
New fossils of small and medium-sized bovids from the Early Pleistocene Site of Shanshenmiaozui in Nihewan Basin, North China
2022-06-18 09:57TONGHaoWenZHANGBeiCHENXiWANGXiaoMin
古脊椎动物学报(中英文) 2022年2期
TONG Hao-Wen ZHANG Bei CHEN Xi WANG Xiao-Min
(1 Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences Beijing 100044* Corresponding author: tonghaowen@ivpp.ac.cn)
(2 CAS Center for Excellence in Life and Paleoenvironment Beijing 100044)
(3 University of Chinese Academy of Sciences Beijing 100049)
(4 Beijing Museum of Natural History Beijing 100050)
(5 Nanjing Normal University Nanjing 210023)
(6 Institute of Archaeology, Chinese Academy of Social Sciences Beijing 100710)
Abstract Shanshenmiaozui site in Nihewan Basin in North China is a recently discovered Early Pleistocene site which yields rich and diverse mammalian fossils. In the fauna, the small and medium-sized bovid fossils are well represented and can be referred to the following taxa:Spirocerus wongi, Gazella sinensis, Ovis shantungensis and Megalovis piveteaui respectively,among which G. sinensis is the dominate species. S. wongi and G. sinensis are mainly represented by horn-cores and partial skull bones as well as mandibles; in addition, metacarpal and/or metatarsal bones were also recognized for all of the four species. The horn-cores are easy to be identified to the species level, while the dentitions and the postcranial bones underwent a series of examinations and comparisons before getting properly determined and referred to the most approximate taxa. Among the postcranial bones, the metapodials, especially to the metacarpal bones special attentions were paid, which are crucial not only for taxonomic identification, but also for phylogenetic and paleoecological reconstructions; the previously misidentified metapodial specimens in Nihewan fauna were reconsidered in this paper. In the SSMZ fauna, the bovid guild is dominated by Gazella and Bison, which indicates steppe was the most important biome in Nihewan Basin during Early Pleistocene.
Key words Shanshenmiaozui, Nihewan, China; Early Pleistocene; small-medium bovid; new fossils
1 Introduction
Nihewan (=Nihowan which is abbreviated as “NIH” in the MNHN collections) Basin is located at Yangyuan County of Hebei Province in North China, which is famous for its late Cenozoic fluvio-lacustrine deposit sequence and the Early Pleistocene mammalian fauna—the Nihewan Fauna. Nihewan fauna was discovered in the 1920s and published in 1930 by Teilhard de Chardin and Piveteau. The early excavations were conducted mainly in the areas around the Xiashagou and Nihewan villages on the north bank of the Sanggan River(Sangkanho).
In the past decades, much more explorations and excavations have been conducted,and almost all of them were took place on the southern bank of the Sanggan River, and the Shanshenmiaozui Site (Fig. 1) is among the most productive sites in mammalian fossils.Up to now, 25 species, including undetermined species, have been recognized and/or published:Allactaga sibirica(Tong et al., 2021),Ochotonoides complicidens(Tong et al.,2021),Ochotona youngi(Tong et al., 2021),Nyctereutessp.,Canis chihliensis(Tong et al.,2012, 2021),Homotheriumsp.,Acinonyxsp.,Lynxsp.,Pantherasp.,Pachycrocuta licenti,Mammuthus trogontherii(Tong, 2012; Tong and Chen, 2016; Chen and Tong, 2017; Tong et al., 2021),Hipparionsp.,Equus sanmeniensis,Coelodonta nihowanensis(Tong and Wang,2014),Elasmotherium peii(Tong et al., 2014, 2018),Sus lydekkeri,Paracamelus gigas(Tong et al., 2021),Nipponicervus elegans(Tong et al., 2021),Eucladoceros boulei(Tong and Zhang,2019),Elaphurus bifurcatus(Tong et al., 2021),Spirocerus wongi,Gazella sinensis,Ovis shantungensis,Megalovis piveteaui,Bison palaeosinensis(Tong et al., 2017). Among the 25 taxa mentioned above, 23 of which once appeared in the classical Nihewan fauna (=Xiashagou fauna), which account for 92% of the SSMZ fauna.

Fig. 1 Location of Shanshenmiaozui (SSMZ) site, with distributional map of living antelopes and related fossil forms of the Pleistocene Epoch in China Data for the extant taxa are from Jiang (2004), Smith and Xie (2008). Fossil localities:1. Xiashagou of Nihewan; 2. Yushe; 3. Dongdoubi of Yuxian; 4. Jinyuan Cave; 5. Zhoukoudian Loc.1;6. Xujiayao; 7. Mashandong; 8. Salawusu; 9. Xifeng of Qingyang; 10. Loufangzi; 11. Jinniushan;12. Miaohoushan; 13. Haimao of Dalian; 14. Gulongshan; 15. Xiaogushan; 16. Chicheng; 17. Dingcun;18. Xihoudu; 19. Tianshuigou of Dali; 20. Wenxi; 21. Pinglu; 22. Danangou of Yuxian; 23. Bajiazui;24. Gonghe; 25. Yangguo; 26. Tuozidong; 27. Tunliu; 28. Banpo; 29. Linyi; 30. Yuanmou; 31. Heshui;32. Xingtai; 33. Xiaochangcun; 34. Wuwang of Linyi; 35. Gengjiagou; 36. Luhuo; 37. Zhoukoudian Loc.15;38. Upper Cave; 39. Guxiangtun; 40. Zhoujiayoufang; 41. Dali Man site; 42. Aba; 43. Chifeng;44. Rouyuan; 45. Yanjiagang; 46. Yuhongcun of Dali; 47. Lingjing

Fig. 2 Partial skulls, jaw bones and teeth of Spirocerus wongi (A-D) and Ovis shantungensis (E) from SSMZ A-D. Spirocerus wongi: A. partial skull with horn-cores (IVPP V 28650), B1-B3. juvenile maxilla with DP2-M1 (V 28651), C1-C4. left mandible with dp2-m1 (V 28652.1), D1-D4. partial right mandible with p4-m3(V 28653); E1-E2. Ovis shantungensis, maxilla with left DP4-M1 and right DP2-4 and M1 (V 28693)A. anterior view; B1, E1. palatal views; B2, C1, D1, E2. buccal views; C2, D2. lingual views;B3, C3-4, D3-4. occlusal views. The unmarked scale bars equal 20 mm
In the SSMZ fauna, the dominant species includeCanis chihliensis,Mammuthus trogontherii,Equus sanmeniensis,Coelodontanihowanensis,Sus lydekkeri,Eucladoceros boulei,Spirocerus wongi,Gazella sinensisandBison palaeosinensis, most of which have gotten published; the present paper is going to focus on the study of the small and mediumsized bovids, all of which are regarded as having the same geologic horizon.
The SSMZ site lies at the neighboring hill of Xiaochangliang, a well-known Paleolithic Site in Nihewan Basin. Based on the stratigraphic correlation in the field, the fossil-bearing sand-silt bed at SSMZ site is a little higher than the cultural layer at XCL site (Tong et al.,2011; Liu et al., 2012), whose paleomagnetic age is about 1.36 Ma BP (Zhu et al., 2001). On the other hand, the SSMZ fauna is very similar with the classical Nihewan fauna in faunal composition, which means the SSMZ fauna should have an approximate age as the latter based on faunal correlation (Tong et al., 2021), whose recent dating result is 2.2-1.7 Ma (Liu et al.,2012).
2 New fossil materials and the extant specimens compared
The information of all of the studied specimens from SSMZ is listed in Table 1. All of the fossils are reposited in IVPP.

Table 1 Studied fossil specimens of small to medium-sized bovids newly unearthed from SSMZ
The compared local extant antilopines include the following species as defined by Jiang(2004):Gazella subgutturosa,Procapra picticaudata,P. gutturosa,P. przewalskii,Pantholops hodgsoniiandSaigatatarica. The compared extant specimens are from the following collections: Institute of Vertebrate Paleontology and Paleoanthropology, CAS; Institute of Zoology, CAS; Northwest Institute of Plateau Biology, CAS.

Continued
3 Methods, terminology and abbreviations
The suprageneric classification of Bovidae is after Pilgrim (1939), Sokolov (1953),Gentry (1992, 2010), McKenna and Bell (1997), Vrba and Schaller (2000), Grubb (2001),Matthee and Davis (2001), Kuznetsova and Kholodova (2003), Hassanin et al. (2012); the taxonomy at generic level is after Groves (2014) and Castelló (2016); the classification of the local Antilopinae and Caprinae is after Jiang (2004).
The osteological terms are after Reshetov and Sukhanov (1979), Gee (1993), Brown and Gustafson (1979). The terms of tooth nomenclature are after Janis and Lister (1985), Gentry(1992, 2010), Bärmann and Rössner (2011). Upper case letters are used for upper, and lower case letters for lower teeth respectively.
The specimens were measured according to the methods used by von den Driesch(1976). The measurement of horn-core is after Young (1932) and Ratajczak et al. (2016). The measurements were taken with sliding calipers in millimeters. The internal structures of the horn-cores were examined by using the CT scan machine (450ICT) at IVPP.
The biochronological framework follows Tong et al. (1995) and Qiu (2006).
Institutional and locality abbreviationsCAS, Chinese Academy of Sciences; GWL,Gongwangling; HPICR, Hebei Province Institute of Cultural Relics; IOZ, Institute of Zoology,CAS; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, CAS; MNHN,Muséum National d'Histoire Naturelle; NM, Nihewan Museum; NNNRM, Nihewan National Nature Reserve Management; NWIPB, Northwest Institute of Plateau Biology, CAS; OV,Prefix to the catalog numbers of extant specimens in IVPP; SIZ, Shaanxi Institute of Zoology;SSMZ, Shanshenmiaozui; TNHM, Tianjin Natural History Museum; V, Prefix in the catalog numbers for vertebrate fossils in IVPP; XCL, Xiaochangliang; XSG, Xiashagou; YS, Yushe.
Morphological abbreviationsAPD, anteroposterior diameter; TD, transverse diameter;Mc, metacarpal; Mt, metatarsal.
4 Systematic paleontology
4.1 Spiral-horned antelope
Class Mammalia Linnaeus, 1758
Order Artiodactyla Owen, 1848
Suborder Ruminantia Scopoli, 1777
Infraorder Pecora Linnaeus, 1758
Family Bovidae Gray, 1821
Subfamily Antilopinae Baird, 1857 (=Gazellinae Coues, 1889)
Tribe Antilopini Gray, 1821
Genus SpirocerusBoule & Teilhard de Chardin, 1928
Spirocerus wongi Teilhard de Chardin & Piveteau, 1930
MaterialsSee Table 1.
Skull bones: One partial adult skull (Fig. 2A) and one partial juvenile skull (Fig. 2B1-3)can be referred to the the speciesS. wongi. The adult one has very limited frontal bone, but with almost complete horn-cores of both sides. In front view, the frontal bone shows two moderately developed supraorbital pits which accommodate the two prominent supraorbital foramina which are connecting directly to the anterosuperior corner of the orbit and each has a very large internal opening, even larger than in an ox. The eye socket is very large and deep,and has a trench-like roof with flat superior wall rather than domed as in other taxa. The horn is very close to the orbit, which is only 30 mm apart; the postorbital constriction is also obvious.The cranial width at the horn bases is 149.0 mm, and the two horn-cores are 65.6 mm apart at the base. The frontal sinus (pneumatization) is also quite developed, but the diverticulum doesn’t extend to the cornual process (Fig. 3A2).
Horn-core: The horn-core is a typical form ofSpirocerus wongi, with rugose surface and only one spiral carena (or carène or keel) (Fig. 3B1-4), rather than two as inS. peiiandS. kiakhtensis. The carena derives from the basal anterior part and heteronymously spirals upwardly (the left horn spirals anticlockwise but the right one spirals clockwise) and ends near the tip after finishing one complete revolution. The pedicle is quite short. The interior of the horn-core is nearly solid but with a complicated minor canal system (Fig. 3A1-2, C1-5),which resembles those of the true gazelles. The cross-section of the horn-core is elliptical(Fig. 3C1-5). Around the basal part of the horn-core, there are many nutrient foramina, among which the ones above the orbit are the largest (Fig. 3B2). The dimensions of the horn-cores are shown in Table 2.

Table 2 Measurements of the horn-cores of Spirocerus wongi, compared with related species (mm)
Maxilla: One juvenile maxilla with DP2-4 and unfully erupted M1 on both sides (Fig.2B1). Except the trench-like anterior part, the palatal surface is quite flat; the palatal widths between DP2, DP3, DP4 and M1 are 37.0, 47.1, 49.0 and 58.9 mm respectively.
Mandible: One partial adult mandible with p4-m3 (Fig. 2D1-4) and two juvenile partial mandibles (Fig. 2C1-4) are referred to the speciesS. wongi. The two juvenile mandibles(IVPP V 28652.1 and V 28652.2) should belong to the same individual because of their exactly similar dental features. One of the juvenile mandible (V 28652.1) retains most of the mandibular body and the ramus, but lacks the angular process and the tip of the coronoid process. The adult partial mandible has the mandibular condyle completely preserved. All the mandibular specimens have no speciality except the moderately pachyostosed mandibular body (Fig. 2C3, D3), which is similar with those of the extant sumatran serows (Capricornis sumatraensis).
Deciduous upper teeth (Fig. 2B3; Table 3): All of the deciduous upper cheek teeth are completely molarized except DP2. DP2 is elongated, but the posterior lobe is relatively reduced and the posterior fossette (or infundibulum) is shallow. The DP3 has much more wider but shorter posterior lobe relative to the anterior one, and the two fossettes are V-shaped in occlusal view; the mesostyle rib is quite pronounced, the lingual wall of the first lobe is roundish. The DP4 has a much more wider anterior lobe relative to the posterior one; the mesostyle rib is also quite pronounced; the lingual wall of the posterior lobe is more roundish relative to the anterior one. The metacone rib is less developed in all three kinds of tooth. The dimensions of the deciduous teeth are shown in Table 3.
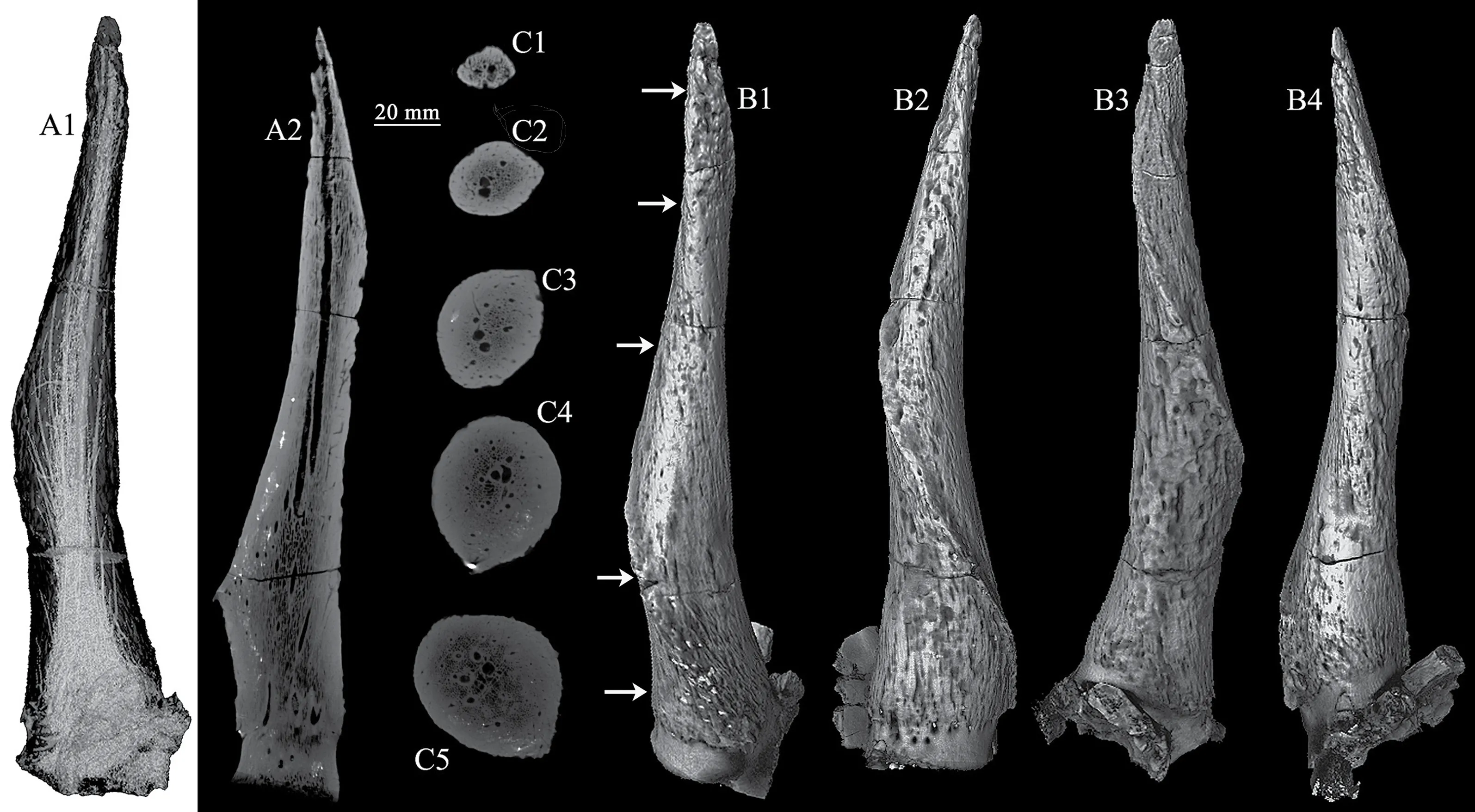
Fig. 3 CT scan images and 3-D reconstructions of the horn-core of Spirocerus wongi from SSMZ (IVPP V 28650)A1-A2. CT scan images showing the general canal system (A1) and a longitudinal slice (A2);C1-C5. CT scan slices showing the changes of cross sections and the canal system at different levels;B1-B4. CT image reconstruction of the right horn-core in anterior (B1), lateral (B2), posterior (B3) and medial (B4) views
Deciduous lower teeth (Fig. 2C1-4): The dp2 is less complicated; the crown has a rectangle outline but the front portion becomes slightly narrower; the paraconid is crest like;there is no parastylid; the protoconid is robust; the metaconid and hypoconid are equally developed; the entoconid is not developed. Between the protoconid and the metaconid, there exists a shallow valley. The dp3 is much more complicatedly constructed, and larger than dp2(Fig. 2C4); the outline is in narrow trapezoid, with the lingual side concave and the narrow portion toward anterior; in front of the paraconid there is a parastylid; the metaconid is very elongated and extended backwards; the valley between parastylid and paraconid is closed, but the valley between the paraconid and metaconid is open; the buccal wall is flat but slightly convex, and the hypoflexid is quite shallow. The dp4 is the most complicated and longest tooth which has three lobes, each lobe has a separate infundibulum, namely anterior, posterior and third fossettid respectively, and the three lobes are nearly equally developed but the second one is slightly more expanded; according to the nomenclature by Bärmann and Rössner (2011), the anterior lobe consists of anterobuccal conid and anterolingual conid, the middle lobe consists of protoconid and metaconid, and the posterior lobe is composed of hypoconid and entoconid respectively; on the buccal side, there is no basal pillars. The lingual cuspids, the parastylid,mesostylid and metastylid are well developed.
Permanent lower teeth (Fig. 2D1-4; Table 3): Only one partial mandible preserves p4 to m3. The p4 is very advanced in well developed protoconid and metaconid, but other cuspids are extremely reduced; metaconid is column-like but connects with paraconid mesially by a narrow bridge, it doesn’t contact protoconid; there is a narrow and deep groove between metaconid and entoconid. The m1 and m2 are very similar in morphology, except the latter is longer and has more developed goat fold; the infundibulums are very narrow; the lingual cuspids are compressed linguobuccally and the lingual wall is quite flat. The m3 has three lobes, the front two of which are very similar to m2 in shape and size; the third lobe has no infundibulums and is linguobuccally compressed. From the lingual aspect, prominent parastylid and entostylid ribs on m2 and m3 can be seen, and even a rib at the linguo-distal corner of hypoconulid lobe can also be observed, but the mesostylid rib is only slightly developed at the top part of the crown. From the buccal view, no basal pillars can be observed.
Postcranial bones: See Table 1.

Table 3 Measurements of the teeth of Spirocerus wongi, compared with related species (mm)
Mc III+IV: The Mc III+IV (Fig. 4A1-3) is a kind of short but robust one, with the distal end expanded. The cranial aspect is more rounded on the shaft while the caudal surface is more flat. At both the proximal and distal ends the medial side or Mc III looks prominently thicker than the lateral side. In proximal view, there are two large articular facets, the larger facet or magnum trapezoid facet is relatively wide and has a curved edge, which is articulated with the fused 2nd and 3rd carpals; the smaller facet or unciform facet is triangular and articulates with the 4th carpal; between the two facets there exists a groove or depression. The posterior medial tubercle is not developed. In anterior view, the vascular groove (or metacarpal gully)is nearly unobservable and the nutrient foramen is tiny. In posterior view, the shaft has a quite flat surface, except near the proximal end, there is a depression; a small nutrient foramen exists near the distal articular surface. The dimensions are shown in Table 4.
Naviculo-cuboid (IVPP V 28656.1): Except the astragalus and calcaneum, a complete set of right tarsal bone was preserved, which still articulates with the metatarsal bones (V 28656.4) in situ. In proximal view, the articular surface consists of a large trochlear facet with a pair of depressions and a distinct posterior beak-like projection (or internal process) at the medioposterior corner, but the internal process is not developed; a narrow sloped belt-like facet occupies the lateroposterior part, which articulates with the calcaneum; there is a tiny foramen at the bottom of the lateral depression which corresponds with the lateral condyle of the distal trochlea of the astragalus. In distal view, the general outline of the distal surface is that of a rounded quadrilateral with two large anterior articular facets and two small posterior facets; the anteromedial facet is articulated with the fused 2nd and 3rd tarsals; the smallest facet articulates with the 1st tarsal; the other two lateral facets articulated with the Mt III+IV directly. In anterior view, the facet articulated with fused 2nd and 3rd tarsals is higher in position than other distal facets; the calcaneal facet is sloping upward anteroposteriorly. In posterior view, there is a large projection at the posteromedial corner; the posterior surface is not flat and with some small nutrient foramina; there exists a notch between the medial and the calcaneal facets, and the latter extends downward along the back wall (Fig. 4B4). The TD is 43.4 mm, and the APD is 43.8 mm.
The fused 2nd+3rd tarsals (IVPP V 28656.2) (Fig. 4B2-3): Its anatomical position is under the anteromedial portion of the naviculo-cuboid. The proximal articular facet is concave,but the distal facet is convex. Its form is anteroposterior elongated, and its greatest dimension is 29.5 mm.
The 1st tarsal (or medial cuneiform): Short and pillar-like, with facets at both ends; the navicular facet is larger but depressed, and the Mt III facet is smaller but flat. Its anatomical position should be under the beak-like projection of naviculo-cuboid (Fig. 4B2-3).

Table 4 Measurements of metacarpals of bovids from Nihewan Basin, compared with related taxa (mm)Mc III+IV Taxa Sources Length Proximal width Proximal APD Distal width Distal APD Midshaft width Midshaft APD Index of stoutness (Distal width/total length) (%)Bison palaeosinensis Teilhard and Piveteau (1930)231-248 72-74 65 28-29 Tong et al. (2018)226-233 60-67.8 35-38.6 59-63.5 34.2-34.7 36-39.5 26-27.6 26-28 Megalovis latifrons Viret (1954)200 56.5 60 30 Megalovis latifrons Radulesco and Samson (1962)192 56 64 35.7 33.3 Megalovis piveteaui This paper 212 59 37 59 32 34 25 27.8 Teilhard and Piveteau (1930); Schaub(1937); Radulesco and Samson (1962)192-204 51-57 54-61 28-30#Megalovis wimani Schaub (1937)185 5241 5742 30.8#Ovis shantungensis This paper 221 29 27 26 20 19 Teilhard and Piveteau (1930)231-233 42-44 42-44.5 18-19 Ovis ammon Fedosenko and Blank (2005)218 Ovis shangi Teilhard and Young (1936)150 2647 28 17 18.7 Spirocerus wongi This paper 175 32 56 29 31 19 32 Spirocerus kiakhtensis Sokolov (1959)170 60.9#35.0#22.0#35.8 Gazella cf. G. lydekkeri Zhang and Yang (2016)141 Gazella sinensis This paper 180 22 17 23 16 12.5 11.3 12.8 Gazella subgutturosa Demircioglu and Ince (2020)146.0(F)18.4(F)18.0(F)10.5(F)9.7(F)150.7(M)19.6(M)18.9(M)11.4(M)9.9(M)Procapra przewalskii Boule et al. (1928)142 18-21 12.7-14.8 Procapra gutturosa Boule et al. (1928)180 23 12.8 Procapra gutturosa This paper (OV 1124)146 15.5 8.0 16.7 12.1 8.7 7.0 11.4 Saiga tatarica Prat (1966b)153 25 18.5 25.5 13.5 17.7 Capra ibex Prat (1966a)126-15528.5-38.5 20.5-2731.5-43 19-24 20-27.5(139.2)(33)(23.8)(36.7)(21.3)(23.8)Praeovibos priscus Kahlke (1964)193.7#Ovibos moschatus sussenbornensis Kahlke (1961)189.9#62.8#36.5#69.1#44#36.4 Mt III+IV Spirocerus wongi This paper 173.0 36.9 34.0 45.3 26.1 22.0 21.5 26.2# Calculated by the first author of this paper; mean values in parentheses.
Mt III+IV (Fig. 4C1-3): In general view, the Mt III+IV has the very proximate length as the Mc III+IV, which is quite different from the common ungulates whose metatarsals are prominently longer than metacarpals belonging to the same individual; furthermore, the distal end expands more sharply relative to the shaft. In proximal view, the proximal surface has a polygon outline, and consists of four articular facets; the two anterior facets are large and kidney-shaped, but the two posterior facets are small and elongated. The mediolateral dimension of the proximal end is slightly larger than the anteroposterior dimension. The two large articular facets meet at the front middle part but diverge posteriorly; both of them have longer anteroposterior dimensions. The anteromedial facet articulates with the fused 2nd and 3rd tarsals; the anterolateral facet articulates with naviculo-cuboid. There are two smaller facets located near the posterior margin. The posterolateral facet has larger mediolateral dimension and articulates with naviculo-cuboid. The small roundish facet articulates with the 1st tarsal. The dimensions are shown in Table 4.

Fig. 4 Postcranial bones of small and medium-sized bovids from SSMZ A-C. Spirocerus wongi: A1-A3. left Mc III+IV (IVPP V 28655),B1-B4. right naviculo-cuboid+lat-mid+medial cuneiforms (V 28656.2-3),C1-C3. right Mt III+IV (V 28656.4); D-I. Gazella sinensis: D1-D3. partial left humerus (V 28687),E1-E4. left Mc III+IV (V 28688), F. distal epiphysis of left tibia (V 28691),G1-G2. left astragalus (V 28692), H1-H2. left naviculo-cuboid (V 28690), I1-I3. 3rd phalanx (V 28689);J-K. Megalovis piveteaui: J1-J4. left radius (V 28654), K1-K4. right Mc III+IV (V 28695);L1-L4. Ovis shantungensis: left Mc III+IV (V 28694). A1, C1, D1, E1, G1, J1, K1, L1. anterior views;A2, B4, C2, D2, E2, G2, J2, K2, L2. posterior views; A3, B1, C3, E3, H1, J3, K3, L3. proximal views;B2, D3, E4, F, H2, J4, K4, L4. distal views; B3. medial view; I1. lateral view; I2. interdigital view;I3. volar view. The arrows indicate the lateral projection of the radius. The unmarked scale bars equal 20 mm
In posterior view, a very small nutrient foramen positioned slightly off-center some way under the posterior apophysis. Very shallow central trough only can be observed at the midshaft.
At the distal end, there are two distal condyles whose intercondylar margins are parallel.On the anterior surface, the vascular groove (or midline dorsal groove) is wide but shallow and opens to the intercondylar groove.
Comparison and discussionThe degree and direction of horn-core torsion, the type of spiraling and the development and position of the keels are the crucial characters to classify the fossil spiral-horned antelopes (Kostopoulos, 2004). In the early publications, the torsion direction (clockwise or anticlockwise) always means for the right horn (Pilgrim, 1939).Moreover, the recent publications prefer to use “homonymous” and “heteronymous”, if the left horn twists clockwise, the horns are termed homonymous, or else it’s heteronymous.
Spiroceruswas established by Boule and Teilhard de Chardin in 1928, and included the specimens from Sjara-osso-gol (=Salawusu) of Nei Mongol (Inner Mongolia) to it. Later on,the similar fossils from Nihewan (Teilhard de Chardin and Piveteau, 1930) and Yushe (Teilhard de Chardin and Trassaert, 1938) also were referred to this genus.Spiroceruswas originally included into the subfamily Tragelaphinae by Teilhard de Chardin and Trassaert (1938), later on, it was referred to the subfamily Caprine (Sokolov, 1953); while currently it is usually placed in the subfamily Antilopinae (McKenna and Bell, 1997; http://taxonomicon.taxonomy.nl/TaxonTree.aspx?src=0&id=68446). More recent assignment is to Bovinae (Bai et al., 2019).Up to now, the classifications ofSpirocerusat the species level are mainly based on horncore characters, which caused quite a lot of confusions. Kahlke (1999) proposed that the most primitiveSpiroceruswith only anterior carena should be referred toS. wongi. Concerning the bicarinate forms with longer and slender horn-cores, the stratigraphically older ones can be referred toS. kiakhtensispeii, while the younger materials with correspondingly shorter and more massive horn-cores toS. kiakhtensiskiakhtensis. Moreover, all of the species ofSpirocerushave heteronymous spiraled horns.
In China, three species were recognized, namelyS. wongi,S. peiiandS. kiakhtensis; the former one only has one spiral carena while the latter two have two spiral carenas on each horn-core (Teilhard de Chardin and Trassaert, 1938). This genus mainly limited to northern China, Trans-Baikal region and the central Asian countries. FromS. wongiviaS. peiitoS.kiakhtensis, the size is getting smaller and the horn-cores are becoming straighter. Moreover,the current knowledge is mainly about the horn-cores, whereas the cranial features of the genusSpirocerusis still unknown; the dental and postcranial characters are still insufficiently studied yet, because no dentition was found attached to a skull with horn-cores in the fossil record. Relative to its cranial size,Spirocerusshould have big postcranial bones, but the partial skeleton associated with skull materials ofS. kiakhtensisdiscovered near Ulan-Ude of Russia showed that the limb bones of this kind of animals are quite short but robust; therefore, it was inferred thatSpirocerushas big skull but short limbs and similar body-build as the extant muskox does (Sokolov, 1959).
It has to be indicated that the maxillary bone described by Teilhard de Chardin and Piveteau (1930:pl. XII, fig. 1) (current catalog No. NIH 136) should be rechecked, because it seems that the DP4 was mistaken as M1, which can be easily verified by a CT scanning.Furthermore, the Nihewan limb bones identified asS. wongiand Ovibovine gen indet.respectively by Teilhard de Chardin and Piveteau (1930) should be reconsidered; the metapodials referred toS. wongi(pl. XII: figs. 2 and 4) are too long, they should belong toOvis shantungensis; the metapodials referred to Ovibovine gen indet. (pl. XIII: figs. 3 and 4)have the similar form and size as those ofM. piveteaui.The metacarpal from SSMZ described in this paper has almost exactly the same form and same size as that ofS. kiakhtensisfrom the Trans-Baikal region (Sokolov, 1959).
The lower teeth from SSMZ are different from those of Xiashagou in their slightly less developed goat folds and lack of the basal pillar in m1.
It’s necessary to say that until now, none of the dental materials ever discovered forS. wongiwas directly associated with the horns; that’s to say all of the previous and current descriptions about the teeth ofS. wongican be open to questions. It’s lucky enough that some cranial specimens ofS. kiakhtensishave their horn-cores and teeth preserved jointly (Dong et al., 2009). These specimens are very crucial for the morphological studies of the extinctSpirocerus.
Spirocerushas developed frontal sinuses, but they do not extend to the horn-core at all (Fig. 3A2). According to Farke’s paper of 2010, the frontal sinus doesn’t always have phylogenetic significance. But the present authors think it’s more important if the frontal sinus extends to the horn-core, i.e. if there is a cornual diverticulum of the frontal sinus or if the horn-core is hollowed.
Spirocerusis typical for its spiral and straight horn-cores, but sometimes it could be confused withAntilospira zdanskyi; some of the specimens (IVPP V 24483.4, V 25878.1-2)referred toSpirocerus(Bai et al., 2019) possibly belong toAntilospiraor some other taxon because of their un-straight horn-cores and the different carena feature (with prominent groove alongside the spiral carena). Hermier et al. (2020) proposed that the horn-cores inAntilospiraare variably inclined backwards and divergent but their surface usually shows much deeper longitudinal grooves.Antilospirais the most similar contemporaneous taxon withSpirocerus,and it was also uncovered in the Nihewan Basin, but it can be distinguished from the latter by the following characters: un-straight horn-cores, usually with two grooved and slowly twisting carenas (Teilhard de Chardin and Trassaert, 1938).
The comtemporaneous genusGazellospiraalso has similar horn-cores, but it was only discovered in Europe and northern Asia not including China. Furthermore,Gazellospirahas two spiraled keels of which the posterior one is the sharpest (Hermier et al., 2020), which are less straight as those ofSpirocerus. On the other hand, the metapodials ofGazellospiraare completely different from those ofSpirocerus, the formers’ are much longer and slender(Hermier et al., 2020; Vislobokova et al., 2020). Furthermore, the lower p4 ofGazellospirahas its lingual valleys open (Duvernois and Guérin, 1989), while the anterior valley is closed inSpirocerus. Although the intraspecific variations in cranial and postcranial features have been recognized between different sexes and different latitudes forGazellospira torticornis, its horncores and metapodials as well as the lower p4 are distinctly different from those ofSpirocerus.
The other comtemporaneous genusPontocerosis typical for its homonymous torsion of horn-core, which is different from other Early Pleistocene spiral-horned antelopes, includingTragelaphus,GazellospiraandSpirocerus(Vislobokova and Titov, 2020).
Concerning the taxonomic position of the extinctSpirocerus, it’s still controversial.Its large supraorbital foramina (generally sunken in large frontal depressions) and horn-core characters (straight, solid, moderately spaced, twisted clockwise and situated directly over the orbits) support an attribution to the subfamily Antilopinae, but its solid and quite divergent horn-core, lack of post-cornual fossa, short but stout metapodials and the limb proportions share more similarities with the ovibovines under the subfamily Caprinae. Sokolov (1959) postulated that the extinctSpirocerusshould have a similar appearance as muskox (Ovibos). It’s a pity that the fossil records for this animal are too few to reach a definite phylogenetic approach currently.Although some recent publication attributedSpirocerusto the subfamily Bovinae Gill, 1872 (Bai et al., 2019), it seems that few morphological characters can support this proposal.
4.2 Chinese gazelle
Genus GazellaBlainville, 1816
Gazella sinensisTeilhard de Chardin & Piveteau, 1930
MaterialsSee Table 1.
Skull bones: One partial adult skull (Fig. 5B1-3) and one partial juvenile skull (Fig. 5A)can be identified asGazella sinensis. The adult specimen has partial frontal bone preserved,but with almost complete horn-cores of both sides. In front view, the frontal bone has two large supraorbital foramina which are situated in triangular-shaped depressions of the frontals(supraorbital pits) near the base of the horn pedicels and closely above the orbit, they connect directly to the eye socket and have big roundish internal openings. At the lateroposterior aspect of the basal part of the pedicle, there exists a post-cornual fossa (Fig. 5D2, 6A1). The wall of the cranial bone is very thin (Fig. 5B3). The dimensions are shown in Table 5.
Horn-core: The general characters of the horn-cores are short but with moderately long pedicle, without torsion of their axes, and the surface is decorated with longitudinal grooves or striate. In anterior view the two horns are straight and are nearly sub-parallel,i.e. the divergence angle is small (16°-22°) (Fig. 5A, B1, B3); in lateral view they have a gentle backward curvature (Fig. 5D2, E). The horn-cores are solid interiorly but with many longitudinal canals, most of which open upwardly (Fig. 6A1’-A4’, B1’-B3’); at the basal part adjacent to the pedicle, there are a few large foramina and the one just above the post-cornual fossa is the largest (Fig. 6A4’, B3’). The horn-core has a sub-circular cross section with slight transverse compression (Fig. 6A1’-A4’). The horn-cores vary in size (Fig. 5; Table 5) and the development of longitudinal grooves, which may be subject to gender and age difference. The dimensions of the horn-cores are shown in Table 5.
Mandibles: There is no peculiarity in the mandibles. They have short symphyseal length but long diastema between lower canine and p2. The mental foramen is large and lies at the midway of the diastema at the buccal aspect, but sometimes an extraordinary mental foramen will occur below p3. The mandibular depth is shallow under the premolars but becomes deep rapidly as it passes back from beneath the premolars to the molars. The angular process is moderately developed. The coronoid process is long and recurved backward distally. The mandibular condyle is highly superior to dentition. Mandibular foramen is small and the maximum height of opening is at the same horizontal level as the alveolar margin (Fig. 7C3).
Upper teeth: Only two specimens with upper teeth (IVPP V 28673 and V 28674) can be assigned to the speciesG. sinensis. All of the upper cheek teeth are less hypsodont. The P3 is very special in its tendency of molarization by the appearing posterior infundibulum and has greater length than width (Fig. 7A; Table 6). The P4 has only one lobe but without any peculiarity except its developed styles. Each molar consists of two lobes constructed by paracone and protocone, metacone and metaconule respectively; in M1 and M2, the anterior lobe usually has larger width (buccolingual dimension), but in M3 the anterior lobe is narrower than the posterior one. The infundibulum has simple structure and the fold can be seen only in M3. No basal pillar can be observed (Fig. 7A-B).

Fig. 5 Incomplete skulls and horn-cores of Gazella sinensis from SSMZ A. partial skull of a juvenile with horn-cores (IVPP V 28658); B1-B3. partial skull with horn-cores (V 28657);C1-C2. left and right horn-cores (V 28659.1, V 28659.2); D1-D2. partial skull with left horn-core (V 28667);E. partial skull with right horn-core (V 28661). A, B1, C1-2, D1. anterior views; B3. posterior view;B2. dorsal view; E. medial view; D2. lateral view. Scale bars equal 50 mm

Fig. 6 CT scan images of the horn-core of Gazella sinensis (IVPP V 28661) from SSMZ A1-A4. CT image reconstruction showing positions of the cross CT scan slices (A1’-A4’);B1-B3. CT image reconstruction showing positions of the longitudinal CT scan slices (B1’-B3’)
Lower teeth: The lower premolars are less hypsodont than those of the caprinines, but complicatedly constructed, except p2 which is simply constructed and prominently smaller than the other two (Table 6). Lower p3 and p4 has the similar structure, both of them are elongated but p3 has the lingual wall open both mesially and distally (Fig. 7C4, D1, G-H), sometimes with the anterior valley partially closed (Figs. 7D1; 8A); on the other hand, the p4 only has the posterior valley open, like a narrow groove (Figs. 7C4, D3, F-J; 8A) and coincides with the type 5 of p4 patterns proposed by Janis and Lister (1985). Each premolar has two roots. Lower molars are elongate and with developed goat folds, but lack basal pillars; m1 and m2 are very similar in size and shape; m3 has three lobes, but the third lobe has very shallow infundibulum.Molars are quite hypsodont. The m1 and m2 with two roots, but m3 has three roots.
Lower deciduous teeth: The deciduous teeth are brachyodont (Fig. 7E2). The dp2 resembles p2 in shape; dp3 is similar with p3; while dp4 is similar with m3 in having three lobes but is different in having a larger third lobe and with a basal pillar at the buccal groove between the second and the third lobe (Fig. 7E3).
Postcranials: A humerus with partial shaft and complete distal end preserved, the length of the remained part is 73.9 mm (IVPP V 28687) (Fig. 4D1-3). In anterior view, the indentation (radial or coronoid fossa) above the articular surface is shallow; the medial edge of the trochlea is curved and with the upper portion inclined laterally; at the lateral two-fifths part of the trochlear surface, there exists a sagittal ridge whose lower part is nearly vertical but with the upper part slightly inclined laterally; the lowest part of the distal articular surface occurs at the lateral side; there is a longitudinal trench at the middle part of the articular surface; below the articular surface at the medial side,the medial epicondyle can be observed. In posterior view, the shaft has a very straight medial margin and a nutrient foramen at the lateral aspect can be seen;the olecranon fossa at the distal end is very deep; the medial epicondyle extends much more downward than the lateral one. The distal APD is 27.3 mm; the distal TD is 29.7 mm.
Mc III+IV: A complete left Mc III+IV (IVPP V 28688) (Fig. 4E1-4). In general, the bone is very slender; the cranial aspect is more rounded on the shaft and the caudal aspect is much flatter and slightly concave at the upper part; the distal epiphysis is not fused yet. In proximal view, there are two articular facets, the larger one or magnum trapezoid facet is prominently wider and deeper, which articulates with the fused 2nd and 3rd carpals; the smaller one or unciform facet is oval-like and is the articular surface for 4th carpal; between the two facets there exists a narrow ridge; no nutrient foramen can be observed at the proximal end (Fig. 4E3). In anterior view, a prominent tubercle (extensor carpi radialis insertion:Gentry, 1966: fig. 12) occurs at the cranial aspect of the proximal end; no vascular groove (or metacarpal gully) can be observed, while the nutrient foramen is clear but small and located near the epiphyseal suture;the widest portion lies at the epiphyseal suture rather than at the trochlea (Fig. 4E1). In posterior view, no prominent groove can be observed except the upper part; the articular facet for the vestigial 5th metacarpal is absent. A prominent nutrient foramen exists near the epiphyseal suture and a tiny nutrient foramen occurs at the mid-shaft (Fig. 4E2). The dimensions are shown in Table 4.

Fig. 7 Jaw bones and teeth of Gazella sinensis from SSMZ (A-J), compared with related taxa (K-P)A. maxilla with P3-M1 (IVPP V 28673); B. maxilla with M1-3 (V 28674);C1-C4. left mandible with p2-m3 (V 28680); D1-D3. left mandible with p2-m3 (V 28681.2);E1-E2. left mandible with dp2-4 and m1-2 (V 28685.1), E3. detail of dp2-4; F. right p4-m3 (V 28677);G. left p3-m3 (V 28682); H. right p2-m3 (V 28675); I. left p4-m3 (V 28686); J. left p4-m3 (V 28678);K. Procapra przewalskii, right p2-m3 (horizontally flipped) (NWIPB-0001172♀);L. Procapra gutturosa, left p2-m3 (NWIPB 620032); M. Gazella subgutturosa, left p2-m3 (NWIPB 609001);N. Pseudois nahaur, left p2-m3 (NWIPB-KX1); O. Capra ibex, left p2-m3 (IOZ-2);P. Ovis ammon, left p2-m3 (OV 1346-2). A, B, C1, C4, D1, E3, F-P. occlusal views;C2, D2, E1. buccal views; C3, D3, E2. lingual views. The arrows indicate the variations of p4 The unmarked scale bars equal 20 mm

Table 5 Measurements of partial cranial bones and horn-cores of Gazella sinensis (mm)IVPP V 28657 V 28658 V 28659.1V 28659.2V 28660V 28661V 28662V 28665V 28667V 28670V 28671V 28672 Greatest breadth across orbits: ectorbitale-ectorbitale>81.8>80.9 Width between supraorbital foramina 30.9 35.0 Frontal angle 34°Angle of divergence of horn-cores 16°22°Width between horns 65.8 66.6 Minimum distance between inner margins of horn processes 23.2 29.0 Diameter of horn-core base (APD)28.4 24.9 26.3 26.6 27.2 27.0 27.7 25.3 28.2 25.9 27.6 24.7 Latero-medial diameter of horn-core base (TD)21.3 19.5 21.3 21.4 21.3 20.8 20.4 19.6 21.7 19.8 21.4 17.5 Full curved length of horn-core 170 118 146 135 130 132 135>101 152 131 Straight length of horn-core 160 110 132 123 120 117 120>95 140 116

Table 6 Measurements of teeth of Gazella sinensis from SSMZ, compared with those from Xiashagou (mm)IVPP V 28675 28676286732867428677 28678 28679 28680 28681.1 (r)28681.2 (l)V 286822868328684 28685.1 (l)28685.2 (r)28686 Teilhard & Piveteau(1930)P3 6.9 LWLWLWLWLW 6.4 P4 8.3 7.8 M1 11.0 9.8 10.9 12.7 M2 13.4 12.9 M3 15.1 10.3 M1-M3 LLWLWLW 38-45 dp2 5.2 4.9 2.5 2.5 dp3 8.5 7.5 4.3 3.5 dp4 12.9 13.8 5.4 5.3 dp2-dp4 L 26.4 27.1 p2 5.3 5.6 6.0 6.4 5.8 LWLWLW 3.0 3.7 3.1 3.2 3.1 p3 8.2 8.4 9.1 8.4 8.3 9.6 3.9 4.2 3.8 4.4 4.2 4.5 p4 8.6 8.7 8.5 7.9 8.1 8.9 8.7 9.7 7.8 7.9 4.5 5.1 5.1 4.5 4.3 4.7 4.7 5.0 4.6 5.0 p2-p4 LLWLWLW 22.2 22.3 20.8 21.5 22.5 22.4 22-28 m1 10.1 10 11.5 10.2 10.6 10.0 11.3 12.9 11.5 9.4 10.3 13.4 13.7 10.1 5.9 6.5 7.2 6.8 6.6 6.6 6.7 7.8 5.2 7.1 6.8 6.8 7.1 m2 12.1 12.0 14.2 12.5 11.9 13.6 13.8 13.4 11.8 13.0 14.9 15.4 12.6 7.2 7.0 7.7 7.5 7.8 7.2 7.1 7.1 8.0 7.7 7.7 6.9 6.9 7.8 m3 19.4 19.1 21.9 19.0 19.2 18.6 20.3 20.2 19.4 19.4 19.2 20.1 7.3 7.5 7.2 7.5 7.6 7.5 7.3 7.1 7.7 7.2 7.9 7.6 m1-m3 L 41.8 42.5 46.1 43.1 41.7 46.4 45.7 44.9 39.5 41.9 41.2 43-50 p2-m3 L 64.1 64.0 64.2 66.8 67.0<67.8 69.1 69.4 65-78
Distal epiphysis of tibia: On the distal end (IVPP V 28691) (Fig. 4F), the articular grooves are nearly parallel to the sagittal plane of the shaft, the medial groove is narrow but longer, the lateral groove is wide but shorter; between the two grooves, there exists a sagittal ridge, and the anterior end of the ridge is beak-like and distally projected; at the anteromedial corner, there is a long projection. There are two small articular surfaces on the lateral edge for the attachment of the lateral malleolus (or distal fibula). The APD is 18.5 mm, the TD is 21.3 mm.
Astragalus: A complete astragalus (IVPP V 28692) is preserved (Fig. 4G1-2). In anterior view, there are two proximal trochlear ridges (or condyles), which are parallel to each other and to the sagittal plane, the lateral ridge is prominently higher and wider. Between the two proximal trochlear ridges, there exists a deep trench-like median groove. The lateral border of the trochlear condyles is straight. The distal trochlea also consists of two parts, between them is a shallow saddle-like groove. In posterior view, the posterior articular surface for calcaneum is flat to slightly convex, but with a shallow longitudinal medial groove; the lateral edge is nearly straight; the medial bottom extends further downward and becomes confluent with the distal trochlear surface. The lateral face is trench-like and with anterior and posterior rims;the anterior rim extends upward and beak-like; at the bottom, the facet for calcaneum is ovalshaped. On the medial aspect, there is no articular facet. The length is 29.2 mm, the width is 17.2 mm.
Naviculo-cuboid: A complete naviculo-cuboid bone (IVPP V 28690) is preserved (Fig.4H1-2). In proximal view, two articular surfaces can be observed, one for astragalus, the other for calcaneum; the former consists of a pair of depressions and a distinct posterior beak-like projection (or internal process) at the medioposterior corner; the latter is a narrowly sloped belt-like facet occupies the lateroposterior part; there is a tiny foramen at the bottom of the lateral depression. In distal view, the general outline is that of a rounded quadrilateral with two larger anterior articular facets and two smaller posterior facets. The anteromedial facet is articulated with the fused 2nd and 3rd tarsals; the smallest facet articulates with the 1st tarsal;the other two lateral facets articulated with the Mt III+IV. In anterior view, there is a large projection at the posteromedial corner, which is separated from other part by a deep notch; the facet articulated with the fused 2nd and 3rd tarsals is higher in position than other distal facets;the posterior surface is not flat. The APD is 21.6 mm, and the TD is 22.3 mm.
The 3rd phalanx: One complete distal (or 3rd or ungula) phalanx (IVPP V 28689) (Fig.4I1-3) was unearthed, whose general look is quite narrow. In lateral view, both the dorsal and plantar margins are nearly straight except a slight concave at the middle part of the plantar margin. In proximal view, the dorso-posterior corner is projected; the articular surface can be divided into two facets, which articulate with distal end of 2nd phalanx. In distal view, the plantar surface is nearly flat and triangular and with the acute angle toward the anterior.
Comparisons and discussionsPremolars are crucial for taxonomic identification for the gazelles, especially the tooth structures of the lower p4 which is usually regarded as the most practical tooth in taxonomic distinctions (Boule et al., 1928; Teilhard de Chardin and Piveteau, 1930; Gentry, 1966; Janis and Lister, 1985; Chen, 1997). Among all theGazellaspecies, including both extant and fossil forms, the lower p4 usually has open lingual valleys(Teilhard de Chardin and Piveteau, 1930; Teilhard de Chardin and Trassaert, 1938; Zhang and Yang, 2016; Li et al., 2018) (Fig. 7M, 8E), exceptG. sinensis(Fig. 7C4, D3, F-J; 8A) andG.yushensisChen, 1997; whereas theCapraspecies usually has the anterior valley closed in p4,i.e. paraconid and metaconid joined to form a continuous anterolingual wall (Fig. 7O), and the posterior valley sometimes is also closed. Among all theGazellaspecies,G. sinensishas the most complicated lower p3, some of which have the anterior valleys partially closed lower in crown, and all the cuspids and stylids are well developed, especially the parastylid is very robust, and the metaconid is the most developed relative to other compared Asian gazelles’ (Fig.8A); but most p3s have their lingual valleys open. Because of the preorbital fossa, the fossils represented byG. sinensiswere included into the genusGazella; on the other hand, its lower p4 has closed anterior valley, which is a crucial character ofProcapra; therefore, the speciesG.sinensiswas considered as the direct ancestor ofProcapra(Sokolov and Lushchekina, 1997).
In addition to lower p4, there exist other crucial premolar characters to distinguish the gazelle taxa in China, e.g.Saiga tataricausually lacks andPantholops hodgsoniiexclusively lacks p2 in their dentitions (Fig. 8F-G).
The toothrow length is rather stable than their cranial sizes among the extant gazelles in China (Fig. 9: raw data from Jiang, 2004); although the Mongolian gazelle (Procapra gutturosa) is remarkably larger in body size and cranial length than others, its toothrow length has no prominent difference. Therefore, it seems impossible to distinguish the small gazelles only based on their toothrow lengths (Fig. 9); in addition, the blue sheep also has similar toothrow length with the gazelles; while the wild goat has slightly larger tooth-row length and the sheep has obviously larger tooth-row length than others (Fig. 7P).

Fig. 8 Comparison of premolar series among some Quaternary gazelle species of China A. Gazella sinensis, left p2-4, IVPP V 28681.2, SSMZ; B. Procapra gutturosa, left p2-4, NWIPB 0006065,extant; C. P. przewalskii, right p2-4 (horizontally flipped), NWIPB 0001172, extant;D. P. picticaudata, left p2-4, NWIPB 0001179, extant; E. Gazella subgutturosa, left p2-4, IVPP-c-05, extant;F. Saiga tatarica, right p3-4 (horizontally flipped), NWIPB S-80503, extant;G. Pantholops hodgsonii, left p3-4, NWIPB 77001, extant. All are in occlusal views

Fig. 9 Toothrow length and cranial size of extant gazelles in China Raw data was employed from Jiang, 2004
Furthermore, the horn-cores are highly uniform among the gazelle species, neither the size (Boule et al., 1928: table 13) nor the form can be used independently for specific distinctions. Although the post-cornual fossa of the frontal bone is also a common character among the gazelle species under the generaGazellaandProcaprain China, both fossil and extant, while it’s absent in the species ofSaigaandPantholops, that’s to say post-cornual fossa can be used for taxa distinction only to some extent.Pantholopsis also different from other gazelles by its obviously smaller supraorbital foramen. Up to now, no fossil records ofSaigaandPantholopshave ever been reported in China.
It’s worth mentioning that the upper P3 has some kind of peculiarity in its tendency of molarization, which is a general phenomenon only occurs in some kinds of gazelle taxa, such asG. subgutturosaand the species ofProcapraas well as the fossil forms ofGazella. In p4 morphology,G. sinensisis closer toP. przewalskii.
Concerning the horn-cores of gazelles, they are fairly uniform, not only in size, but also in form and interior structures, and usually have grooved surfaces and are nearly completely interiorly solid, i.e. without cornual diverticulums; therefore, it’s almost impossible to distinguish the gazelle taxa only by horn-cores. On the contrary, the horn-cores of caprines are exclusively hollowed and diversified in shape.
The recent study in frontal sinuses and cranial pneumaticity of the bovid species shows a strong phylogenetic correlation with sinus morphology, except few exceptions (Farke, 2010).The extant species ofProcaprahas no frontal sinus, which is distinctly different from theGazellaspecies.
The metacarpal bone is prominently larger than such allied taxa, asGazellasubgutturosaandProcapra przewalskii, but with the same size as that ofP. gutturosa(Table 4); in morphology, it has no peculiarity. The metapodial bone does have distinctive characters relative to those of the caprines in their extremely slenderness. Meanwhile, the BT (maximum breadth of trochlea) of the metacarpal of SSMZ specimen is 20.9 mm, which stands between those of theGazella(15.1-20.1) andProcapra(21.5-21.6) (Wang et al., 2020).
Gazelles are adapted to open and arid environment, therefore, their range, both fossil and extant, only limited to the northern part in China, and the southernmost occurrences are Xuchang Man site at Lingjing in Henan Province (Dong and Li, 2008) and the Tuozi Cave in Jiangsu Province (Dong et al., 2007) (Fig. 1). As to the “Gazellakaihuaensis” from Zhejiang Province (Jin et al., 1994), it’s an incorrect identification because of the following evidences:larger size, lack of a pedicle, horn far away from orbit, horn-cores with smooth surface rather than with longitudinal grooves, supraorbital foramen too small, small angle between horn-core and cranial roof, none of which coincides with the characters of aGazella. The partial skull bone described by Jin et al. (1994) should belong to a serow (Capricornis).
4.3 Shandong sheep
Subfamily Caprinae Gill, 1872
Tribe Caprini Gray, 1821
Genus OvisLinnaeus, 1758
Ovis shantungensisMatsumoto, 1926
1930Spirocerus wongi, Teilhard de Chardin and Piveteau, p. 71, Pl. XII, figs. 2 and 4
Maxilla: One maxilla with left DP4-M1 and right DP2-M1 (IVPP V 28693) (Fig.2E1-2) can be referred toOvis shantungensis. The DP2 is deeply worn and the infundibulum has disappeared. The DP3 is moderately worn, with robust paracone and metacone as well as metaconule, the protocone is less developed, with robust parastyle, the posterior lobe is wider than the anterior one. The DP4 has equally developed main cusps, and the styles are also equally developed; the paracone rib is very robust, the anterior lobe is wider than the posterior one. In form, the M1 is very similar with DP4, but much bigger and more hypsodont. The dimensions of the teeth are shown in Table 3.
Metacarpal: One almost complete Mc III+IV is available (IVPP V 28694) (Fig. 4L1-4).The peculiarity is its slendness, which is really unuasal for a caprine taxon. In morphology,size and proportions, it’s very similar with the metacarpal of a medium-sized deer, except its less developed vascular groove and less concave midshaft posterior surface. The two large proximal articular facets form an asymmetric semicircular, between them there is a groove which opens postwardly, but the nutrient foramen in the groove is tiny. The metacarpal gully is weak; the situation of the lower nutrient foramen is unclear because of the breakage; the cranial side is more rounded. In posterior view, the shaft has a shallow but broad longitudinal depression at the upper three-fifths of the bone, and the surrounding area is quite rugose, the lower part has quite flat posterior surface; a small nutrient foramen occurs near the distal trochlea. The greatest length is 221 mm, which is very close to that ofO. ammon(218 mm)(Fedosenko and Blank, 2005); the BT (maximum breadth of trochlea) of the metacarpal of SSMZ is 39.3 mm, which falls within the range ofO. ammon(30.1-41.9 mm) (Wang et al.,2020).
Comparisons and discussionsThe DP2 and DP3 have the similar form and size as the counterparts ofO. shantungensisfrom XSG (Teilhard de Chardin and Piveteau, 1930:text-fig.26-B), but the M1 is prominently larger than the type specimen (Matsumoto, 1926). Compared with the deciduous teeth ofS. wongi, Teilhard de Chardin and Piveteau (1930:text-fig. 26) had indicated their differences:S. wongihas much bigger and more elongated teeth, especially the DP3 and DP4; furthermore, it’s also different fromO. shantungensisin its faint cingulum-like structure at the antero-lingual part of DP3, the buccal wall is broad and flat, the lingual wall of DP2 is concave at the posterior lobe.
According to the data of metapodials of variousOvisspecies, both fossil and extant, it seems wrong assignments of the metatarsal bones from Yushe Basin toOviscf.shantungensis(Teilhard de Chardin and Trassaert 1938:83), and from Zhoukoudian Loc.1 toOvissp. (Young,1932:76), because their total lengths are only 135 and 144 mm respectively, which are too short for anOvisspecies.
4.4 Piveteau’s giant sheep
Tribe Ovibovini Gray, 1872
Genus MegalovisSchaub, 1923
Megalovis piveteaui Schaub, 1937
1930 Ovibovine gen. indet., Teilhard de Chardin and Piveteau, p. 76-79, Pl. XIII, fig. 2, 2a, 3-4 1942 ?Budorcussp., Teilhard de Chardin and Leroy, p.82
Radius: A nearly complete left radius (IVPP V 28654) is preserved (Fig. 4J1-4). The radius is quite massive, the body is anteroposteriorly compressed, and with the middle portion curved anteriorly. The most peculiarities lie at the two ends: the proximal end consists of three glenoid sub-fossae, which are increasing in size from lateral to medial; the proximal articular surface of the glenoid fossa has an expanded and rounded medial outline; a deep V-shaped notch occurs at the middle fossa, which articulates with the ulna. The distal end is expanded, but the articular facet is moderate in size; a pronouced and rugose ulna facet lies at the posterolateral corner, which has a sharp projection toward the lateral side (indicated by an arrow in Fig. 4J4), which is a peculiarity ofOvibos(Gromova, 1960); the distal ulna is not fused to the radius. The total length is 271 mm, the proximal width is 64.5 mm, the proximal depth is 35.0 mm, the distal width is 61.0 mm, the distal depth is 42.2 mm.
Metacarpal: A right anterior canon bone (Mc III+IV) (IVPP V 28695) (Fig. 4K1-4). In general, both the proximal and distal ends expand to some extent, and the metacarpal bone is moderately stout. At both proximal and anterior aspects, the medial side or Mc III looks noticeably thicker than the lateral side. The two proximal articular facets are quite flat, and a narrow groove occurs between the facets. In anterior view, the shaft is roundish, and the vascular groove (or metacarpal gully) is very narrow at the upper par but widened around the nutrient foramen. In posterior view, the proximal portion of the shaft has a medial longitudinal depression, but the lower part is flat; a prominent nutrient foramen exists near the articular surface. The dimensions are shown in Table 4.
Comparisons and discussionsBased on the fossils identified as Ovibovine gen. indet.by Teilhard de Chardin and Piveteau (1930), the speciesMegalovis piveteauiwas established by Schaub in 1937; both the maxillary and the metacarpal bones are very similar with the speciesMegalovis wimaniSchaub, 1937, but slightly larger.
The genusMegalovisof the Villafranchian are thought by many to be similar to the ancestral form ofOvibos, if not directly ancestral (Crégut-Bonnoure, 1984). Actually, Teilhard de Chardin and Piveteau (1930) had indicated the close affinity ofMegalovis piveteauiwithOvibosin the following aspects: cheek teeth without basal pillars, premolars quite hypsodont(Fig. 10A), upper molars with small enamel loops between the lobes at the lingual side (Fig.10B). Furthermore, the present authors also think the upper molars are quite elongated in crown and the upper premolars with spurs (Fig. 10B). Although the cranial and cornual features are almost completely unknown, the close relationship betweenMegalovisand ovibovines is verified. On the other hand, the metapodials ofMegalovisare also quite stout (Table 4), which resembles those of the recentOvibos.

Fig. 10 Maxilla of Megalovis piveteaui (MNHN-NIH 150) from Xiashagou of Nihewan A. buccal view; B. occlusal view
Because of the poor fossil records, the knowledge about the features ofMegalovisin China is very fragmented. The speciesMegalovis guangxiensisHan, 1987 was established based on partial jaw bones and isolated teeth from the LiuchengGigantopithecusCave in Guangxi. The striking differences between the species and otherMegalovisspecies from the Palearctic Region lie in its extremely larger size and developed basal pillars as well as the dental cementum; the m3 basal length ofM. guangxiensiscan be as large as 50 mm (Han,1987), which almost represents the largest bovid tooth ever discovered in China. The present authors suggest that the fossils once attributed to the speciesM. guangxiensisshould be reconsidered, not only because they are too different from the normalMegalovisspecies both in size and form, but also for the far distance between them.
The teeth ofMegaloviscan be easily distinguished from those ofSpirocerusandOvisshantungensisin the same fauna by the characters as mentioned above; in size, they are prominently larger than those of the two latter taxa.
Concerning the other two contemporaneous local genera of Ovibovine,BoopsisandBudorcas, they are also poorly represented in the fossil record. It’s difficult to go any further to make an intensive comparison among them.
Moreover, the SSMZ metacarpal bone has the most similar dimensions with those ofM.piveteauifrom XSG in Nihewan (Teilhard de Chardin and Piveteau, 1930) except its slightly larger size, but with almost the same stoutness index (Table 4).
5 Discussions
5.1 Distinctions of metapodial bones
The bovid metacarpal bones unearthed at SSMZ site are quite diversified, and most of them are easy to be identified according to their size and proportion, whereas some of them are very similar to each other both in size and stoutness. Therefore, an index should be employed,that’s the robusticity index or stoutness index. Concerning the robusticity index, there exist two calculation methods: one is the relative dimensions of mediolateral diaphysis width and maximum length (SD/GL) ×100) (Maniakas and Kostopoulos, 2017), the other is the distal width/total length (Sher, 1997). Because it’s easier and more practical to measure the distal width, this paper follows Sher’s (1997) method to calculate the robusticity index, i.e. (distal width/greatest length) ×100. Based on the morphological observations and robusticity index calculations, the metacarpal bones of bovids from SSMZ site are quite clearly distinguished(Table 4; Fig. 11).

Fig. 11 Length vs distal width of metacarpal bones of diverse bovids The data un-included in Table 4 are from Colbert and Hooijer, 1953; Scott, 1985
The metacarpal bone ofSpirocerus wongiis the shortest and stoutest one, which is one of the crucial characters of the ovibovine taxa. In regard to the metapodial characters, it seems thatSpirocerushas a closer affinity with the ovibovines rather than with the antilopines;Whereas almost all of the current publications place the genusSpiroceruswithin the subfamily Antilopinae or even within the tribe Antilopini together withGazellaand other gazelles.
The metacarpals of the allied taxa,GazellospiratorticornisandPontoceros ambiguous,from Russia (Vislobokova et al., 2020) are much longer than those of the species ofSpirocerus.
Megalovis piveteauiandBison palaeosinensishave quite similar metacarpals both in size and proportion, the only difference is that the formers’ is slightly smaller. Moreover,the Nihewan specimens ofM. piveteauishare the similar size and form withM. latifronsof European (Viret, 1954; Radulesco and Samson, 1962) (see Table 4).Megaloviswas referred to Ovibovini, maybe its metacarpal is the largest, but less stout, among the ovibovine taxa.M.piveteauishould be larger thanSpirocerus wongiin body mass.
The metacarpal bone ofGazella sinensisis the most slender one among the bovid metacarpals in SSMZ fauna. The metacarpal specimens for the Pleistocene gazelle species are very rare in China, there is only one example up to date, i.e. the metacarpal of a Late PleistoceneProcapra przewalskiifrom Sjara-osso-gol, whose length is only 142 mm, but the counterpart of the extantProcapra gutturosais 180 mm (Boule et al., 1928). Therefore, it can be estimated that the body mass ofG. sinensisis similar with that of aP. gutturosa, and larger thanP. przewalskiiand the Late Miocene speciesGazellacf.G. lydekkeriwhose metacarpal’s length is only 141 mm (Zhang and Yang, 2016).
The slenderness and total length of the metacarpal ofOvis shantungensisis very similar with that of a medium-sized deer, which is much longer and slender than those of the other Caprinae taxa, such asPseudoisandCapricornisas well asCapra.
For the extant ovibovines, the metapodials are exclusively short and robust, but for the extinct taxa, their metapodials are completely unknown exceptMegalovis, which is much longer than the counterparts of the living ovibovines. In quite a number of genetic studies,NaemorhedusandCapricorniswere grouped withOvibos(Hassanin et al., 2012), which makes the forms of the subtribe Ovibovina much diversified.
5.2 The bovid assemblage of the SSMZ fauna and its chronological significance
In China, Nihewan Basin and Yushe Basin are the most important fossiliferous spots for Late Cenozoic bovids, but the latter’s bovid fauna is much more diversified (Teilhard de Chardin and Trassaert, 1938) than the former’s. In Yushe Basin of Shanxi Province, the late Cenozoic deposits were subdivided into three biostratigraphical zones from bottom to top:Zone I, Zone II and Zone III (Licent and Trassaert, 1935; Teilhard de Chardin and Trassaert,1938; Tedford et al., 1991); the Zone III shares almost all of its faunal members with the Nihewan fauna. In regard to the bovid taxa, only the advanced and relict ones of each group of Yushe Basin also appear in the Nihewan Basin; the relict elements includeGazella,AntilospiraandMegalovis; the newly appeared taxa includeSpirocerus,OvisandBison. The recently discovered fauna in Jinyuan Cave in Dalian of Liaoning Province bears a very similar bovid assemblage (Jin et al., 2021) with the Nihewan fauna.
In the Nihewan Basin, exceptGazellacf.subgutturosa, all the bovid taxa of the XSG fauna also occur in the SSMZ fauna. Furthermore, the mandible of Antilope gen. indet. of the XSG fauna (Teilhard de Chardin and Piveteau, 1930:68, fig. 25) was reidentified asHydropotesby Tong et al. (2021).
In the SSMZ fauna,Gazella sinensisis the richest in fossil specimens, most of which are horn-cores and mandibles.Spirocerus wongiare better represented than the previous reports,especially its metapodials represent the first discovery. ForMegalovis piveteaui, radius is the first time to be reported. Except a metacarpal bone, no other new anatomic elements were discovered forOvis shantungensis.
Among the spiral horned antelopes,Spirocerus wongishares the same chronology withAntilospiraof the local area andGazellospiraspecies of central Asia and Europe, which is the Late Pliocene to the Early Pleistocene and no younger than 1.6 Ma (Hermier et al., 2020).
TheOvis shantungensisin Nihewan Basin used to be regarded as the earliest record ofOvisuntil the recent reports of the Early PlioceneOvissp. from Udunga in the Trans-Baikal region of Russian Siberia (Vislobokova et al., 1995; Vislobokova, 2008). The horn-cores ofO. shantungensisare less stout than the laterO. ammon(Bohlin, 1938), and it’s almost certain thatO. shantungensisindicates an Early Pleistocene age.
Moreover, the Nihewan fauna and other Early Pleistocene faunas in northern China correspond with a stage of decline of the spiral-horned antelopes and the rise of the hollowedhorn caprines (with developed horn-core sinuses or cornual diverticulum) and the appearing of large bovids (Bovini) and the ovibovines (Ovibovini).
Furthermore, the bovid assemblage of the Nihewan fauna indicates an open prairie or steppe dominated environment.
6 Conclusions
The small and medium-sized bovid fossils from Shanshenmiaozui site were identified asSpirocerus wongi,Gazella sinensis,Ovis shantungensisandMegalovis piveteauirespectively.The spiral-horned antelope was referred toS. wongibased on its twisted and with only one anterior keel of the horn-core;S. wongihas short but stout metapodials as does the speciesS.kiakhtensis; the cranial and horn-core characters ofSpirocerussupport a closer relationship with antilopines, while the metapodials share more similar traits with ovibovines. The speciesG. sinensisis typical for its lower p4 which is different from that ofG. subgutturosaand earlierGazellaspecies in its exclusively closed anterior valley, and it is different from theProcapraspecies in its exclusively open posterior valley, no matter how narrow it is. AlthoughO.shantungensisis poorly represented in the SSMZ fauna, the metacarpal bone is the first time to be recognized. The present study shows thatO. shantungensishas much longer metapodial bones than other caprine taxa do.M. piveteauiis also poorly represented in the fossil record,but it has very typical lower p4 and robust metacarpal bone; both characters support a closer affinity ofMegalovisto ovibovines. Among the postcranial bones, the metapodials,especially the metacarpal bones are crucial not only for taxonomic identification, but also for phylogenetic and paleoecological reconstructions. The Shanshenmiaozui fauna shares 92%of its elements with that of the classical Nihewan fauna, which means they should have the similar geologic age, i.e. 2.2-1.7 Ma BP.
AcknowledgementsThe authors wish to express their thanks to the following people and organizations for their help: Wei Q, Han F, Sun B Y, Lü D, Sun J J, Xu Z J, Qiu Z W, Wang Q Y, Sun B H, Hu N, Liu X T and Yin C for participating the excavations; Xie F of HPICR, Li H Q of NNNRM and Hou W Y of NM for helps during excavations; Zheng M of TNHM, Argot C of MNHN, Li W J of NWIPB and Zhu X C of IOZ for providing access to collections; Qiu Z X, Zhou Z H, Vislobokova I A, van der Made J, Titov V, Salvagno L, Davis S J M, Jiang Z G, Zhang Z Q, Bai W P, Shi Q Q, Wang Y R and Wang J for sharing bibliographies and/or for fruitful discussions. Hou Y M for CT scanning. Thanks are also due to the two reviewers,Kostopoulos D S and Dong W, for their insightful questions and comments. The Editorin-Chief and the editors contributed a lot to the improvement of the article. This work was supported by the following grants: the National Natural Science Foundation of China (Grant Nos. 42172021, 41572003), the Strategic Priority Research Program of Chinese Academy of Sciences (Grant No. XDB26000000) and the National Key R & D Program of China (Grant No. 2020YFC1521500).
