
与蒙古冰草抗旱相关的NAC转录因子生物信息学及其表达分析
2021-07-05 03:49范菠菠张学峰马艳红
草地学报 2021年6期
范菠菠, 张学峰, 于 卓, 赵 彦, 马艳红*
(1. 内蒙古农业大学农学院, 内蒙古 呼和浩特 010019; 2. 内蒙古农业大学草原与资源环境学院, 内蒙古 呼和浩特 010018)
转录因子(Transcription Factors,TF)在植物抗非生物逆境中起关键作用,主要的TF家族有WKRY,bZIP,MYB,BHLH,NAC等[1-2]。其中NAC家族是植物特有的、最大的TF家族之一,具有独特的结构特征,其NAM结构域由N端150个左右的氨基酸残基组成,含有5个亚结构域A—E[3-5],其中A,C,D子域具有高度保守性,B,E子域有较大可变性,使NAC转录因子表现出多种调节功能。首个NAC基因在矮牵牛(PetuniahybridaVilm)中被发现[6],随后在拟南芥(Arabidopsisthaliana)中发现了ATAF1/2和CUC2基因[7],之后NAC基因在许多其他物种中被检测到。目前,植物转录因子数据库PlantTFDB(2021年3月17日)中收录了19 997个NAC转录因子,其中拟南芥138个、粳稻(Oryzasativasubsp.japonica)170个、籼稻(Oryzasativasubsp.indica)158个、小麦(Triticumaestivum)263个、大豆(Glycinemax)269个,NAC转录因子广泛参与植物的生长发育、逆境胁迫响应[8-13]。在干旱胁迫处理葡萄(Vitisvinifera)砧木后,qRT-PCR分析表明VvNAC8,VvNAC17,VvNAC18,VvNAC26,VvNAC40,VvNAC8,VvNAC56共7个基因与抗旱相关[12]。对甘薯(IpomoeabatatasLam.)的IbNAC72基因研究发现,干旱处理的转基因烟草根系会更加发达[14]。在干旱环境中水稻SNAC1基因的表达可提高22%~34%的结实率[8]。水稻ONAC066在干旱中起正向调控作用[15]。小黑麦(TriticosecaleWittmack)的TwNAC01基因在抗旱应激反应中起重要作用[16]。拟南芥的ANAC069[17]、水稻的ONAC095在干旱胁迫中起负调控作用[18]。水稻的OsNAC006[19],OsNAC14[20],大豆的GmNAC8[21],GmNAC019[22],GmNAC109[23]基因都对干旱胁迫有调控作用。
目前,影响植物生长发育的非生物逆境因素中干旱占首位,是造成作物产量大幅下降的主要逆境因素[24]。蒙古冰草(AgropyronmongolicumKeng)是耐旱性强的二倍体禾本科多年生牧草,具有作物遗传改良的优异抗旱基因资源[25-26]。NAC转录因子在作物干旱逆境胁迫响应中发挥重要作用,但蒙古冰草抗旱相关的NAC转录因子的研究尚属空白。本研究基于转录组数据,对蒙古冰草NAC转录因子家族成员进行鉴定,通过对NAC蛋白质结构域、理化性质、亚细胞定位、磷酸化位点、保守基序等进行分析,并与已报道具有抗旱功能的NAC蛋白进行多序列对比、构建系统进化树,以期为蒙古冰草抗旱NAC转录因子鉴定及作用机制研究奠定理论基础。
1 材料与方法
1.1 试验材料与处理
本试验材料为蒙古冰草,种子采自于内蒙古农业大学萨拉齐牧草繁种基地。挑选颗粒饱满的种子于室内培养箱(BIC-300)水培生长,培养温度为24℃,光周期为16 h,在三叶一心时期用含25% PEG-6000的1/5 Hoagland’s营养液进行模拟干旱处理,每天定期补充蒸发水分,将未干旱处理的蒙古冰草叶片设置为对照(CK)。干旱处理12 h,24 h,48 h,3 d,5 d,7 d及复水24 h(fs24 h)的蒙古冰草叶片作为处理组,在每个处理时间取蒙古冰草叶片将其迅速放入预冷的无酶冻存管中,迅速转移到—80℃冰箱进行保存;每个处理时期3次重复,提取蒙古冰草叶片总RNA,委托杭州联川生物技术股份有限公司进行转录组测序。
1.2 蒙古冰草NAC转录因子筛选及结构域鉴定
基于蒙古冰草转录组测序结果,根据功能注释初步筛选出37个NAC转录因子,用ORF Finder在线软件(https://www.ncbi. nlm.nih.gov/orffinder)对基因的开放阅读框(Open Reading Frame,ORF)进行选择,并进行结构域比对,然后再用Pfam(http://pfam.xfam.org/),CDD(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)软件进行预测验证。
1.3 蒙古冰草NAC蛋白的理化性质及亚细胞定位预测
通过Protparam(https://web.expasy.org/protparam/)[27-28]对NAC蛋白的理化性质[Isoelectric Point(PI)值,蛋白相对分子质量,不稳定指数,脂肪系数,亲疏水性以及氨基酸数目]进行分析;利用Softberry(http://linux1.softberry.com/)对NAC蛋白的亚细胞定位进行预测。
1.4 蒙古冰草NAC蛋白磷酸化位点分析
应用NetPhos 3.1 server(http://www.cbs.dtu.dk/services/NetPhos/)[29]对蒙古冰草15个NAC蛋白序列中磷酸化位点进行预测。
1.5 蒙古冰草NAC蛋白系统发育树构建
将蒙古冰草15条具有完整结构域的NAC蛋白,与从Uniprot(https://www.uniprot.org/)及NCBI(https://www.ncbi.nlm.nih.gov/)数据库挑选的57条拟南芥和具有抗旱功能的13条玉米、水稻、小麦和大豆等作物的NAC蛋白进行进化关系分析,用MEGA 7.0[30]的Muscle进行多序列比对,并基于相邻连接法(neighbor-joining)构建系统进化树,应用ITOL(https://itol.embl.de/)软件进行美化。
1.6 蒙古冰草NAC蛋白的保守基序及序列比对
利用MEME(https://meme-suite.org/meme/)软件对蒙古冰草AmNAC102-2,AmNAC100及高度同源的小麦、水稻和小黑麦的NAC蛋白进行保守基序分析,并通过DNAMAN6.0对这5条蛋白进行多序列比对。
1.7 蒙古冰草NAC蛋白二、三级结构预测
通过SOPMA[31](https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page= npsa_sopma. html)在线软件对NAC蛋白二级结构预测,分析其中的螺旋、转角、延伸链及无规则卷曲所占比例,用SWISS-MODEL[32](https://swissmodel.expasy.org/)对其进行三级结构预测。
1.8 蒙古冰草抗旱NAC基因的qRT-PCR
用SnapGene 3.2.1对筛选出的蒙古冰草抗旱相关的AmNAC100和AmNAC102-2基因的特异性引物进行设计,以U6作为内参基因(引物序列见表1),并委托生工(Sangon,上海)合成。

表1 蒙古冰草抗旱相关NAC基因qRT-PCR引物序列Table 1 The sequence of qRT-PCR primers of drought resistance related NAC gene of A. mongolicum Keng
用FastQuant RT Kit(with gDNase)试剂盒(KR106)将蒙古冰草处理组及对照组的Total RNA反转录为cDNA,用MonAmp SYBR Green qPCR Mix试剂盒(MQ10201S)进行qRT-PCR试验,每个处理4次生物学重复。反应体系为MonAmp SYBR Green qPCR Mix试剂10 μL,cDNA模板1 μL,上下游引物(10 μM)各0.5 μL,Nuclease-Free Water 8 μL。反应程序为预变性95℃,30 s,变性95℃,10 s,退火延伸62℃,30 s,循环数为45,在FTC-3000P上运行程序。
用livak等的2-ΔΔCT法[34]计算蒙古冰草2个NAC基因的相对表达量,用SPSS 26.0软件进行差异显著性分析。
2 结果与分析
2.1 蒙古冰草NAC转录因子家族的鉴定分析
从转录组数据库挑选出注释为NAC转录因子的37个基因,通过ORF分析和结构域预测,获得具有NAC蛋白结构域的26个基因,其中11条NAM结构域存在残缺现象,14条NAC蛋白的NAM结构域在150个氨基酸残基左右,为完整的NAM结构域;只有一条AmNACA2蛋白预测为完整的NAC-NACA结构域(图1)。
2.2 蒙古冰草NAC蛋白理化性质和亚细胞定位分析
通过Protparam对26个NAC蛋白进行理化性质分析,发现NAC蛋白氨基酸数目在60~432之间,相对分子量在7 080.09~47 615.03之间;PI值为4.24~10.48,表明既有酸性蛋白也有碱性蛋白;不稳定指数在31.92~71.92之间,脂肪系数介于57.10~77.83,NAC蛋白的疏水性为—0.780~-0.257,表明所有蒙古冰草NAC蛋白均为亲水性蛋白(表2)。
通过Softberry软件进行亚细胞定位分析,发现AmNACA2,AmNAC1-2蛋白定位在线粒体,AmNAC046定位于细胞质,其余蛋白均定位于细胞核,说明NAC蛋白主要在细胞核中行使功能,也有少数在其他细胞器中发挥作用。
2.3 蒙古冰草NAC蛋白磷酸化位点分析
蛋白磷酸化是植物中普遍存在的一类现象,出现在植物蛋白激酶及信号转导过程中,属于蛋白翻译后修饰中的一种[31-32]。对蒙古冰草的15个NAC蛋白进行磷酸化位点分析,发现其均含有丝氨酸、苏氨酸以及酪氨酸,其中磷酸化位点总数最多的是AmNAC103蛋白(41个),最少的是AmNAC036-4蛋白(8个)。

图1 蒙古冰草NAC家族序列结构域Fig.1 Sequence domain of A. mongolicum Keng NAC family

表2 蒙古冰草NAC蛋白理化性质及亚细胞定位Table 2 Subcellular localization and physicochemical properties of NAC protein of A. mongolicum Keng

表3 蒙古冰草NAC蛋白磷酸化位点预测Table 3 The predicted phosphorylation sites of NAC proteins from A. mongolicum Keng
2.4 蒙古冰草NAC蛋白进化关系分析
为研究蒙古冰草NAC蛋白家族的进化关系,绘制系统进化树,蛋白名标签分为6种颜色,将进化树分为Ⅰ~Ⅵ亚族。进化树枝的颜色从绿色到红色过渡,代表同源性从0到1。纵观整个进化树,除Ⅳ和Ⅴ亚族不含蒙古冰草NAC蛋白外,其余亚族中均有分布,说明Ⅳ和Ⅴ中的拟南芥蛋白与蒙古冰草NAC蛋白无进化关系。AmNACA2蛋白单独成一个分支,AmNAC073处于Ⅱ亚族与拟南芥AtNAC73,AtNAC10等在一个小支上。有7个蒙古冰草NAC蛋白处于第Ⅲ亚族,分别为AmNAC103,AmNAC071,AmNAC090-1,AmNAC090-2,AmNAC042,AmNAC036-1和AmNAC036-4。其余蒙古冰草蛋白都处于第Ⅵ亚族,其中AmNAC2-1与拟南芥AtNAC50在同一支上;AmNAC100与小黑麦的抗旱蛋白TwNAC01[16]同源性极高;AmNAC041与拟南芥AtNAC83处于同一分支;AmNAC2-2,AmNAC047-1与AtNAC47有较近进化关系;AmNAC102-2蛋白与抗旱功能已知的小麦TaNAC6[34]和水稻OSNAC6[35],OsNAC5[36]同源性较高。由此推测AmNAC100和AmNAC102-2参与干旱胁迫调控。
2.5 蒙古冰草抗旱相关的NAC蛋白二、三级结构分析
对AmNAC100和AmNAC102-2进行蛋白结构预测分析,结果表明AmNAC100蛋白二级结构中无规则卷曲占51.26%,α-螺旋占25.49%,延伸与β-转角分别占16.25%,7.00%;AmNAC102-2蛋白中无规则卷曲占58.62%,α-螺旋占13.10%,延伸与β-转角分别占20.69%,7.59%;TaNAC6中无规则卷曲占67.11%,α-螺旋占18.12%,延伸和β-转角分别占11.07%,3.69%;水稻OsNAC5中无规则卷曲占60.49%,α-螺旋占22.19%,延伸和β-转角分别占12.46%和4.86%;小黑麦中TwNAC01无规则卷曲占53.98%,α-螺旋占22.73%,延伸和β-转角分别占16.48%,6.82%(图3)。由以上数据可知,蒙古冰草的2个抗旱相关NAC蛋白的二级结构主要以无规则卷曲为主,其次含α-螺旋、延伸和转角。
NAC蛋白三级结构分析结果发现AmNAC100与TwNAC01空间构型一致;AmNAC102-2与TaNAC6,OsNAC5蛋白空间构型保持一致(图4),与蛋白进化树中同源关系相一致。
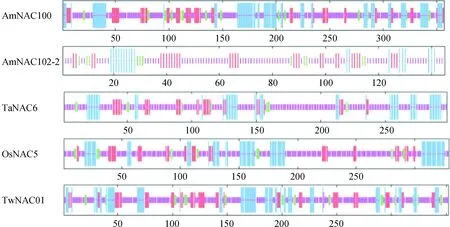
图3 NAC蛋白二级结构Fig.3 Secondary structure of NAC protein注:蓝色,α-螺旋;紫色,无规则卷曲;红色,延伸;绿色,β-转角Note:Bue,alpha-helix;purple,random coil;red,extended strand;green,beta turn

图4 NAC蛋白三级结构Fig.4 Tertiary structure of NAC protein
2.6 蒙古冰草抗旱相关NAC蛋白基序及保守结构域分析
保守基序分析发现,5个NAC蛋白输出的有效motif有8个,均含有motif 1,2,3和8(图5),说明这4个motif是蒙古冰草AmNAC100和AmNAC102-2蛋白的共有基序。AmNAC102-2蛋白不含motif4,其motif 1,2,3和8与TaNAC6,OsNAC5一致;AmNAC100与TwNAC01都含有motif5,6,7,这两个蛋白基序结构一致。
将蒙古冰草AmNAC100,AmNAC102-2与小麦、水稻和小黑麦的已知抗旱功能NAC蛋白进行多序列比对发现,其保守结构域均分为A—E亚结构域,其中D,E亚结构域是重要的DNA结合区域,A亚结构域在NAC二聚体蛋白形成中发挥作用。

图5 抗旱相关NAC蛋白基序Fig.5 The motif of NAC protein related to drought resistance

图6 保守结构域的多序列比对Fig.6 Multiple sequence alignment of conserved domains
2.7 蒙古冰草2个抗旱相关的NAC基因的表达分析
蒙古冰草抗旱相关的AmNAC100和AmNAC102-2在25% PEG-6000处理期间表达量均高于CK。其中AmNAC100基因干旱处理12 h,3 d以及复水24 h的相对表达量分别是CK的15.37,18.70和14.65倍(P<0.05),只有干旱处理24 h后AmNAC100基因相对表达量与CK差异不显著;AmNAC102-2基因相对表达量较高的处理组有12 h,48 h,3 d及复水24 h,分别是CK的7.34,3.67,4.04以及7.12倍(P<0.05)(图7),表明蒙古冰草的2个NAC基因在25% PEG-6000 模拟干旱处理中均可被诱导表达。

图7 25% PEG-6000 干旱胁迫下蒙古冰草抗旱相关的NAC基因相对表达量Fig.7 Relative expression of drought resistance related NAC genes in Agropyron mongolicum Keng under 25% PEG-6000 drought stress注:不同小写字母表明差异显著(P<0.05)Note:Different lowercase letters indicate significant difference at the 0.05 level
3 讨论
为筛选与蒙古冰草抗旱相关的NAC转录因子,本研究基于转录组测序数据筛选出26个蒙古冰草NAC家族成员,其中15个具有完整NAC转录因子结构域。生物信息学分析发现蒙古冰草NAC转录因子均属于亲水性蛋白,与大豆、苦荞、苹果等[32,37-38]对NAC蛋白亲疏水性的研究结果一致。亚细胞定位发现蒙古冰草NAC蛋白主要定位于细胞核中,有2个定位于线粒体中、1个在细胞质中,说明大部分蛋白在细胞核中发挥作用,少数在其他细胞器中行使功能。
大量研究表明NAC蛋白在非生物胁迫中起正调控作用,比如IbNAC72[14],SNAC1[8],ONAC066[15]等。将15个NAC蛋白与拟南芥和已知抗旱功能的13个NAC蛋白构建系统进化树,预测到2个抗旱相关的NAC转录因子,其余蒙古冰草NAC蛋白分布于Ⅰ,Ⅱ,Ⅲ和Ⅵ亚族中。处于同亚族的蒙古冰草NAC基因与其他物种的NAC基因同源性存在差异,推测是由环境和物种间差异引起的,这与周棋赢、黄文娟和荣玉萍的研究一致[5,37,39]。磷酸化位点预测结果与方志红研究结果一致,均含有丝氨酸、苏氨酸及酪氨酸磷酸化位点[32]。二、三级结构预测发现蒙古冰草AmNAC100与TwNAC01结构相似,AmNAC102-2与TaNAC6,OsNAC5相似,推测其有相同的抗旱功能。
保守结构域承载着基因的重要遗传信息,对其发挥特定调控功能具有重要意义[5,40],拥有相同的保守结构域可能发挥相似的功能。根据Hisako等人对NAC蛋白保守结构域的划分,将5个抗旱相关NAC蛋白的保守结构域分为A—E亚结构域[41],通过颜色来区分NAC蛋白序列的保守关系,黑色区域表示序列相似性在100%即完全相同,粉紫色代表相似性≥75%,浅蓝色表示相似性在50%及以上,黄色表示相似性在33%以上,无色代表相似性在0~33%。通过着色可以清晰的看到保守性从高到低依次为A>C>D>B>E,高保守性结构域用来维持NAC基因特定功能,有研究表明保守的NAC结构域承担DNA结合的功能[41];子域B和E的保守性相对较小,是可变动的结构域,推测其可用来激活调控植物其他功能[41]。
王婉妮对干旱胁迫下3种葡萄砧木进行实时荧光定量分析发现,NAC基因相对表达量均高于对照[12];张欢等对甘薯抗旱基因IbNAC72的qRT-PCR发现,其在20% PEG-6000处理中相对表达量均高于对照,起正调控作用[14],这与本研究结果相似。本试验中qRT-PCR分析表明,经过生物信息学筛选得到蒙古冰草2个抗旱相关的AmNAC100和AmNAC102-2基因,在25% PEG-6000干旱处理下其相对表达量上升,呈上调表达。
4 结论
本研究首次从蒙古冰草转录组测序结果中筛选鉴定出26个NAC转录因子,得到15个有完整结构域的NAC蛋白,通过进化关系比对及保守基序,保守结构域和NAC蛋白二、三级结构分析,获得2个抗旱相关的AmNAC100和AmNAC102-2基因,对其进行差异表达分析发现其在干旱胁迫中起正调控作用。
猜你喜欢
国际放射医学核医学杂志(2020年4期)2020-07-27
阅读与作文(小学高年级版)(2019年8期)2019-10-16
食品与生活(2019年11期)2019-05-15
蒙古学问题与争论(2019年0期)2019-03-29
西域历史语言研究集刊(2018年0期)2018-11-09
草原与草业(2018年4期)2018-03-20
草原歌声(2017年1期)2017-04-23
上海农业学报(2017年3期)2017-04-10
中国工程咨询(2016年1期)2016-02-14
食品与健康(2015年8期)2015-08-11
