
河水添加对栉孔扇贝生理和免疫指标的影响
2021-11-19 07:35李长青车建锋崔玉龙邹战明黄二帅卢龙飞常丽荣
江苏农业科学 2021年20期
李长青 车建锋 崔玉龙 邹战明 黄二帅 卢龙飞 常丽荣

摘要:为探究河水注入养殖海域对栉孔扇贝的影响,挑选山东省荣成市爱莲湾海域的养殖扇贝为试验对象,研究其在毗邻河水(0、10%、20%、30%、50%)添加后存活率、耗氧率、排氨率、SOD活性及重金属含量的变化。在试验过程中,0、10%、20%河水添加组存活率均≥95%,30%河水添加组存活率为91%,50%河水添加组栉孔扇贝于120 h时全部死亡;在1~48 h间,10%、20%、30%河水添加组栉孔扇贝的耗氧率、排氨率和闭壳肌SOD活性均呈现先下降后上升的趋势,并在120 h后逐渐趋于稳定。50%河水添加组栉孔扇贝的耗氧率、排氨率和闭壳肌SOD活性持续最低(P<0.05)。在试验结束(336 h)时,10%河水添加组耗氧率显著高于其他试验组(P<0.05),排氨率和闭壳肌SOD活性略高于0、20%河水添加组(P>0.05)。30%河水添加组耗氧率、排氨率和闭壳肌SOD活性显著低于其他试验组(P<0.05);试验结束时,各试验组栉孔扇贝体内的Pb、Cr、Hg、As含量均有提高,显著高于无(0)河水添加组。
关键词:栉孔扇贝;河水;耗氧率;排氨率;重金属;免疫指标
中图分类号:S944.4+3;S968.31 文献标志码: A
文章编号:1002-1302(2021)20-0172-05
收稿日期:2021-03-18
基金项目:国家重点研发计划(编号:2018YFD0900703);山东省泰山产业领军人才工程高效生态农业创新类项目(编号:LJNY201816)。
作者简介:李长青(1967—),男,山东威海人,从事水产养殖与增殖研究。E-mail:592378772@qq.com
通信作者:常丽荣,硕士研究生,从事水产养殖和水产品加工研究。E-mail:xs-chengguo@163.com。
栉孔扇贝(Chlamys farreri)主要产于山东半岛及辽宁大连周边海域[1],是我国北方沿海贝类养殖的重要经济种类,其肉质鲜美,深受人们的喜爱。然而,近年来环境因子的变化,致使养殖贝类品质和产量下降,制约了贝类产业的发展[2]。已有学者探究了温度、盐度、pH值、溶解氧含量、营养盐含量等对贝类生理代谢和免疫能力的影响[3-8],其中盐度是影响贝类生长存活的重要环境因子[9-10]。盐度的变化不仅会对贝类的呼吸和摄食活动产生影响,还会影响体内免疫酶的活性,进而影响贝类的生长及存活[11-13]。贝类作为变渗透压动物,对海水盐度的适应具有一定范围,能根据盐度调节自身渗透压,从而保持正常的生命活动;但超出适应范围,贝类不能将其双壳完全关闭,并且其自身血淋巴渗透压调节具有一定的延迟性,不能维持正常的生理代谢[14]。胡益鸣等研究发现,盐度对岩牡蛎的存活和免疫指标影响具有显著的差异,适宜岩牡蛎存活的盐度为2.0%~3.6%[15]。聂鸿涛等报道了在盐度2.0%~4.0%之间,耗氧率随着盐度的升高先减少后增加,排氨率随着盐度的升高先增加后减少[16]。因此,研究盐度胁迫这一环境因子对栉孔扇贝生理和免疫指标的影响,有助于人们了解栉孔扇贝的逆境适应机制。
扇贝养殖主要集中在浅海,夏季持续的高温和降雨导致地表径流增加,会引起近岸海域盐度的大幅度变化[17]。地表径流汇入河槽,流入大海,在改变盐度的同时,水体中可能混有营养盐和重金属等成分[18-20],很可能对近岸扇贝养殖产生影响。然而,河水對贝类影响的相关探究并未见报道。因此,本试验探究海水中添加河水对栉孔扇贝生理和免疫指标的影响,以期为近岸栉孔扇贝健康养殖提供数据参考。
1 材料与方法
1.1 试验材料
试验所用栉孔扇贝取自威海长青海洋科技股份有限公司的爱莲湾近岸养殖海域,选取壳长、个体质量基本一致的栉孔扇贝作为试验个体,扇贝初始壳长平均为(46.79±4.35) mm,个体质量为(11.25±3.42) g;河水取自该海域的车道河中上游,以保证试验结果与生长养殖活动相符。
1.2 试验方法
1.2.1 前期处理
将栉孔扇贝保持潮湿状态下运回养殖车间进行暂养,暂养水温12~14 ℃;暂养期间每天07 ∶30和17 ∶30定量投喂三角褐指藻,连续充气,流水养殖,暂养7 d后挑选活力较强的健康扇贝进行试验。
1.2.2 试验设计
设置无(0)、10%、20%、30%、50%河水添加量共计5个试验组,海水盐度为3.3%。添加后的试验组组盐度分别为3.30%、2.97%、2.64%、2.31%、1.65%。每组设3个平行,每个平行放入60只栉孔扇贝,养殖水温12~14 ℃。试验期间每天07 ∶30和17 ∶30定量投喂三角褐指藻,连续充气,每天换水(换水量为100%)。
暂养期间,随机选取9只栉孔扇贝测定耗氧率和排氨率,取3只栉孔扇贝测定超氧化物歧化酶(SOD)活性,每个指标设3个平行。试验开始后,在试验1、6、24、48、120、192、264、336 h时,每个平行分别随机选取3只栉孔扇贝测其耗氧率和排氨率,取3只栉孔扇贝测其SOD活性,每个指标设3个平行;在试验结束时,取剩余栉孔扇贝测定其体内重金属含量。
1.3 指标测定
1.3.1 营养盐及水文参数
铵态氮(NH+4-N)浓度测定采用次溴酸钠氧化法[21-23],铵态氮(NO-3-N)浓度测定采用重氮-偶氮法[21-23],硝态氮(NO-2-N)浓度测定采用锌-镉还原法,pH值采用pH计测定,溶解氧含量测定采用Winker碘量法。
1.3.2 存活率
试验过程中记录各试验组栉孔扇贝死亡数,试验结束后计算存活率。
1.3.3 耗氧率和排氨率
采用静水密封法,将完好的自封袋(确保严格密封)放入水槽中,灌入一定体积的海水,放入栉孔扇贝并排出空气,持续2 h。溶解氧(DO)含量的测定采用Winker碘量法,NH+4-N浓度测定采用次溴酸钠氧化法[21-23]。
根据试验前后溶解氧含量的变化计算单位湿质量耗氧率:
RO=[C0(DO)-Ct(DO)]V/mt;
式中:C0(DO)和Ct(DO)分别为试验开始和结束时试验水中DO含量,mg/L;V为试验用水的体积,L;m为试验贝软组织湿质量,g;t为试验持续时间,h。
根据试验前后氨氮浓度的变化计算单位湿质量排氨率:
RN=[Ct(N)-C0(N)]V/mt;
式中:C0(N)和Ct(N)分别为试验开始和结束时试验水中氨氮浓度,μmol/L;V为试验用水的体积,L;m为试验贝软组织湿质量,g;t为试验持续时间,h。
1.3.4 超氧化物歧化酶(SOD)活性
冰浴条件下取栉孔扇贝闭壳肌放入冻存管,-80 ℃保存备用。SOD活性的测定采用南京建成生物公司的试剂盒,按照说明书步骤进行。
1.3.5 重金属含量
采用原子吸收分光光度法[24-25],测定不同试验组扇贝体内Cr、Pb、Cd、As、Hg等重金属含量。
1.4 数据分析
试验数据采用SPSS 19.0軟件进行分析比较,采用“平均值±标准差”表示,进行单因素方差分析(One-Way ANOVA)和Duncans多重比较分析,P<0.05表示差异具统计学意义。
2 结果与分析
2.1 河水、海水水质的基本情况
由表1可知试验用海水和河水水质的基本情况,海水符合国家规定的二类水质标准,河水符合国家规定的三类水质标准。
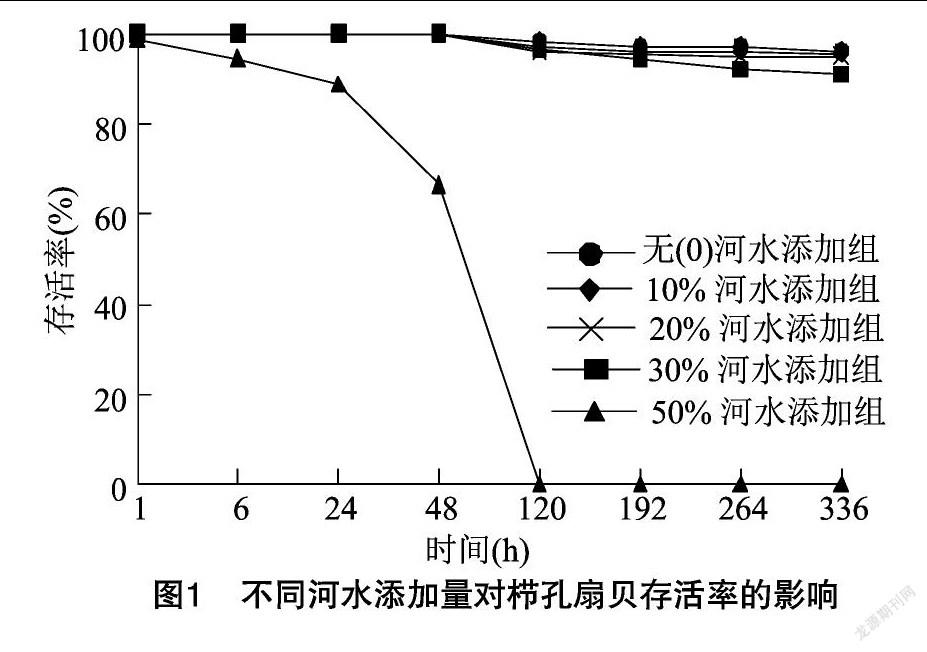
2.2 不同河水添加量对栉孔扇贝存活率的影响
由图1可知,0、10%、20%河水添加试验组存活率较高,均在95%以上,显著高于其他试验组(P<0.05);其次是30%河水添加组,存活率为91%;50%河水添加组存活率显著低于其他试验组(P<0.05),并随着时间的延长逐渐下降,6 h为94%,24 h 为89%,48 h为67%,120 h时全部死亡。
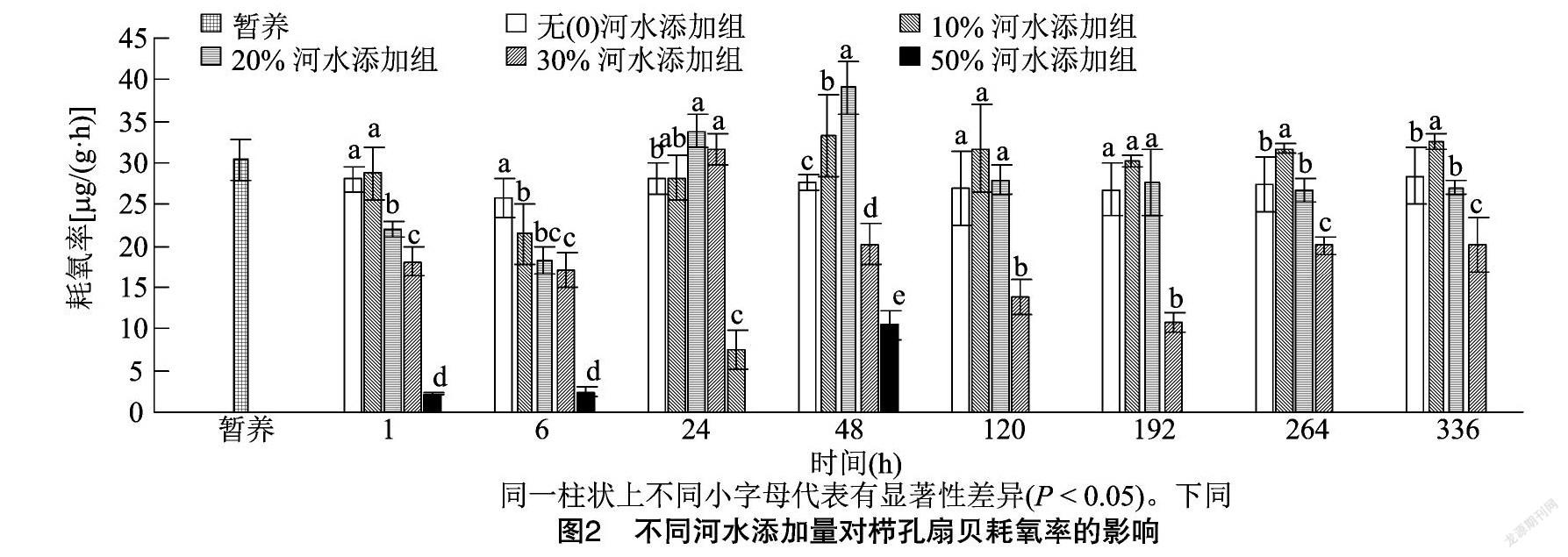
2.3 不同河水添加量对栉孔扇贝耗氧率和排氨率的影响
由图2可知,试验过程中栉孔扇贝的耗氧率在2.21~39.09 μg/(g·h)之间,0、10%、20%河水添加试验组的耗氧率在1、48、120、192、264、336 h时均显著高于30%、50%河水添加试验组(P<0.05);在6 h时,各试验组耗氧率均出现不同程度降低,无河水添加试验组降低的原因可能与流水养殖突变为静水养殖有关;在24、48 h时,各试验组耗氧率呈现上升趋势,处于不稳定阶段,经分析,可能是因为盐度改变导致栉孔扇贝出现应激反应,进行自身调节;120 h后逐渐趋于稳定;在264、336 h 时,10%河水添加试验组耗氧率最高,显著高于0、20%河水添加试验组(P<0.05),30%河水添加试验组最低,显著低于其他试验组(P<0.05);在1~48 h,50%河水添加试验组耗氧率持续最低,显著低于其他试验组(P<0.05)。
由图3可知,不同河水添加量对栉孔扇贝排氨率的影响与耗氧率呈现类似的趋势,栉孔扇贝的排氨率在0.05~0.77 μg/(g·h)之间。在试验 1~48 h 之间,无河水添加试验组排氨率与暂养时差异不显著(P>0.05),其他试验组均呈现先下降后上升的趋势;120 h后趋于稳定,在264、336 h时,10%河水添加试验组排氨率最高,略高于0、20%河水添加试验组(P>0.05),30%河水添加试验组最低,显著低于其他试验组(P<0.05);在1~48 h,50%河水添加试验组排氨率持续最低,显著低于其他试验组(P<0.05)。
2.4 不同河水添加量对栉孔扇贝超氧化物歧化酶(SOD)活性的影响
由图4可知不同河水添加量对栉孔扇贝闭壳肌SOD活性的影响,栉孔扇贝闭壳肌SOD活性在23.73~45.42 U/mg之间,整个试验过程中,无河水添加试验组SOD活性与暂养时无显著差异(P>0.05)。在试验1 h时,0、10%、20%、30%河水添加试验组SOD活性与暂养时处于同一水平,无显著差异(P>0.05),50%河水添加试验组SOD活性最低,显著低于其他试验组(P<0.05);在试验1~48 h 之间, 10%、 20%、 30%、50%河水添加试验组出现先下降后升高的趋势;120 h后趋于稳定,无河水添加试验组SOD活性显著高于10%、20%、30%河水添加试验组(P<0.05)。
2.5 不同河水添加量对栉孔扇贝体内重金属含量的影响
由表2可知,试验结束时,各不同处理组栉孔扇贝体内Pb、Cr、Cd、Hg、As的含量(以湿质量计),栉孔扇贝体内Pb含量范围为0.086~0.124 mg/kg,Cr含量范围为0.059~0.086 mg/kg,Cd含量范围为 0.085~0.363 mg/kg,Hg含量范围为0.034~0.044 mg/kg,As含量范围为0.276~0.391 mg/kg,可见栉孔扇贝体内5种重金属含量均未超标。50%河水添加组于120 h全部死亡,不参与比较。无河水添加组Pb、Cr、Hg、As均显著低于其他试验组(P<0.05),可见河水的添加可能导致栉孔扇贝体内重金属含量的增加。
3 讨论与结论
本研究发现50%河水添加试验组于120 h全部死亡,其他试验组存活率均在90%以上。吴静研究发现,盐度在2.6%~3.2%时,华贵栉孔扇贝的存活率较高,均在85%以上,盐度1.8%时,120 h全部死亡[14],与本研究结果基本一致。说明极端盐度胁迫导致海水渗透压的改变超过了栉孔扇贝自身的调节能力,导致栉孔扇贝代谢紊乱,甚至死亡[24]。
呼吸和排泄是贝类新陈代谢的基本生理活动,也是贝类能量学、贝类养殖容量研究以及贝类对海洋生态系统影响的重要评价内容[18],不仅可以反映贝类的生理活动,还可以反映环境条件对贝类生理活动的影响。在一定盐度范围内,华贵栉孔扇贝、泥螺、泥蚶的耗氧率和排氨率都随着盐度的升高而升高,分别在盐度3.2%、2.8%、2.8%时最高,随后逐渐降低[14,25-26]。本研究的耗氧率和排氨率也呈现先下降后上升再趋于稳定的趋势,稳定后10%河水添加组最高,即盐度2.97%时达到最大值,与上述研究结果一致。有学者认为,出现上述情况的原因是贝类在超出适应的海水盐度范围后,为了适应周围环境渗透压的变化,通过降低耗氧率和排氨率等来维持正常的生理活动[25];在试验1~48 h耗氧率和排氨率均出现较大的波动,尤其在24、48 h时,10%、20%、30%河水添加组耗氧率出现显著升高,可能是由于栉孔扇贝为减轻盐度变化的影响,需要消耗大量的能量调节自身渗透压[13],进而保持正常的生命体征。
超氧化物歧化酶(SOD)是一种抗氧化酶,可以增强机体的免疫能力和吞噬细胞的防御能力[27],保护机体免受活性氧自由基的损伤[28-29],可以反映贝类机体的健康状态[30]。本研究显示,在试验1 h时,其他试验组SOD活性与暂养时处于同一水平,显著高于50%试验组(P<0.05)。50%试验组SOD活性持续最低,表明极端的盐度变化超出了栉孔扇贝的自我调节能力。而10%、20%和30%河水添加组SOD活性在6~336 h之间先下降后上升并趋于稳定,稳定后显著低于无河水添加组(P<0.05)。刘美剑报道了盐度变化导致扇贝血清中SOD活性显著下降后逐渐恢复[8],与本研究这一趋势类似。6~48 h之间是SOD活性上升的阶段,说明机体在通过自身调节提高SOD活性以清除由于盐度胁迫积累的大量活性氧自由基,来提高自身免疫能力。
本研究发现,添加河水后,栉孔扇贝体内Pb、Cr、Hg、As含量显著增加,海洋动物对重金属具有较强的富集性,人类经食物链可将富集在其体内的重金属进一步积累,危害身体健康[31]。工业废水、生活污水等经河流进入海洋,造成海洋环境污染,尤其是重金属污染,对于栉孔扇贝、贻贝、牡蛎等滤食性贝类来说,极易造成体内重金属含量超标,人类食后在体内聚集又很难去除,严重导致结蹄组织受损,肌肉萎缩,甚至癌变死亡[32]。
综上所述,夏季降雨频发、河水大量流入海洋,不仅会对近岸养殖的海产品造成产量和品质下降,严重时可能威胁人类健康,因此工业废水和生活污水要处理达标后排放。本研究对河水添加后栉孔扇贝生理、免疫、重金属等指标的影响进行初步探究,并不能全方位了解其本质影响,还需进一步探究。
参考文献:
[1]李成林,宋爱环,胡 炜,等. 山东省扇贝养殖产业现状分析与发展对策[J]. 海洋科学,2011,35(3):92-98.
[2]王 尧. 盐度变化对岩扇贝相关酶活性及基因表达的影响[D]. 大连:大连海洋大学,2019.
[3]王有基,李丽莎,李琼珍,等. 海洋酸化和全球变暖对贝类生理生态的影响研究进展[J]. 生态学报,2014,34(13):3499-3508.
[4]刘大鹏. 三种生态因子对三角帆蚌生理指标的联合效应研究[D]. 重庆:西南大学,2020.
[5]董莎莎,聂鸿涛,闫喜武. 贝类低温胁迫响应机制研究进展[J]. 大连海洋大学学报,2019,34(3):457-462.
[6]姜娓娓. 扇贝和皱纹盘鲍对温度变化的生理响应研究[D]. 北京:中国科学院大学,2017.
[7]霍恩泽. 两种牡蛎对盐度骤降的生理响应及长牡蛎动态能量收支模型构建[D]. 上海:上海海洋大学,2020.
[8]刘美剑,常亚青,杨芸菲,等. 盐度渐变对虾夷扇贝(Patinopecten yessoensis)免疫指標的影响[J]. 中国农业科技导报,2011,13(3):129-135.
[9]Cain T D. The combined effects of temperature and salinity on embryos and larvae of the clam Rangia cuneata[J]. Marine Biology,1973,21(1):1-6.
[10]Widdows J. The effects of fluctuating and abrupt changes in salinity on the performance of Mytilus edulis[C]//Gray J S, Christiansen M E. Marine biology of polar regions and effects of stress on marine organisms: Proceedings of the 18th European Marine Biology Symposium. Norway,1983:555-566.
[11]Rupp G S,Parsons G J. Effects of salinity and temperature on the survival and byssal attachment of the lions paw scallop Nodipecten nodosus at its southern ditribution limit[J]. J Exp Mar Biol Ecol,2004,309(2):173-198.
[12]Laing I. Effect of salinity on growth and survival of king scallop spat (Pecten maximus)[J]. Aquaculture,2002,205(1/2):171-181.
[13]时少坤. 环境因子对贝类几种免疫因子影响的研究[D]. 上海:上海海洋大学,2013.
[14]吴 静. 温度和盐度对华贵栉孔扇贝(Mimachlamys nobilis Reeve)存活、免疫指标及生理指标的影响[D]. 福州:福建师范大学,2016.
[15]胡益鸣,李 琪,刘士凯,等. 温度和盐度急性胁迫对岩牡蛎存活及免疫指標的影响[J]. 中国水产科学,2020,27(3):286-294.
[16]聂鸿涛,霍忠明,侯晓琳,等. 温度和盐度突变对菲律宾蛤仔斑马蛤耗氧率和排氨率的影响[J]. 水生生物学报,2017,41(1):121-126.
[17]Durack P J,Wijffels S E,Matear R J. Ocean salinities reveal strong global water cycle intensification during 1950 to 2000[J]. Science,2012,336(680):455-458.
[18]劳齐斌,刘国强,申友利,等. 北部湾入海河流营养盐的分布特征及入海通量研究[J]. 海洋学报,2020,42(12):93-100.
[19]夏同法,梁胜跃,徐明钻,等. 江苏灌河入海口附近重金属元素环境地球化学质量分析[J]. 地质学刊,2018,42(4):641-650.
[20]张思洋,于大涛,张 戈. 锦州湾三河入海口重金属分布特征及污染评价[J]. 海洋地质前沿,2020,36(6):20-25.
[21]海洋调查规范 第4部分:海水化学要素调查:GB/T 12763.4—2007[S].
[22]海洋调查规范 第4部分:海水分析[S].
[23]海洋调查规范 第6部分:生物体分析[S].
[24]刘志刚,刘建勇,王 辉,等. 墨西哥湾扇贝稚贝盐度适应性的研究[J]. 湛江海洋大学学报(自然科学),2006,26(6):12-16.
[25]吴文广,张继红,方建光,等. 盐度对不同规格泥螺耗氧率和排氨率的影响[J]. 生态学杂志,2013,32(9):2457-2461.
[26]熊安安,袁 星,汪先进,等. 盐度和光照对泥蚶耗氧率和排氨率的影响[J]. 水生态学杂志,2015,36(5):96-100.
[27]姚翠鸾,王维娜,王安利. 水生动物体内超氧化物歧化酶的研究进展[J]. 海洋科学,2003,27(10):18-21.
[28]Chen J H,Mai K S,Ma H M,et al. Effects of dissolved oxygen on survival and immune responses of scallop (Chlamys farreri Jones et Preston)[J]. Fish & Shellfish Immunology,2007,22(3):272-281.
[29]Ekanayake P M,de Zoysa M,Kang H S,et al. Cloning,characterization and tissue expression of disk abalone (Haliotis discus discus) catalase[J]. Fish & Shellfish Immunology,2008,24(3):267-278.
[30]吕昊泽,刘 健,陈锦辉,等. 盐度对缢蛏超氧化物歧化酶和过氧化氢酶活性的影响[J]. 海洋渔业,2013,35(4):474-478.
[31]姜 杰,丘红梅,张慧敏,等. 广东沿海海域海产品中重金属的含量及评价[J]. 环境与健康杂志,2009,26(9):814-816.
[32]陈红英,陈旭凌,陈笑霞,等. 华贵栉孔扇贝体内重金属含量及评价[J]. 微量元素与健康研究,2015,32(1):33-34,36.
猜你喜欢
淡水渔业(2023年2期)2023-03-29
小学生作文(低年级适用)(2019年12期)2020-01-18
人大建设(2019年5期)2019-10-08
河北渔业(2019年7期)2019-08-27
水产科学(2018年3期)2018-06-04
广东海洋大学学报(2015年3期)2015-12-22
应用海洋学学报(2014年1期)2014-11-22
浙江海洋大学学报(自然科学版)(2014年6期)2014-03-13
