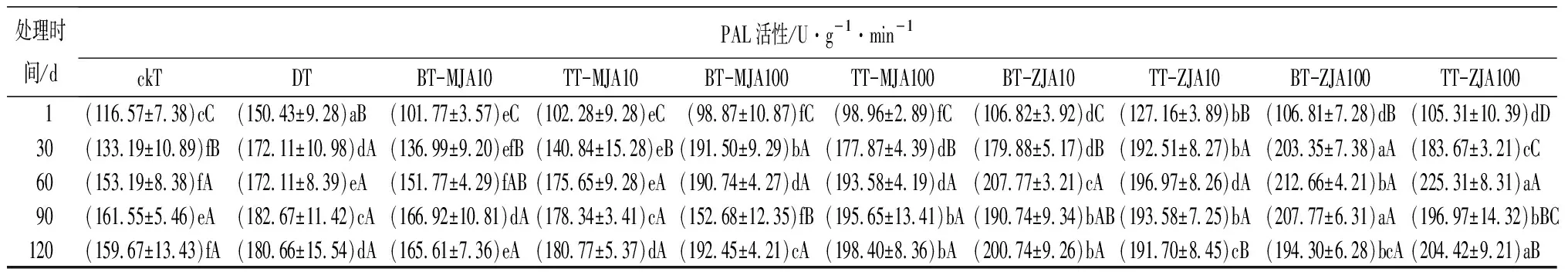
处理时间/dPAL活性/U·g-1·min-1ckTDTBT-MJA10TT-MJA10BT-MJA100TT-MJA100BT-ZJA10TT-ZJA10BT-ZJA100TT-ZJA100 1(116.57±7.38)cC(150.43±9.28)aB(101.77±3.57)eC(102.28±9.28)eC(98.87±10.87)fC(98.96±2.89)fC(106.82±3.92)dC(127.16±3.89)bB(106.81±7.28)dB(105.31±10.39)dD30(133.19±10.89)fB(172.11±10.98)dA(136.99±9.20)efB(140.84±15.28)eB(191.50±9.29)bA(177.87±4.39)dB(179.88±5.17)dB(192.51±8.27)bA(203.35±7.38)aA(183.67±3.21)cC60(153.19±8.38)fA(172.11±8.39)eA(151.77±4.29)fAB(175.65±9.28)eA(190.74±4.27)dA(193.58±4.19)dA(207.77±3.21)cA(196.97±8.26)dA(212.66±4.21)bA(225.31±8.31)aA90(161.55±5.46)eA(182.67±11.42)cA(166.92±10.81)dA(178.34±3.41)cA(152.68±12.35)fB(195.65±13.41)bA(190.74±9.34)bAB(193.58±7.25)bA(207.77±6.31)aA(196.97±14.32)bBC120(159.67±13.43)fA(180.66±15.54)dA(165.61±7.36)eA(180.77±5.37)dA(192.45±4.21)cA(198.40±8.36)bA(200.74±9.26)bA(191.70±8.45)cB(194.30±6.28)bcA(204.42±9.21)aB
2.2 健康、虫害及不同茉莉酸类化合物处理的红松球果和主梢蛋白酶抑制剂活性
表4虫害取食诱导的球果胰蛋白酶抑制剂活性在取食的1~120 d显著高于健康球果,不同浓度茉莉酮原位及系统喷施诱导的胰蛋白酶抑制剂活性显著高于(1~60 d)或与虫害无显著差异(90、120 d)。虫害取食诱导球果胰凝乳蛋白酶抑制剂活性只有在取食后60 d显著低于健康球果,其他时间二者无显著差异。不同浓度茉莉酸化合物诱导的胰凝乳蛋白酶抑制剂,除系统喷施茉莉酸甲酯30 d时诱导的抑制剂显著低于虫害外,在处理的1~120 d的其他处理均高于或与虫害无显著差异。

表4 健康、虫害及不同浓度茉莉酸类化合物处理的红松球果蛋白酶抑制剂活性

处理时间/d胰凝乳蛋白酶抑制剂活性/U·g-1·min-1ckCDCBC-MJA10CC-MJA10BC-MJA100CC-MJA100BC-ZJA10CC-ZJA10BC-ZJA100CC-ZJA100 1(12.12±0.82)fC(13.32±0.93)eB(13.48±0.74)eB(22.64±0.94)bA(14.19±0.23)dB(18.38±0.42)cA(13.15±0.980eC(12.75±0.34)fC(18.19±0.350cAB(27.29±0.78)aA30(19.54±0.87)dA(20.31±0.93)cA(19.75±0.82)dA(18.75±0.84)eAB(18.50±0.54)eA(11.75±0.43)fB(22.05±0.78)bA(24.29±0.35)aA(20.21±0.98)cA(24.23±0.45)aAB60(15.17±0.92)cdB(12.65±0.39)eB(15.75±0.34)cB(11.25±0.95)efD(15.28±0.75)dB(10.15±0.83)fB(16.15±0.89)bB(20.63±0.45)aA(20.23±0.47)abA(20.17±0.87)aBC90(12.84±0.75)fC(13.03±0.94)fB(18.75±0.78)cA(13.25±0.49)fDC(19.23±0.34)bA(19.25±0.35)bA(15.25±0.98)eBC(16.13±0.45)dB(20.15±0.78)aA(16.75±0.43)dC120(12.54±0.33)dC(12.31±0.83)dB(10.75±0.45)fC(15.75±0.78)bB(15.15±0.99)bB(11.75±0.34)eB(12.05±0.86)deC(24.16±0.78)aA(16.25±0.38)bB(14.17±0.92)cC
表5虫害诱导的主梢胰蛋白酶抑制剂活性只有在处理后120 d低于健康主梢,其他时间段均显著高于健康主梢。不同浓度茉莉酸类化合物诱导的胰蛋白酶抑制剂活性在处理后1和30 d均显著高于虫害取食。处理后60 d,只有系统喷施10 mmol·L-1茉莉酮显著低于虫害取食,处理后90 d,10 mmol·L-1茉莉酸甲酯原位喷施处理显著低于虫害取食。在120 d,各处理均显著低于或与健康主梢无显著差异。虫害取食诱导的胰凝乳蛋白酶抑制剂在取食的1~120 d均显著高于健康主梢。不同浓度茉莉酸化合物处理主梢后,主梢胰凝乳蛋白酶抑制剂活性显著增高,除处理后30 d原位喷施茉莉酸甲酯显著高于同一浓度同种化合物系统喷施外,其他时间段的系统喷施诱导的抑制剂活性均较高。

表5 健康、虫害及不同浓度茉莉酸类化合物处理的红松主梢蛋白酶抑制剂活性

处理时间/d胰凝乳蛋白酶抑制剂活性/U·g-1·min-1ckTDTBT-MJA10TT-MJA10BT-MJA100TT-MJA100BT-ZJA10TT-ZJA10BT-ZJA100TT-ZJA100 1(12.34±0.38)eB(15.67±0.33)cB(18.75±0.29)abC(13.48±0.41)dC(12.63±0.83)deC(11.92±0.94)fC(19.25±0.62)aC(15.93±0.76)cC(16.33±0.38)bB(11.75±0.78)fC30(15.34±0.72)fA(18.75±0.45)cA(18.93±0.65)dC(27.92±0.88)aA(18.95±0.67)dB(26.65±0.44)aA(20.31±0.83)bC(19.43±0.62)bB(18.26±0.82)cB(16.47±0.78)eA60(13.15±0.97)eAB(19.53±0.35)dA(22.67±0.54)cB(25.96±0.46)aB(25.32±0.43)aA(12.31±0.23)fC(23.25±0.75)bC(22.46±0.78)cB(23.73±0.71)bA(12.31±0.65)fC90(12.84±0.82)fB(15.75±0.46)dB(28.24±0.46)aA(15.27±0.78)dC(25.96±0.56)bA(14.42±0.74)eC(25.11±0.93)bcB(25.34±0.66)cA(25.05±0.78)cA(14.06±0.65)eAB120(13.03±0.98)fAB(15.34±0.34)eB(22.24±0.53)dB(28.12±0.35)aA(25.27±0.36)bA(15.15±0.53)eB(29.16±0.56)aA(23.18±0.62)cAB(24.28±0.47)cA(15.60±0.37)eAB
3 结论与讨论
植物防御酶的合成是植物防御体系形成的重要前提[16-18]。环境胁迫、损伤、昆虫取食或外源激素使植物呼吸作用加强,活性氧(ROS)的产生和代谢失去平衡,导致氧胁迫[19-22]。活性氧爆发过程中O2-·常常最先产生,SOD、CAT和POD调节氧爆发过程中产生的H2O2,协同植物诱导防御基因的表达[23],之后涉及PAL[24]和PPO[25]等催化酚类代谢的合成酶的进一步提高。
虫害、茉莉酸甲酯和茉莉酮诱导后,球果和主梢SOD、CAT和POD活性在1 d便迅速提高,说明3种处理均可诱导植物的抗性反应。SOD在抗性反应中主要是将O2-·歧化为H2O2[19]。在之后的过程中CAT可降低H2O2引起的植物表面局部性坏死;并对作为防御信号的H2O2起到调控作用,此过程可触发植物水杨酸代谢途径,外源施用水杨酸可以显著诱导H2O2水平,并抑制CAT活性[26]。在植物抗性反应中水杨酸途径与植物抗病性密切相关并与茉莉酸途径存在交互作用[27]。本研究利用茉莉酸类化合物诱导植物系统防御反应可能在一定程度上抑制了水杨酸途径,从而使CAT的活性更高。POD同样可以清除H2O2[19]。在细胞壁中它催化NADH或NADPH氧化产生O-2,O-2进一步歧化为H2O2和分子氧,H2O2和POD与细胞内酚底物发生反应,如将酪氨酸低聚物进步交联,这种作用参与细胞壁伸展蛋白的偶联形成木质素[28]。木质素被认为是抵抗病原侵入和扩展以及昆虫取食的重要防卫手段。但本研究中POD仅在1 d时活性较高,后期活性无显著增加。研究认为当参与偶联的POD降低时,细胞壁的强度降低,则会促进植物细胞膨大植株生长[29]。红松的球果和主梢均为生长较旺盛的器官,这些部位的特点即为细胞膨大或增生,因此木质素等物理障碍可能不是这些部位的主要防御手段。
PPO存在于植物质体内囊体,基因家族表达的多样性是其显著特征,保证了其能够对各种因子做出反应,以满足植物体复杂多变的生理活动[30]。创伤损伤、取食和茉莉酸甲酯均可诱导杂交杨(Populustrichocarpa×P.deltoides)PPO基因特异性表达[31]。烟草被棉铃虫(Helicoverpaarmigera)和烟夜蛾(Helicoverpaassulta)取食后,棉铃虫诱导的PPO活性显著高于烟夜蛾取食[32]。研究认为PPO主要通过与细胞内大量产生的酚底物结合,生成活性分子醌来抵御害虫,醌会将昆虫食物蛋白中的必需氨基酸烷基化,使昆虫不能利用其营养[33]。还有研究认为PPO在腺体香毛簇中与酚底物结合后产生类似“超级黏着”的物质,以捕获体型微小的昆虫[31]。Martin用茉莉酸甲酯处理云杉幼苗,6~9 d后靠近形成层的木质部细胞与周围细胞相比,细胞质变浓,细胞壁变薄,在富含萜的小腔周围形成树脂导管初生上皮细胞,15 d后小腔充满树脂[10]。本研究中茉莉酸甲酯、茉莉酮和幼虫取食诱导的PPO在处理后的1~120 d均保持较高水平。众所周知,红松在受到损伤后伤口会大量流脂,这种流脂可以黏着害虫、封闭损伤伤口避免伤口感染,因此推测这可能是红松球果和主梢防御取食和损伤的一种主要方式。植物蛋白酶抑制剂和PPO都有抗营养的作用,使昆虫营养缺乏,从而对害虫进行防御。喷施不同浓度茉莉酸类化合物使球果和主梢胰蛋白酶抑制剂活性分别在处理后的1~60 d和1~30 d显著增加,球果和主梢胰凝乳蛋白酶抑制剂活性分别在处理的1~120 d和30~120 d显著高于或与虫害取食无显著差异,且系统诱导主梢胰凝乳蛋白酶抑制剂活性显著高于原位诱导。
PAL是细胞壁酚底物形成的关键酶,它催化L-苯丙氨酸通过非氧化脱氨反应,生成反式肉桂酸和氨,为苯丙烷代谢的第一步,由此进入植物次生代谢反应,生成香豆酸、阿魏酸等一系列酚酸[24]。本研究中PAL活性也在测定的1~120 d保持较高水平。PAL和PPO活性持续时间与课题组先前对酚酸的测定结果[15]相吻合。可见茉莉酮同样能够诱导植物自身防御物质的积累。Maria利用茉莉酮处理小麦,诱导的除了反式香豆酸、丁香酸、对羟基苯甲酸、香草酸及顺式和反式阿魏酸等酚酸外,还包括多种苯并噁唑嗪酮[14]。Dabrowska et al.[34]研究发现,茉莉酮的反应生成途径包括2条:PathA通过顺式-OPDA经顺式茉莉酸异构而形成;PathB顺式-OPDA在一定条件下可被转化为反式-OPDA,再经过一系列反应形成茉莉酮,但这个驱使条件目前尚不明确。同样的反应在龙眼焦腐病菌(Lasiodiplodiatheobromae)也被发现[35]。可见,在防御反应中,植物会根据自身情况启动不同茉莉酮反应途径,以应对复杂的外部环境。
植物对茉莉酮信号的响应还存在组织特异性。Michael分别用茉莉酸甲酯和茉莉酮的雾化气体处理蚕豆(Viciafaba)植株,发现2种物质均诱导茎部防御基因D251序列表达上调,而叶部D251序列只有在茉莉酮处理的植株上才能稳定表达[13]。本研究中,红松球果和主梢经茉莉酸甲酯或茉莉酮处理后诱导的防御酶与蛋白酶抑制剂无组织特异性,可能是因为主梢和球果均属于细胞分类比较旺盛的组织器官,因此二者无显著差异。
此外,研究发现一些昆虫还将茉莉酮作为其性信息素的成分[36]。茉莉酮以及茉莉酸途径的化合物在植物病虫害防治过程中可能存在更多的潜在价值,需进一步挖掘研究。
猜你喜欢
格言·校园版(2022年17期)2022-07-06
陕西科技大学学报(2022年3期)2022-05-27
江西水产科技(2022年2期)2022-05-17
中国饲料(2022年5期)2022-04-26
浙江工业大学学报(2022年1期)2022-01-25
安徽科技(2021年10期)2021-11-12
大自然探索(2021年2期)2021-04-02
食品安全导刊·中旬刊(2020年2期)2020-06-01
高中生·青春励志(2019年4期)2019-05-09
文苑(2018年22期)2018-11-19
