
bta-miR-1 通过调控PAX7 参与骨骼肌发育的功能研究
2021-11-09 14:18刘瑞莉刘贤勋柏学进董雅娟
中国畜牧杂志 2021年9期
于 堃,刘瑞莉,刘贤勋,柏学进,董雅娟,3*
(1.青岛农业大学动物科技学院,山东青岛 266109;2.山东省黑牛繁育工程技术研究中心,山东青岛 266109;3.山东布莱凯特黑牛科技股份有限公司,山东淄博 256306)
骨骼肌是畜禽肉类食品的重要组成部分,其正常发育是机体维持一系列重要生命活动的基础,而成肌细胞的增殖与分化是骨骼肌发育的关键环节。近年来,有关microRNAs(miRNAs)的研究成为热点,越来越多的人关注研究miRNAs 调控生长发育和肿瘤发生的功能。miRNAs 是一组内源性非编码的小RNA 分子,由19~22 个核苷酸组成,在病毒、动物和植物等生物中广泛存在[1],miRNAs 可以通过影响mRNA 的表达,进而对细胞增殖、细胞分化、细胞凋亡以及脂类代谢等过程进行调控[2-4]。
已有研究表明,miRNAs 的表达具有组织和发育阶段特异性,大部分miRNAs 只在特定组织和发育阶段表达并发挥作用[5]。目前已发现的肌肉特异性表达的miRNAs 有miR-1、miR-133a、miR-133b、miR-206、miR-208a、miR-208b、miR-486 和miR-499[6]。miR-1在多种生命活动中尤其是心肌和骨骼肌发育中发挥重要作用,能通过抑制PAX3的表达而抑制牛骨骼肌卫星细胞的增殖[7],同时miR-1 可通过调控细胞周期蛋白D1进而调控肌细胞周期进程,最终影响肌细胞增殖[8]。近期研究表明当GDF8基因的3'UTR 内一个点突变作用于miR-1、miR-206,引起肌肉生长抑制素基因表达降低,也会发生肌肉异常发育现象[9-10]。本实验的研究对象是布莱凯特黑牛和鲁西黄牛,布莱凯特黑牛是运用体细胞克隆技术、超数排卵技术、胚胎分割及胚胎冷冻技术以及杂交育种与分子标记辅助育种的方法培育而成的优良肉牛新种质[11]。鲁西黄牛是我国优秀的地方良种,具有明显的“三粉特征”,素有“山东膘牛”的美称,1983 年被纳入《中国牛品种志》。由于现有研究虽然证实了bta-miR-1 在骨骼肌发育过程中的重要调控作用,但是关于bta-miR-1 与靶基因的分子调控机制尚无报道,因此探究bta-miR-1 与靶基因之间的分子调控机制,能够为布莱凯特黑牛育种工作奠定理论基础,为肉牛肉质性状的分子选育提供参考。
本研究基于前期实验室建立的布莱凯特黑牛和鲁西黄牛背最长肌的转录组高通量测序数据库,通过KEGG通路和GO 富集分析,筛选出与肉牛骨骼肌发育相关的差异表达bta-miR-1。以bta-miR-1 为研究目标,利用生物信息学方法预测bta-miR-1 的靶基因,分析其保守性,再以C2C12 细胞作为细胞模型,转染blank、NC mimic、bta-miR-1 mimic、NC inhibitor 和bta-miR-1 inhibitor 后,以2%马血清诱导分化,检测靶基因及相关增殖分化标志性基因的表达量变化,探究bta-miR-1通过调控靶基因参与肉牛骨骼肌发育过程的分子机制。
1 材料与方法
1.1 主要材料与试剂 实验动物来自青岛农业大学科研成果转化基地、教学实习基地——山东布莱凯特黑牛科技股份有限公司。选取12 月龄布莱凯特黑牛和鲁西黄牛,采集背最长肌组织置于液氮保存。C2C12 细胞系、bta-miR-1 mimic/inhibitor,购自普健生物(武汉)科技有限公司;miRNA 反转录试剂盒,购自TakaRa Bio USA;iTaqUniwersal SYBR Green Supermix 5 mL(5×1 mL Vials),购自青岛鑫宇恒一科技有限公司;Lipofectamine 3000 试剂,购自赛默飞世尔科技(中国)有限公司;CCK-8 检测试剂盒(CK04),购自东仁化学科技(上海)有限公司。
1.2 小RNA 文库的构建测序与质量控制 用Trizol 试剂(Invitrogen)提取总RNA,使用高质量的RNA 样品构建序列文库。采用Illumina Genome Analyzer(Illumina Genome Analyzer,Illumina,San Diego,CA,USA)对文库进行测序。原始测序数据由Fastp 软件处理,删除包含适配器、ploy-N 和低质量读取数据,获得干净原始读取数据。所有后续分析均基于高质量的数据。通过比对分析软件Bowtie1.1.2 将sRNA 测序的干净读数定位到参考基因组进行比对。
1.3 bta-miR-1 的保守性和靶基因预测 为了探索bta-miR-1的生物学功能,从miRBase(http://www.mirbase.org/)检索牛(Bos taurus,Bta)、人(Homo sapiens,Hsa)、黑猩猩(Pan troglodytes,Ptr)、大鼠(Rattus norvegicus,Rno)、小鼠(Mus musculus,Mmu)、山羊(Capra hircus goat,Chi)、猕猴(Macaca mulatta,Mml)的miR-1 成熟序列,分析其保守性。使用TargetScan(http://www.targetscan.org/vert_72/)软件对bta-miR-1 进行靶基因预测[12]。
1.4 成肌细胞转染及诱导分化 正常培养的C2C12 细胞,吸弃原培养液,PBS 洗后加入胰酶消化1~3 min 后终止消化,剩余细胞悬液用培养基重悬,4 000 个/孔(96孔板)进行铺板(200 μL),3 个复孔,5% CO2、37℃过夜培养。当细胞密度达到90%时,用Lipofectamine 3000 试剂转染浓度为20 μmol/L 的blank、NC mimic、bta-miR-1mimic、NC inhibitor 和bta-miR-1 inhibitor,转染6 h 后,换GM 培养基,12 h 后用2%马血清诱导分化,分别再培养24 h(1 d)、48 h(2 d)、120 h(5 d)和144 h(6 d)观察细胞生长状态,48 h 后提取C2C12成肌细胞总RNA 及总蛋白。
1.5 细胞增殖率检测 转染后24 h、48 h 和72 h 分别从培养箱中取出一个96 孔板,使用细胞计数试剂盒-8(Cell Counting Kit-8,CCK-8)向每孔中加入20 μL CCK8 溶液,培养4 h 后,检测450 nm 处的OD 值[13]。
1.6 实时荧光定量PCR(qRT-PCR)随机选取4 个miRNAs(bta-miR-1、bta-miR-206、bta-miR-196a、bta-miR-200a)进行qRT-PCR 检测验证。检测成肌细胞增殖分化标志性基因PCNA、CDK2、Myog、Myod1和靶基因PAX7的表达量。使用Trizol 法提取blank、NCmimic、bta-miR-1mimic、NC inhibitor 和bta-miR-1 inhibitor 5 组转染48 h 后细胞的总RNA,以1%琼脂糖凝胶电泳鉴定总RNA 质量。将提取的细胞总RNA 逆转录为cDNA,以cDNA 为模板进行荧光定量PCR。荧光定 量PCR 反应体系为20 μL:RNA-freeWater 7.8 μL,上下游引物(10 μmol/L)各0.6 μL,SYBRGreen mix 10 μL,cDNA(80 ng/ μL)1 μL。PCR 反应程序:94℃预变性10 min;94℃变性30 s,60℃退火30 s,72 ℃延伸40 s,40 个循环[14-15]。以GAPDH作为内参基因,对PAX7、PCNA、CDK2、Myog、Myod1基因进行标准化,以U6 作为内参,对bta-miR-1、bta-miR-206、bta-miR-196a、bta-miR-200a 进行标准化,采用2-ΔΔCT值法计算。引物信息见表1。

表1 实时荧光定量PCR 引物
1.7 数据分析 采用SPSS18.0 软件分析数据,结果以“平均值±标准差”表示,测定的数据进行单因素方差分析和显著性检验,P<0.05 表示差异显著,P<0.01 表示差异极显著。
2 结果
2.1 miRNA 数据验证 随机选取4 个miRNA(bta-miR-1、bta-miR-206、bta-miR-196a、bta-miR-200a),通 过qRTPCR 验证测序结果的准确性,结果如图1 所示,通过比较qRT-PCR 与高通量测序的结果(表2)发现,这4个miRNA 在鲁西黄牛和布莱凯特黑牛中表达的变化趋势基本一致,证明高通量测序结果准确可靠。
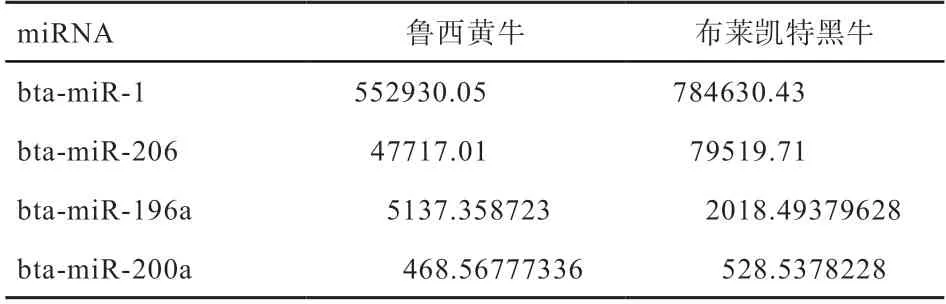
表2 用于qRT-PCR 验证的miRNA 测序表达量数据

图1 miRNA 的qRT-PCR 验证分析
2.2 背最长肌组织中miRNA 的筛选 使用Illumina 基因组分析仪系统测序后,构建sRNA 文库。原始数据过滤后,通过软件Bowtie1.1.2 比对到参考基因组上,共获得26 061 624 个干净读数。与miRBase 数据库的牛物种的参考序列进行比较,匹配到已知miRNA上的Total 和Unique Clean Reads 的总数量分别为29 849 254 和37 946。根据差异miRNA 筛选标准:|log2(Fold_change)| ≥1 且Q ≤0.05,获得差异表达的miRNA 71 个,其中上调39 个,下调32 个(图2a)。根据与肉牛骨骼肌发育相关的功能数据分析结果,筛选出表达量(Transcripts Per Million,TPM)前十位的miRNA 分 别 为bta-miR-1、bta-miR-133a、bta-miR-206、bta-miR-26a、bta-miR-27b、bta-miR-143、bta-miR-378、bta-miR-10b、bta-let-7g、bta-miR-499(图2b)。

图2 转录组文库中差异表达miRNA
2.3 bta-miR-1 的保守性分析 使用miRBase 数据库检索出包括牛在内的7 个物种的miR-1 的成熟序列,结果如表3 所示,可见miR-1(UGGAAUGUAAAGAAGUA UGUAU)在各个物种中高度保守。

表3 不同物种miR-1 成熟序列
2.4 bta-miR-1 靶基因预测 通过KEGG 通路和GO富集分析[16],筛选出与肉牛骨骼肌发育相关的btamiR-1,使用TargetScan 7.1 软件进行靶基因预测,预测到bta-miR-1 的靶基因633 个,其中排名前10 位的靶基因如表4 所示。通过富集分析,筛选到与骨骼肌发育相关的靶基因10 个,分别为PAX7、RORA、QKI、SPEG、PAX5、CITED2、HIVEP3、SERP1、RNF165、GJA1。结合相关文献中已报道的相关功能性靶基因研究分析,选取PAX7作为bta-miR-1 的靶基因。使用TargetScan7.1 软件预测到bta-miR-1 与靶基因PAX7的靶标位区4 个,详细结合靶标位点如表5 所示。

表4 bta-miR-1 的靶基因预测

表5 bta-miR-1 与靶基因PAX7 预测靶标位区
2.5 bta-miR-1 促进肌管形成 如图3 所示,诱导分化1 d时,各组之间无明显差异(P>0.05);2 d 时,与空白组相比,bta-miR-1mimic 组出现大量细胞融合,形成多核细胞,bta-miR-1 inhibitor 组细胞融合数量较少,大部分依然是单核细胞;5 d 时,与空白组相比,btamiR-1mimic 组已有大量肌管形成,bta-miR-1 inhibitor组开始出现少量肌管;6 d 时,与空白组相比,bta-miR-1mimic 组肌管直径增大,数量无明显差异,btamiR-1 inhibitor 组肌管的直径较短,数量无明显差异。
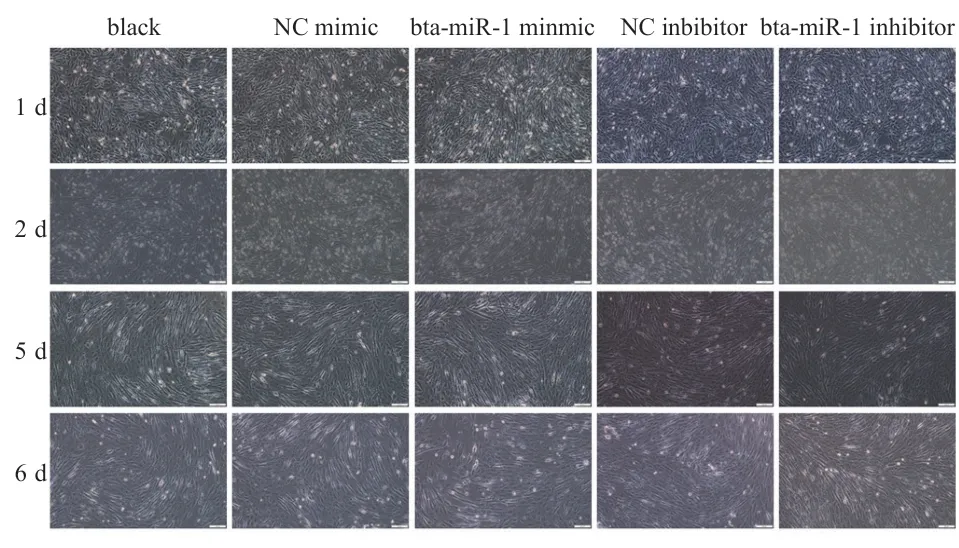
图3 1 d、2 d、5 d、6 d 成肌细胞的诱导分化过程
2.6 bta-miR-1 降低成肌细胞的增殖率 转染后,分别在24 h、48 h 和72 h 进行CCK-8 细胞增殖率检测,结果如图4 所示。由图4 可知,24 h 时,各处理组之间无显著性差异;48 h 时,与空白组相比,bta-miR-1mimic组显著降低成肌细胞的增殖率,bta-miR-1 inhibitor组无显著性差异;72 h 时,与空白组相比,bta-miR-1mimic 组显著降低成肌细胞的增殖率,bta-miR-1 inhibitor 组显著提高成肌细胞的增殖率。
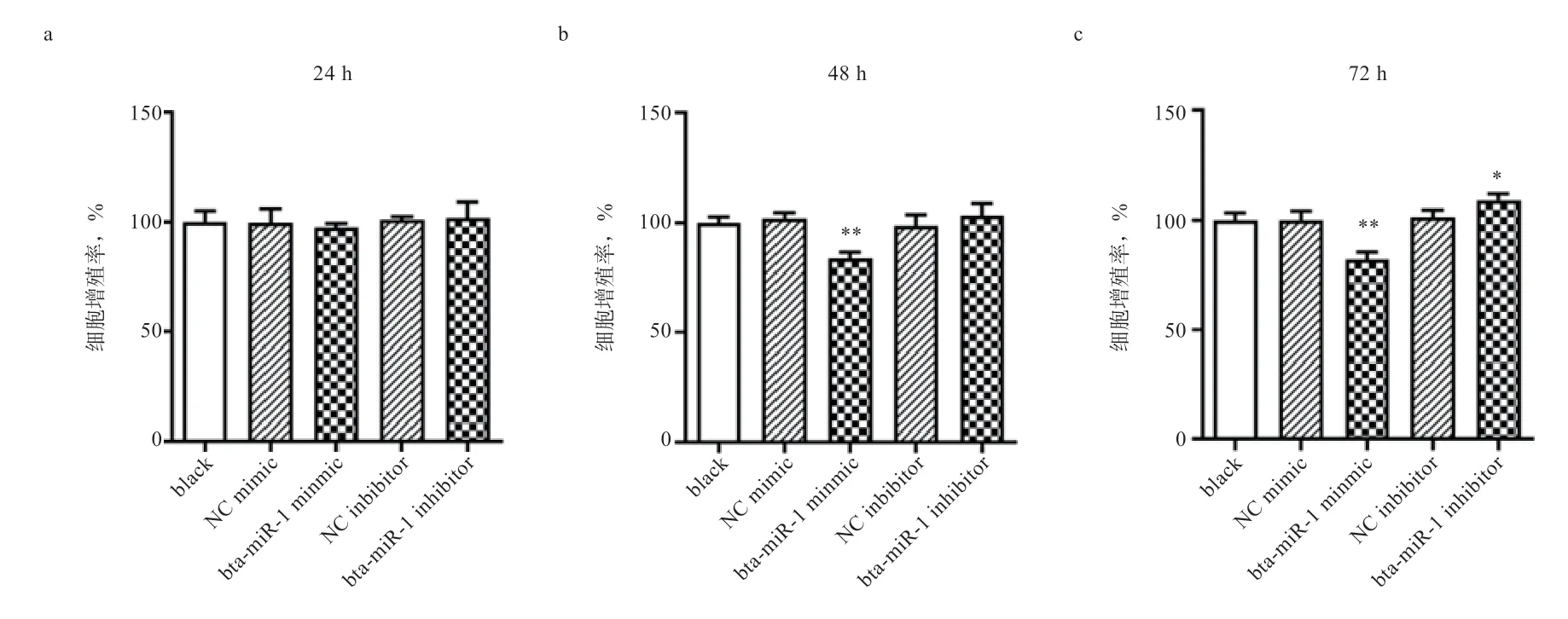
图4 CCK-8 检测在24 h、48 h、72 h 细胞增殖率
2.7 bta-miR-1 对靶基因和增殖分化相关基因的影响转染后,48 h 提取细胞RNA,使用qRT-PCR 检测PAX7、PCNA、CDK2、Myog、Myod1的表达量,结果如图5a 显示,可见bta-miR-1 mimic 组极显著抑制靶基因PAX7的表达,bta-miR-1 inhibitor 组极显著促进PAX7的表达。在细胞增殖方面,如图5b 所示,btamiR-1mimic 组极显著抑制PCNA的表达,对CDK2的表达没有显著影响,bta-miR-1 inhibitor 组极显著促进PCNA和CDK2的表达;在细胞分化方面,如图5c 所示,bta-miR-1mimic 组极显著促进Myog和Myod1的表达,bta-miR-1 inhibitor 组显著抑制Myog和Myod1的表达(P<0.05)。
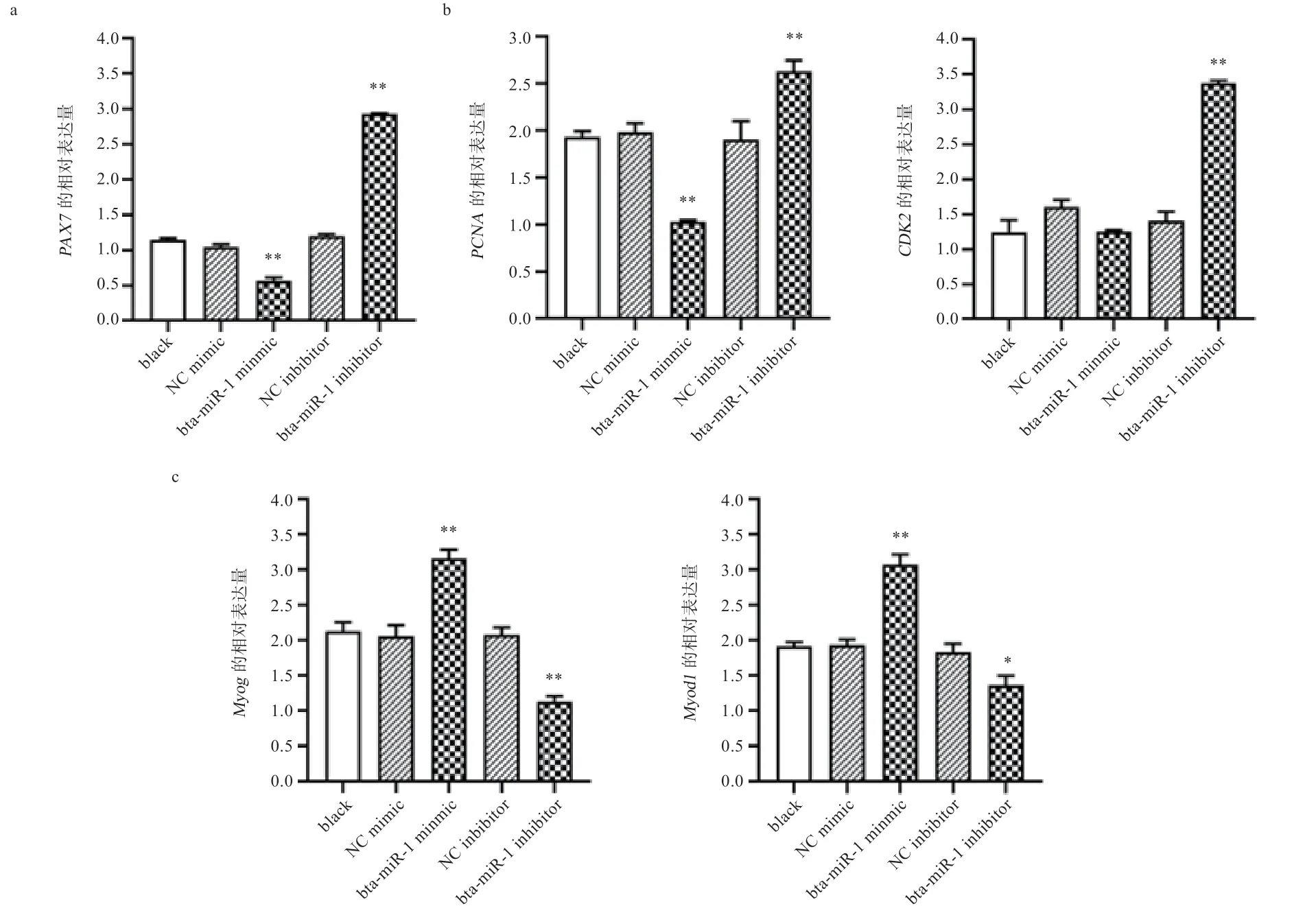
图5 PAX7、PCNA、CDK2、Myog、Myod1 基因的相对表达量
3 讨论
已有研究报道骨骼肌形成的过程是一个受多种调节因子和信号通路共同调控的复杂有序的生物学过程,其中miRNAs 广泛参与调控骨骼肌的发育过程,当前关于miRNAs 在骨骼肌发育中的潜在机制和作用的研究备受关注。本研究使用Illumina Solexa 测序技术,建立布莱凯特黑牛和鲁西黄牛骨骼肌测序RNA 文库,采用KEGG 通路和GO 富集分析筛选出与肉牛骨骼肌发育相关的差异miRNAs(bta-miR-1、bta-miR-133a、btamiR-206、bta-miR-26a、bta-miR-27b、bta-miR-143、bta-miR-378、bta-miR-10b、bta-let-7g、bta-miR-499),选取TPM 值最高的bta-miR-1 作为研究对象。miR-133a 可通过靶向SRF,降低其蛋白水平,进而抑制成肌细胞分化,促进成肌细胞增殖[17];bta-miR-143 可通过靶向调控靶基因IGFBP5 的表达进而调控牛骨骼肌卫星细胞增殖与分化过程[18];bta-miR-206、bta-miR-26a、bta-miR-27b、bta-miR-143 等与肌肉发育同样密切相关,与Qin 等[19]研究结果相一致。通过保守性分析,比对不同物种miR-1 成熟序列(UGGAAUGUAAA GAAGUAUGUAU)的同源性,发现miR-1 在各个物种中高度保守,表明miR-1 在生物进化过程中可能具有重要功能。已有研究发现,miR-1 在小鼠、羊等动物肌肉中高度表达,广泛参与骨骼肌的生长发育[20-21]。通过生物信息学软件分析,进行bta-miR-1 靶基因预测,获得靶基因633 个,发现排名前10 位的靶基因(TNKS2、PAX7、SLC44A1、BDNF、NCL、SRSF9、HNRNPU、PDE7A、SNAP25、MON2)中,多与肿瘤疾病相关,例如:TNKS2基因异常激活会导致宫颈癌的发生[22];NCL基因可促进细胞增殖分化,引发胃癌[23];SRSF9基因多与肺癌细胞、胶质瘤细胞增殖和迁移有关[24]。基于本实验对不同品种间肉牛骨骼肌生长发育机制的研究,对bta-miR-1 靶基因进行富集分析,获得与骨骼肌生长发育相关的靶基因10 个(PAX7、RORA、QKI、SPEG、PAX5、CITED2、HIVEP3、SERP1、RNF165、GJA1),根据相关文献报道,QKI 转录水平下调促使卫星细胞进入细胞周期,完成肌肉损伤修复,且研究多集中在神经系统上[25];GJA1与心肌和肺脏等多种器官的细胞功能发挥有一定的联系,且主要影响肿瘤的发生等过程[26];靶基因PAX7主要参与心肌与骨骼肌的生长发育,在成年期和肌肉再生过程中起主要作用,对卫星细胞的生物发生和存活至关重要[27-28]。同时也有研究表明,PAX7可通过与肌肉调节因子家族成员之间的交叉抑制,维持着卫星细胞的自我更新,从而决定了骨骼肌维持和修复所必需的增殖、分化和自我更新[29-32]。以上研究表明PAX7与肌肉生长发育调节机制的功能相关性最强,且与bta-miR-1 结合的稳定性最强。本研究进一步预测到bta-miR-1 的种子序列(UGUAAGG)与靶基因PAX7的靶标位区4 个,因此选取PAX7作为靶基因,推测bta-miR-1 可能通过靶基因PAX7参与调控骨骼肌的发育,具体的靶标位区有待于进一步探究。
为进一步探究bta-miR-1 参与骨骼肌发育的分子调控机制,以C2C12 细胞作为细胞模型,转染bta-miR-1后,诱导分化发现,与空白组相比,2 d 时,各处理组之间开始有明显差异,均进入细胞融合过程;5 d 时,bta-miR-1mimic 组的细胞有大量肌管形成;6 d 时,bta-miR-1mimic 组的肌管直径明显增粗,而转染btamiR-1 inhibitor 组肌管直径相对较短。结果发现,在转染后,各个处理组的细胞均进入肌管分化过程,虽然bta-miR-1 inhibitor 组的成肌细胞分化较慢,但依然能形成肌管,表明bta-miR-1 inhibitor 可以抑制成肌细胞分化,但不能完全抑制其分化,只能减缓其分化速度。同时发现,在0~5 d 时,肌管分化过程中,bta-miR-1作用并不明显,但在5~6 d 时,肌管的分化速度明显加快,且bta-miR-1mimic 组和bta-miR-1 inhibitor 组肌管数量和直径大小对比最为明显,表明bta-miR-1 主要在成肌细胞分化后期发挥作用。以上实验结果证实bta-miR-1 促进肌管分化,与毛旭东[33]、秦瑞峰[34]、陈永乐等[35]人探究的关于成肌细胞分化凋亡的过程相一致;此外,关于bta-miR-1 在肌管分化后期发挥主要作用的调节机制有待于进一步研究。同时采用CCK-8 检测细胞增殖率,发现转染后72 h 内,bta-miR-1 mimic组抑制细胞增殖的速率在24~48 h 最大,而bta-miR-1 inhibitor 组促进细胞增殖的速率在48~72 h 较大,结果说明bta-miR-1 抑制成肌细胞增殖,且主要在细胞增殖的前期发挥作用。
为进一步探究bta-miR-1 通过靶基因PAX7参与调控骨骼肌发育的具体调节机制,进行细胞功能实验验证。通过qRT-PCR 检测靶基因PAX7表达量,发现bta-miR-1 可抑制靶基因PAX7表达。Margaret 等[36]人发现p38α/ YYI/ PRC2 复合物会导致PAX7的转录受到抑制,进而启动细胞成肌分化,因此推测bta-miR-1促进成肌细胞分化。在成肌细胞的增殖分化过程中已报道多个标记基因,如增殖细胞核抗原(PCNA)、周期蛋白依赖性激酶(CDK2)、生肌调控因子家族(MRFs)和肌细胞增强因子家族(MEFs)等[37]。目前关于成肌细胞增殖和分化的研究中,多采用Myod1、Myog和PCNA、CDK2来反映分化和增殖程度[38]。因此,本实验选取PCNA、CDK2、Myod1和Myog作为细胞增殖分化标志基因,实时荧光定量检测其表达量。与空白组相比,过表达bta-miR-1 显著促进分化标志基因Myod1和Myog表达,抑制增殖标志基因PCNA和CDK2表达,而抑制bta-miR-1 显著抑制分化标志基因Myod1和Myog表达,促进增殖标志基因PCNA和CDK2表达。已有研究表明,在细胞增殖过程中,增殖标志基因PCNA和CDK2广泛存在于增殖的细胞中,促进细胞增殖[39]。在成肌细胞分化过程中,分化标志基因Myod1和Myog同属于MRFs 家族,Myod1能促进肌卫星细胞向成肌细胞转化以及促进成肌细胞融合形成肌管;Myog促使单核成肌细胞向多核的肌纤维转化[40]。因此,本实验结果表明,bta-miR-1 促进成肌细胞分化,抑制成肌细胞增殖,结果与Chen 等[41]人的研究相一致。
研究进一步表明,bta-miR-1 对成肌细胞的分化过程的调控不仅仅通过靶基因PAX7,也调控其他调节因子来参与骨骼肌的发育过程,bta-miR-1 既能作用于多个靶基因,也可以作用在同一个靶基因的不同位点。在骨骼肌发育过程中,bta-miR-1 与靶基因之间的分子调控机制十分复杂,有待于进一步探索[42]。本研究表明,在成肌细胞的增殖分化过程中,bta-miR-1 促进成肌细胞分化,抑制成肌细胞增殖,进而参与骨骼肌生长发育调控过程。
4 结论
本研究基于转录组数据库筛选出与肉牛骨骼肌生长发育相关的bta-miR-1,并证实bta-miR-1 通过抑制靶基因PAX7的表达,促进肌细胞的分化调节,参与骨骼肌的生长发育,为肉牛肉质性状的分子选育以及功能基因组学的研究提供参考。
猜你喜欢
中国人兽共患病学报(2022年9期)2022-10-19
体育科技文献通报(2022年3期)2022-05-23
清华金融评论(2022年4期)2022-04-13
医学综述(2021年16期)2021-12-01
科学导报(2021年29期)2021-06-03
国际放射医学核医学杂志(2021年10期)2021-02-28
房地产导刊(2020年7期)2020-08-24
中国外汇(2019年14期)2019-10-14
科海故事博览·下旬刊(2019年6期)2019-04-16
运动(2018年14期)2018-07-16
