
深圳市红树林典型群落的物种组成及结构多样性研究
2021-08-20 03:01关开朗张信坚谭广文曾凤易慧琳廖文波
生态科学 2021年4期
关开朗, 张信坚, 谭广文, 曾凤, 易慧琳, 廖文波
深圳市红树林典型群落的物种组成及结构多样性研究
关开朗1, 张信坚1, 谭广文1, 曾凤1, 易慧琳1, 廖文波2,*
1. 广州普邦园林股份有限公司, 广州 510600 2. 中山大学生命科学学院, 广州 510275
在深圳市红树林主要分布区设置8个调查样地, 以调查深圳市红树林典型群落的物种组成和生物多样性特征。结果表明: (1)秋茄为深圳市天然红树林的建群种或优势物种; 白骨壤及海漆是天然红树林的重要组成物种; 海桑及无瓣海桑是人工林的主要组成物种; 桐花树及老鼠簕是林下灌木层以及低发展程度的红树林群落的主要组成物种。(2)海桑、无瓣海桑、海漆及白骨壤等真红树植物丰富了以秋茄为建群种、优势种或伴生种的深圳市红树林的物种种类、垂直结构和径级结构, 从而提高了红树林的物种多样性和结构多样性, 使林分更加稳定性。在对红树林进行质量提升或营建红树林人工林时, 可以通过增加本土红树植物的种类和提高林分混交程度和结构多样性, 来提高红树林的生态稳定性和景观观赏价值。
红树林; 物种组成; 物种多样性; 结构多样性; 深圳
0 前言
红树林是分布于热带和亚热带地区潮间带的以红树植物为主体的常绿群落, 具有消浪护岸、净化环境污染、维护生物多样性等作用[1]。我国的红树林主要分布在东南沿海滩涂, 包括海南、广东、广西、福建、台湾、香港和澳门[2]。在广东, 南至雷州半岛, 北达饶平和南澳岛, 沿海滩涂均有红树林间断分布[3]。
红树林是深圳市植被的重要组成部分[3]。历史上, 红树林曾经广泛分布于深圳海岸潮间带, 如保安沙井至西乡一带沿海滩涂的红树林分布面积曾达到113.4 hm2(1988年), 深圳湾红树林分布面积达79.56 hm2(1979年), 坝光沿海滩涂有大面积的红树林约200 hm2[4–5]。自改革开放以来, 随着深圳市城市建成区扩张、填海造陆、水库建设、病虫害等原因, 红树林生态系统受到极大破坏, 分布面积减少至原来的1/4[6]。目前, 深圳市的红树林总面积约169.7 hm2, 主要分布在宝安区的、南山区、福田区和大鹏新区的海岸、港湾、河口湾等周期性受淹水域中。其中, 福田区红树林包括天然林和人工林, 宝安区和南山区的红树林以人工林为主, 大鹏新区的红树林以天然林为主[7]。为更好地对深圳市红仅存的树林进行保护, 已有不少学者和单位通过人工造林、防治病虫害、消除有机污染物等措施, 对退化红树林进行生态修复[8–11], 深圳市的红树林分布面积得到一定程度的增加, 林分质量得到一定程度的提升。本研究在对深圳市红树林全面考察的基础上, 对红树林典型群落的物种组成、群落结构等做深入调查, 并对其物种多样性和结构多样性进行分析和比较, 了解物种组成及群落结构对群落生物多样性的影响, 为深圳市对红树林进行质量提升和营造红树林人工林等提供理论支撑。
1 材料与方法
1.1 样地设置与调查方法
在深圳市主要红树林分布区进行路线考察, 记录优势红树植物群落的组成、分布以及红树林内植物的种类。在福田红树林保护区、大鹏坝光、大鹏东涌河口3个天然红树林分布区, 根据优势红树植物群落的类型和分布情况进行选点并设置8个调查样方(表1)。样方面积和调查方法参考《湿地生态系统定位观测技术规范》[12], 每个调查样方大小为20 m×20 m, 分为4个10 m×10 m。植物调查指标包括: 种名、胸径(乔木)、树高、冠幅、盖度(灌木、草本、藤本)、数量。
1.2 数据处理
1.2.1 物种优势度
物种的优势度以重要值表示, 计算方法:

1.2.2 群落结构
采用韦霄等[13]的方法, 按5 m一个高度划分片层, 对高度区间内的植株数量及所有物种的优势度进行统计, 分析其垂直结构特征; 按5 cm一个等级划分胸径级别, 比较不同径级的植株数量和优势度, 分析群落径级结构特征。

表1 深圳市红树林典型群落调查样地情况
1.2.3 群落多样性的测度方法
(1)林分物种多样性
采用Shannon-Wiener多样性指数、Simpson优势度指数以及基于Shannon-Wiener指数的均匀度指数来评价群落的物种多样性[14]。
Shannon-Wiener多样性指数
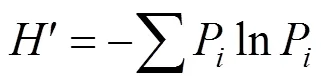
Simpson优势度指数

均匀度指数

(2)林分结构多样性
采用LEI等[15]方法, 通过测定群落物种直径大小多样性和物种多样性的二维联合熵以及物种直径大小多样性和树高多样性的二维联合熵, 来表示林分结构多样性。
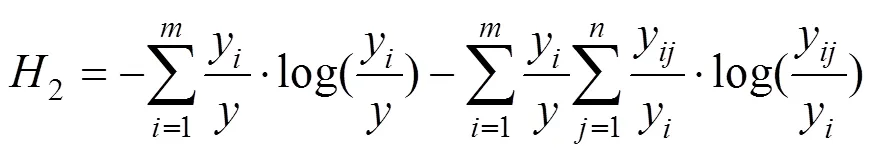
2 结果与分析
2.1 深圳市红树林典型群落的物种组成及优势度
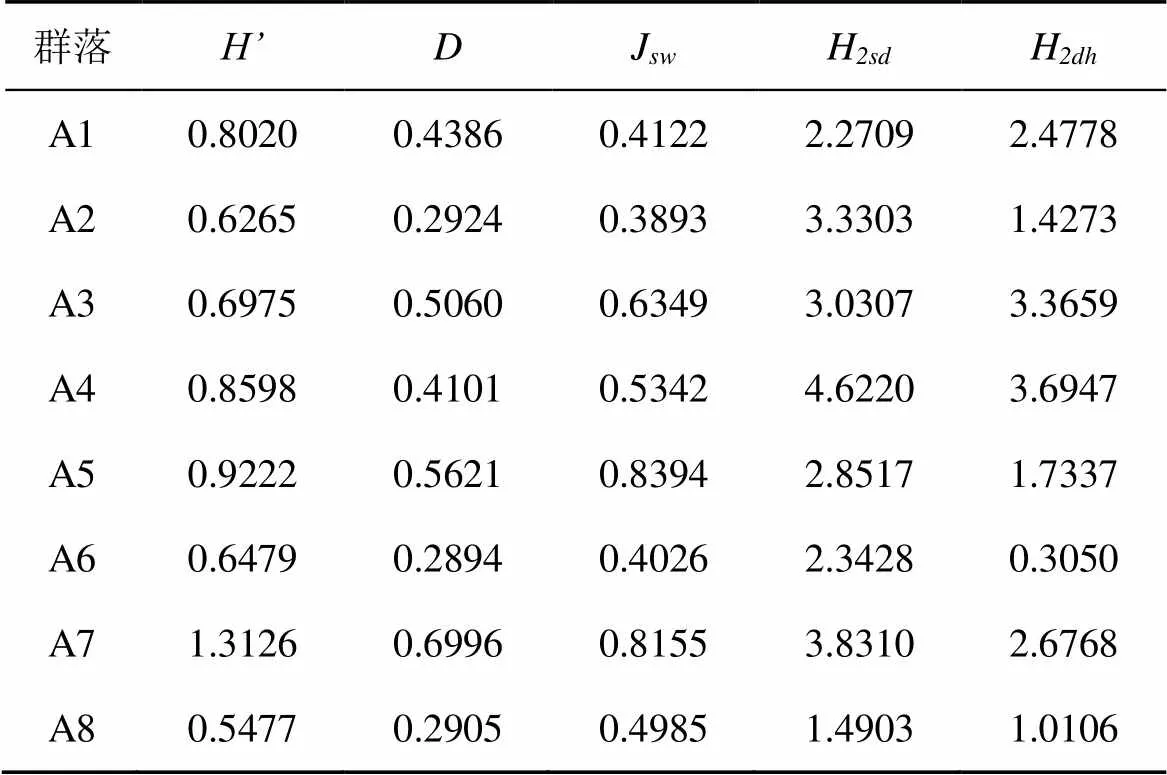
福田红树林典型红树群落乔木层组成物种主要有秋茄、白骨壤、无瓣海桑、海桑及木榄, 形成分别以秋茄、白骨壤、无瓣海桑、海桑中的一种或几种为优势种的红树群落。从重要值来看, 秋茄的重要值区间范围为21.5—100, 表明秋茄主要作为建群种、优势种或伴生种, 广泛分布于福田红树林各群落内, 其种群年龄结构主要以平均高度从3.8 cm至7.7 cm不等的壮树(2.5 cm 大鹏东涌的红树林为自然林, 虽然由于靠近东涌村而受到长期的人为干扰, 但总体上还是保留了比较完整的红树林群落。根据路线调查结果, 东涌红树林潮间内带真红树植物种类主要有海漆、秋茄、桐花树、老鼠簕、木榄及白骨壤。真红树植物群落共有两类, 分别是以海漆为特征种的混合群落及以秋茄为特征种的混合群落, 还有沿着河涌上游小面积分布的桐花树、老鼠簕群落。如表2所示, 东涌红树林真红树群落乔木层以秋茄和海漆为建群种或优势种, 重要值区间范围为11.6—66.1。其中, 秋茄主要以壮树为主, 多分枝, 平均胸径仅2.8—6.6 cm, 平均高度仅1.8—6.3 m, 表明秋茄种群总体年龄较小; 海漆主要以壮树为主, 平均胸径5.8—8.0 cm, 平均高度2.3—7.3 cm, 种群总体年龄适中; 木榄在东涌红树林零星分布, 局部形成优势种, 其种群也主要以壮树为主。灌木层主要有老鼠簕和桐花树, 两者优势度相近, 但在河中心位置, 桐花树优势明显, 可能与潮位低有关。 表2 深圳市红树林典型群落的物种组成及优势度 大鹏坝光红树林是大鹏半岛现存面积最大的红树林, 真红树植物主要有白骨壤、秋茄、桐花树、老鼠簕及海漆。真红树群落类型主要以白骨壤群落为主, 其次为小面积的与白骨壤群落混交的秋茄人工群落。典型的白骨壤群落分布于盐灶村入口滩涂。相对于深圳其他区域的红树林而言, 该白骨壤群落物种组成较为单调, 仅包括白骨壤、秋茄、桐花树3种。其中, 白骨壤在乔木层占优势, 重要值达78.4; 秋茄次之, 重要值为21.6。从胸径及高度来看, 白骨壤及秋茄的平均胸径为2.2—6.9 cm, 平均高度为2.3—2.8 m, 表明白骨壤和秋茄的种群年龄均偏小。灌木层稀疏, 仅见桐花树, 数量十分稀少。 2.2.1 群落垂直结构特征 群落的垂直结构主要指群落的分层现象, 群落中各种种群的个体在空间不同层次上分布的象, 也叫成层性[16]。一般而言, 生境条件越优或者发育阶段越老的群落, 层次结构越多, 群落也就越高[13]。 对深圳市红树林8个典型群落乔木层的垂直结构进行分析, 结果见图1。在福田红树林, 秋茄–桐花树+老鼠簕群落(A1)分为两个片层, 10—15 m片层仅见少量秋茄, 5—10 m片层全部由秋茄组成, 5 m以下片层主要由桐花树及少量的老鼠簕组成, 偶见无瓣海桑、白骨壤及短叶茳芏, 层间植物还有少量鱼藤(图1a)。秋茄+白骨壤–老鼠簕+桐花树群落(A2)乔木层明显可分为两个片层, 5—10 m片层主要由秋茄、白骨壤的成年植株组成, 且植株数量与优势度较高; 5 m以下片层主要由少量的秋茄、白骨壤的小树以及林下的密集的桐花树、老鼠簕组成。无瓣海桑–老鼠簕群落(A3)和海桑–老鼠簕群落(A4)的乔木层群落整体高度一般高于10 m, 且明显分为四个片层, 各片层的植株数量与优势度比较接近。15 m以上片层全部由海桑或无瓣海桑组成, 10—15 m片层主要由海桑或无瓣海桑组成, 还包括少量的秋茄; 5—10 m片层主要由秋茄以及部分海桑和无瓣海桑小树组成; 5 m以下片层主要由秋茄小树、海桑和无瓣海桑幼苗以及老鼠簕组成, 整体优势度略高于其他片层(图1c–d)。(秋茄)–桐花树+老鼠簕群落(A5)明显分为两个片层, 5—10 m主要由秋茄和少量桐花树组成, 5 m以下片层主要由大部分桐花树以及老鼠簕组成(图1e)。 与福田红树林相比, 大鹏东涌红树林、坝光红树林的群落乔木层垂直结构比较简单。群落整体高度低于10 m, 个别海漆植株高度可超过15 m。其中, 东涌红树林的秋茄–桐花树群落(A6)高度低于5 m, 群落乔木层分层不明显, 秋茄、海漆、木榄组成了低矮的乔木层, 少量的桐花树、老鼠簕组成了稀疏的林下灌草层(图1f)。海漆–老鼠簕+桐花树群落(A7)的结构稍复杂, 大致可分为四个片层。15 m以上片层仅见海漆, 植株数量及优势度低; 10—15m片层由少量海漆及秋茄组成, 优势度较低; 5—10 m片层主要由海漆及少量秋茄组成, 植株数量及优势度远大于其他片层; 5 m以下片层主要由少量的海漆及秋茄小树组成, 优势度相对较低(图1g)。坝光红树林的白骨壤–桐花树群落整体高度低于10 m, 群落垂直结构比较简单。优势片层为5 m以下片层, 主要由白骨壤及少量秋茄小树组成; 5—10 m片层包括白骨壤及秋茄, 植株数量少, 优势度低(图1h)。 注: a. 秋茄–桐花树+老鼠簕群落; b. 秋茄+白骨壤–老鼠簕+桐花树群落; c. 无瓣海桑–老鼠簕群落; d. 海桑–老鼠簕群落; e. (秋茄)–桐花树+老鼠簕群落; f. 秋茄–桐花树群落; g. 海漆–老鼠簕+桐花树群落; h. 白骨壤–桐花树群落。 Figure 1 Vertical structures of arbor layers of typical mangrove communities in Shenzhen 2.2.2 群落径级结构特征 按许格希等[17]方法, 对深圳市典型红树林群落的优势物种作径级结构分析。结果显示, 福田红树林秋茄–桐花树+老鼠簕群落(A1)、秋茄+白骨壤–老鼠簕+桐花树群落(A2)及(秋茄)–桐花树+老鼠簕群落(A5)主要以小径级(d<5 cm, 下同)及中径级(5 cm≤ d<15 cm, 下同)植株为主, 表明上述群落正分别处于幼龄林(图2a)以及中林龄阶段(图2b, 2e)。无瓣海桑–老鼠簕群落(A3)和海桑–老鼠簕群落(A4)各径级植株数量与优势度比较一致, 表明群落正处于由中龄林向成熟林过渡阶段, 同时, 林下幼苗或小树的数量和优势度也较大, 显示出群落良好的更新状况(图2c–d)。(秋茄)–桐花树+老鼠簕群落(A5)的植株径级主要集中以中径级为主, 少量植株胸径小于5 cm或者大于15 cm(图2e)。大鹏东涌红树林的秋茄–桐花树群落(A6)主要以小径级为主, 植株数量及优势度占绝对优势, 中径级和大径级(d≥15 cm, 下同)的植株数量较少, 优势度较低, 显示出群落正处于幼龄林阶段, 群落发展空间较大(图2f); 海漆–老鼠簕+桐花树群落(A7)主要以及中径级为主, 小径级植株数量少于中径级, 远多于大径级, 但优势度仅略大于大径级, 表明该群落正处于由中龄林向成熟林过渡的阶段, 有一定的林分更新能力(图2g)。与东涌红树林群落类似, 大鹏坝光的白骨壤–桐花树群落同样以小径级和中径级植株为主, 大径级植株数量较少, 同样处于由中龄林向成熟林过渡的阶段(图2h)。 2.3.1 群落物种多样性 Shannon-Wiener 物种多样性指数、Simpson物种多样性指数及基于Shannon-Wiener物种多样性指数的均匀度指数是反映群落中物种种类与个体数量、优势种的组成及其均匀性的综合指标[14]。对深圳市8个典型红树林群落进行物种多样性及均匀度进行分析, 结果如表3所示。与其他群落相比, 大鹏东涌的海漆–老鼠簕+桐花树群落(A7)的物种多样性指数、生态优势度指数及均匀度指数均最高, 表明该群落具有最高物种多样性, 群落中优势物种最多, 且群落物种的分布最均匀。福田红树林的(秋茄)–桐花树+老鼠簕群落(A5)的生物多样性指数、生态优势度指数及均匀度指数均高于福田红树林其他典型群落, 但低于大鹏东涌的海漆–老鼠簕+桐花树群落(A7)。大鹏坝光的白骨壤–桐花树群落的物种多样性指数及优势度指数均最低(A8), 均匀度指数略高于福田红树林的秋茄–桐花树+老鼠簕群落(A1)、秋茄+白骨壤–老鼠簕+桐花树群落(A2)及大鹏东涌的秋茄–桐花树群落(A6), 表明该群落物种多样性最低, 群落优势物种单一, 物种分布比较集中。整体而言, 福田红树林整体上显示出较高的物种多样性, 优势种比较突出, 群落物种分布比较均匀, 这可能与福田红树林处于长期的保护, 恢复程度较好有关。位于东涌河中心高地的海漆–老鼠簕+桐花树群落(A7)与位于河岸的秋茄–桐花树群落(A6)在物种多样性、优势度及均匀度方面呈现两极分化的情况, 可能是由于群落位置的不同导致受干扰的程度不同有关。坝光红树林由于长期受到人为干扰, 导致物种组成简单, 群落发展程度低, 所以物种多样性、群落优势度与均匀度均较低。 注: a.秋茄–桐花树+老鼠簕群落; b.秋茄+白骨壤–老鼠簕+桐花树群落; c.无瓣海桑–老鼠簕群落; d.海桑–老鼠簕群落; e.(秋茄)–桐花树+老鼠簕群落; f.秋茄–桐花树群落; g.海漆–老鼠簕+桐花树群落; h.白骨壤–桐花树群落。 Figure 2 Diameter structures of arbor layers of typical mangrove communities in Shenzhen 2.3.2 群落结构多样性 Shannon-Wiener多样性指数、Simpson多样性指数及的均匀度指数均从物种多样性角度对群落生物多样性进行定量描述, 不能反映出群落结构对群落生物多样性的影响。而研究表明, 提高林分结构多样性和复杂性, 可以维持和增加林分的生物多样性[18]。LEI等[15]认为, 基于联合熵的多维多样性指数可以代表林分结构的多个组成部分, 是描述林分总体结构多样性的好方法。本研究以5 cm为一个胸径等级, 以5 m为一个高度等级, 对深圳市红树林典型群落的结构多样性进行分析。结果显示, 各群落的结构多样性大小顺序与物种多样性大小顺序差异较大, 主要体现在具有较丰富的物种组成、径级结构和层次结构的群落, 结构多样性一般较大, 如福田红树林的海桑–老鼠簕群落(A4)、无瓣海桑–老鼠簕群落(A3)及东涌的海漆–老鼠簕+桐花树群落(A7)。福田红树林的秋茄+白骨壤–老鼠簕+桐花树群落(A2)具有较丰富的物种和径级结构, 但垂直结构简单(表2, 图1b), 故树种和林木大小的总体多样性较高而林木大小和树高的总体多样性较低(表3)。(秋茄)–桐花树+老鼠簕群落(A5)的物种多样性、优势度及均匀度较高, 但林木大小和树高的总体多样性较低(表3), 原因是该群落具有较大的植株密度和较多的优势种, 但群落物种较少且垂直结构简单(表2, 图1e)。与其他群落相比, 秋茄–桐花树+老鼠簕群落(A1)因物种组成、优势种数量与分布、垂直结构和径级结构比较均匀, 因而结构多样性指数处于比较处于居中位置。东涌的秋茄–桐花树群落(A6)与坝光的白骨壤–桐花树群落(A8)的结构多样性较低, 与这两个群落的物种组成、径级和层次结构均较少有很大关系。 表3 深圳市红树林典型群落多样性指数 深圳市红树林主要以本土少数几种真红树植物以及引种真红树植物形成的单优或者混合群落为主, 在植被类型与种类组成上与淇澳岛红树林保护区[19]、惠州红树林保护区[20]等珠三角地区的红树林类似。本研究调查的8个群落反映出了秋茄、白骨壤、桐花树、老鼠簕、海漆、海桑、无瓣海桑及木榄这几种真红树植物在深圳市红树林的物种组成和群落结构上不同的地位。由表2可以看到, 秋茄是上述调查的全部红树植物群落的建群种、优势种或伴生种, 一定程度上说明了秋茄是深圳市红树林的建群种或优势种。白骨壤在福田红树林中潮位区域形成局部优势群落, 以及是大鹏坝光红树林的建群种, 在深圳市天然红树林物种组成及群落结构的重要性上仅次于秋茄。海漆在东涌河口成规模分布并且是优势种, 在福田红树林仅小面积分布, 在深圳其他区域零散分布, 因此是深圳市红树林的重要组成物种之一。海桑及无瓣海桑是深圳市红树林人工林的主要组成物种, 主要集中分布在福田红树林及宝安西湾红树林公园内, 在群落内的优势度非常明显, 甚至形成单优群落, 在一些入海河口也有零星分布。桐花树及老鼠簕是红树林林下灌草层主要组成物种, 两者在低、中、高潮位均有分布, 无明显分布规律。在一些乔木层发达, 郁闭度较高的林分内, 桐花树、老鼠簕的多度均较低, 可能与这两个物种的生长需要一定强度的光照有关[21–23]。 以物种多样性指数、优势度指数及均匀度指数为代表的一维熵在一定程度上反映出了群落物种组成、植株密度及混交程度的差异。群落中优势物种的种类越多, 且在群落中的分布越均匀, 生物多样性越高, 代表性群落为大鹏东涌的海漆–老鼠簕+桐花树群落。群落中的优势物种越少, 且在群落中的分布越集中, 生物多样性越低, 代表性群落为大鹏坝光的白骨壤–桐花树群落。也就是说, 群落中更多的优势物种组成以及更高的混交程度, 意味着更高的物种多样性(表2、表3)。但仅从物种多样性的角度来评价群落生物多样性显然是不够充分的。同样, 仅从群落的径级结构多样性或层次结构多样性来评价群落生物多样性同样也是有缺陷的[15]。物种与胸径、胸径与高度的二维联合熵可以综合反映出群落的物种组成、垂直结构和径级结构的复杂程度, 是一个能够很好衡量群落结构多样性的指标。从本研究分析结果来看, 以海桑、无瓣海桑为优势种的人工林, 由于高大乔木海桑和无瓣海桑的存在, 使得林分具有较丰富的径级结构和垂直结构, 林分的结构多样性较高, 群落也就比较稳定。与此类似, 东涌红树林的海漆–老鼠簕+桐花树群落也因海漆的存在, 群落的结构多样性也均较高。 从以上结果来看, 通过增加红树林物种种类, 来提高以秋茄为建群种或优势种的红树林的物种多样性和结构多样性, 从而提高林分的稳定性和健康度, 作用是十分明显的。从群落演替过程上来看, 红树林在发展到一定程度时, 会发生自疏效应, 主要表现为植株分枝干枯频繁发生为特征, 其原因主要是种内个体对资源的竞争[24–25]。因此, 除了要控制好种群密度外, 还要增加处于不同生态位的物种, 以充分利用空间资源, 减少生态位重叠。另外, 研究表明, 红树林的害虫种类与树种有关, 不少害虫有其主要的寄主[10]。因此, 通过增加红树林物种种类群落混交度, 增加群落的物种多样性, 以提高群落的稳定性, 从而增强对病虫害的抵抗能力, 预防针对单一树种的大规模病虫害。海桑和无瓣海桑对提高红树林的结构多样性的作用是明显的, 但也会挤压其他种群的生长空间, 对林分的物种多样性可能会存在消极影响。目前, 海桑及无瓣海桑给红树林带来的生态影响尚存在争议。ZAN等[26]从种群生物生态学特性、生态位理论、群落更新理论等方面分析后, 认为海桑和无瓣海桑在深圳湾造成生态入侵的可能性不大。廖宝文等[27]对海南东寨港红树林的无瓣海桑群落进行研究后, 认为无瓣海桑目前不会造成生态入侵, 但它对今后的生态影响仍需加强监测。有研究表明, 无瓣海桑能够对与其混交的桐花树、白骨壤种群的发展产生不利影响[28], 但能提高秋茄的种群密度[28]。与海桑和无瓣海桑相比, 本土物种海漆、白骨壤与木榄既能增加林分的物种多样性和结构多样性, 又不会造成生态入侵, 是改造以秋茄为建群种或优势种的深圳红市红树林或营造红树林人工林的优良树种。东涌的海漆林是深圳地区乃至华南地区难得一见的天然海漆林。每年夏天, 海漆叶子在会逐渐变黄最终变红, 东涌红树林便形成华南地区难得一见的彩叶红树林景观, 为华南地区红树林林分改造、质量提升或营建兼具生态价值和景观价值的红树林人工林提供了优良的模板。 [1] 段舜山, 徐景亮. 红树林湿地在海岸生态系统维护中的功能[J]. 生态科学, 2004, 23(4): 351–355. [2] 王文卿, 王瑁. 中国红树林[M]. 北京:科学出版社, 2007. [3] 缪绅裕, 廖文波, 苏志尧. 广东红树林研究概况及广东红树林的主要特征[C]∥黄玉山, 谭凤仪. 广东红树林研究论文选集. 广州: 华南理工大学出版社, 1997: 73–84. [4] 陈保瑜, 宋悦, 昝启杰, 等. 深圳湾近30年主要景观类型之演变[J]. 中山大学学报(自然科学版), 2012, 51(5): 86–92. [5] 张倬纶, 侯霄霖, 梁文钊, 等. 深圳现存红树林群落的生境及保护对策[J]. 湿地科学与管理, 2012(4): 49–52. [6] 王志明, 刘晶. 一座城市容不下一片红树林?[N]. 中国环境报, 2012–11–30(008). [7] 姜刘志, 杨道运, 梅立永, 等. 深圳市红树植物群落碳储量的遥感估算研究[J]. 湿地科学, 2018, 16(5): 618–625. [8] 陈桂珠, 彭绍坚, 王震宇, 等. 深圳市西部地区滩涂湿地生态的改造与恢复示范研究[C]//周光召. 加入WTO和中国科技与可持续发展–挑战与机遇、责任和对策(上册). 北京: 中国科学技术出版社, 2002: 497. [9] 沈凌云, 宁天竹, 吴小明, 等. 深圳湾凤塘河口红树林修复工程[J]. 价值工程, 2010(14): 55–57. [10] 昝启杰, 谭凤仪, 李喻春. 滨海湿地生态系统修复技术研究–以深圳湾为例[M]. 北京: 海洋出版社, 2013. [11] 沈小雪, 姜立得, 陶佳伟, 等. 退化红树林的生态修复实验研究[J]. 北京大学学报(自然科学版), 2019 (4), doi: 10.13209/j.0479–8023.2019.046. [12] LY/T 2898—2017, 湿地生态系统定位观测技术规范[S]. 北京: 中国标准出版社, 2008. [13] 韦霄, 柴胜本, 陈宗游, 等. 珍稀濒危植物金花茶保育生物学研究[M]. 南宁: 广西科学技术出版社, 2015. [14] 王伯荪, 余世孝, 彭少麟, 等. 植物群落学试验手册[M]. 广州: 广东高等教育出版社, 1996. [15] LEI X D, LU Y C, Information entropy measures for stand structural diversity: joint entropy[J]. Forestry Studies in China, 2004, 7(2): 12–15. [16] 姜在民, 贺学礼. 植物学[M]. 咸阳: 西北农林科技大学出版社, 2016. [17] 许格希, 史作民, 唐敬超, 等. 物种多度和径级尺度对于评价群落系统发育结构的影响:以尖峰岭热带山地雨林为例. 生物多样性[J], 2016, 24(6): 617–628. [18] 郑景明, 罗菊春. 长白山阔叶红松林结构多样性的初步研究[J]. 生物多样性, 2003(11): 295–302. [19] 周凡, 邝栋明, 简永强, 等. 珠海市淇澳岛红树林群落组成初步研究[J]. 生态科学, 2003, 22(3) : 237–241. [20] 姚少慧, 孙妮, 苗莉, 等. 惠州红树林保护区红树植物群落结构特征[J]. 广东农业科学, 2013(17): 153–157. [21] 刁俊明, 曾宪录. 光强对桐花树根系生长和根系活力的影响. 光强对桐花树根系生长和根系活力的影响[J], 嘉应学院学报, 2013(8): 68–75. [22] 刁俊明, 钟福生, 刘惠娜. 遮光处理对桐花树光合特性的影响[J]. 嘉应学院学报, 2011, 29(5): 71–77. [23] 刘滨尔, 廖宝文. 老鼠簕幼苗在潮汐环境下对不同光强的生理生态响应[J]. 林业科学研究, 2013(2): 102–199. [24] LIAO W B, LAN C Y, ZAN Q J, et al. Growth dynamics and self-thinning of the dominant populations in the mangrove community[J]. Acta Botanica Sinica, 2004, 46(5): 522–532. [25] 孙龙, 国庆喜, 孙慧珍. 生态学基础[M]. 北京: 中国建工业出版社, 2013. [26] ZAN Q J, WANG B S, WANG Y J, et al. Ecological assessment on the introducedandat the mangrove forest of Shenzhen Bay, China[J]. Acta Botanica Sinica, 2003, 45(5): 544–551. [27] 廖宝文, 李玫, 郑松发, 等. 外来种无瓣海桑种内、种间竞争关系研究[J]. 林业科学研究, 2003, 16(4): 418–422. [28] 李玫, 廖宝文, 郑松发, 等. 无瓣海桑的直接引入对次生桐花树群落的扰动[J]. 林业科学与环境, 2004, 20(3): 19–21. [29] 姚历强. 无瓣海桑引入对龙海九龙江口乡土红树林影响的调查[J]. 林业勘察设计, 2010(2): 76–78. Species composition and structural diversity of typical mangrove communities in Shenzhen GUAN Kailang1, ZHANG Xinjian1, TAN Guangwen1, ZENG Feng1, YI Huilin1, LIAO Wenbo2,* 1. Pubang Landscape Architecture Co., LTD, Guangzhou 510600, China 2. School of Life Science, Sun Yat-sen University, Guangzhou 510275, China To investigate the species composition and structural diversity of typical mangrove communities in Shenzhen, eight plots were set up in its main mangrove distribution areas. The results are as follows. (1)was the constructive or dominant natural mangrove species;andwere important constituent species in the natural mangroves;andwere the main constituent species for man-made mangroves;andwere the main species constituting the shrub layer and the less developed mangrove areas. (2) True mangrove plants, such as,,and, etc., enriched the species composition, vertical structure, and diameter structure of Shenzhen mangroves; whileacted as the constructive, dominant or associated species in the region, thus improved the species diversity and structural diversity within the mangroves in becoming a stable stand. The development in natural mangrove quality or cultivated mangroves could improve the ecological stability and landscape value of mangroves by increasing the number of native mangrove plant species, stand structural diversity and mixture level. mangroves; species composition; species diversity; structural diversity; Shenzhen 关开朗, 张信坚, 谭广文, 等. 深圳市红树林典型群落的物种组成及结构多样性研究[J]. 生态科学, 2021, 40(4): 83–91. GUAN Kailang, ZHANG Xinjian, TAN Guangwen, et al. Species composition and structural diversity of typical mangrove communities in Shenzhen[J]. Ecological Science, 2021, 40(4): 83–91. 10.14108/j.cnki.1008-8873.2021.04.010 Q948.15 A 1008-8873(2021)04-083-09 2020-01-19; 2020-03-02 深圳市生态环境局科研项目(SZDL2019333099) 关开朗(1988—), 男, 硕士, 主要从事生物资源调查与评估工作, E-mail: guankailang@foxmail.com 廖文波, 男, 博士, 教授, 主要从事植物区系研究, E-mail: lsslwb@mail.sysu.edu.cn
2.2 群落结构特征

2.3 群落多样性

3 讨论
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
园艺与种苗(2021年3期)2021-05-26
延河·绿色文学(2020年6期)2020-09-10
中学生百科·小文艺(2020年6期)2020-01-13
滇池(2019年1期)2019-02-14
儿童故事画报(2019年12期)2019-01-07
中成药(2018年11期)2018-11-24
小天使·二年级语数英综合(2018年3期)2018-06-29
红豆(2016年6期)2016-05-14
