
多数据库研究CRYAB在宫颈癌中的表达及相关机制
2022-07-19 02:11黄宏宝吴小勤卢东林覃莉韦敬锡罗小琼黄秋艳陈发钦
右江民族医学院学报 2022年3期
黄宏宝,吴小勤,卢东林,覃莉,韦敬锡,罗小琼,黄秋艳,陈发钦
(1. 右江民族医学院,广西 百色 533000;2. 右江民族医学院附属医院生殖医学中心,广西 百色 533000;3. 右江民族医学院附属医院临床病理诊断与研究中心,广西 百色 533000;4. 右江民族医学院附属医院妇科,广西 百色 533000)
宫颈癌是妇科最常见的恶性肿瘤之一,对女性的身心健康构成严重的威胁。国际癌症硏究中心(IARC)最新数据显示,宫颈癌在全球女性肿瘤的发病率和死亡率中仍然排名第四[1]。宫颈癌的病因目前不十分明确,公认的高危风险是高危型人类乳头状瘤病毒(HPV)感染,IARC将13种高危HPV亚型认定为Ⅰ类致癌物。近年来,宫颈癌筛查的广泛普及使宫颈癌的早期诊断率得到明显提升,但是中晚期或者复发的患者死亡率依然居高不下。以分子机制研究为基础的靶向治疗取得一定的进步,目前已有应用于临床的靶向药物,前景可观,但同样显现了其局限性,寻找高效的分子靶点对宫颈癌的研究意义重大。αB-晶状体蛋白(CRYAB)是一种蛋白质编码基因,染色体定位于11q23.1,长度约15 177 B,含6个外显子,和其他小分子热休克蛋白一样,它有一个保守的中心结构域,而位于两侧的N端结构域和C端结构域是高度可变的非保守序列,这些序列对于分子伴侣功能至关重要[2]。CRYAB分子伴侣功能可以防止因环境刺激导致的靶蛋白展开和聚集,并将易聚集的蛋白保存在其可溶性寡聚体结构中,避免蛋白质功能的丧失和有毒中间产物的生成[3]。CRYAB最初在人眼的晶状体中被发现,最新的Bgee Gene显示,它在心脏和243 种其他组织中表达。研究发现,CRYAB在鼻咽癌、膀胱癌、前列腺癌中表达下调[4-6],而在乳腺癌、骨肉瘤等肿瘤中高表达[7-8]。目前CRYAB与宫颈癌的关系尚不明确,本研究利用生物信息学系统分析CRYAB在宫颈癌中的表达及相关机制,旨在为宫颈癌的分子研究提供新的思路。
1 材料与方法
1.1 CRYAB在宫颈癌及泛癌中的表达 运用UALCAN网页工具(http://ualcan.path.uab.edu/)分析TCGA数据库宫颈癌组织样本中CRYAB的表达情况。UALCAN是美国阿拉巴马大学伯明翰分校的癌症数据分析门户网站,该网站是一个内容丰富且对用户友好的网络资源,用于分析各种癌症分开数据库(TCGA、MET500、CPTAC 和 CBTTC)中的组学数据[9]。利用临床生信之家(https://www.aclbi.com)网页工具分析GEO数据库宫颈癌组织样本中CRYAB的表达情况。该平台是国内学者搭建,原始数据来源自TCGA和GEO数据库,所有分析数据均由Rv4.0.3分析完成,用户可以在不使用R语言的情况下完成对数据的全方位分析,GEO数据库分析更是其亮点。通过网站上GEO模块中的基因表达比较选项来探索GEO数据库GSE46857组织样本中CRYAB的表达情况[10]。利用仙桃学术网站(https://www.xiantao.love)分析TCGA和GTEx数据库对比宫颈癌样本中CRYAB的表达情况。仙桃学术网的生信工具同样为用户提供了便捷的生信分析方式,将GTEx的数据纳入分析,提高结果的可靠性。
1.2 CRYAB的表达与宫颈癌生存期及临床病理特征的关系分析 通过GEPIA数据库(http://gepia.cancer-pku.cn/)分析CRYAB与宫颈癌患者生存期的关系;该网站基于TCGA和GTEx数据为用户提供快速且可定制的分析功能[11]。UALCAN用来分析CRYAB与宫颈癌患者肿瘤分期、人种、体重、年龄、组织学类型和淋巴结转移的相关性。
1.3 免疫浸润分析 通过TIMER 2.0(http://timer.cistrome.org/)分析在宫颈癌组织中CRYAB表达量与免疫细胞浸润的相关性;TIMER 的最大特色是免费分析不同种类癌症的免疫浸润。2.0版本的网络服务器通过多种计算方法评估免疫细胞的浸润程度,而用户只需简单的操作即可得到高质量的图形,以此来全方位的探索肿瘤免疫学、基因组和临床特征[12-14]。
1.4 共表达基因分析 LinkedOmics数据源来自TCGA项目的 32 种癌症类型和总共11 158名患者的多组学数据和临床数据[15]。通过LinkedOmics(http://www.linkedomics.org)网站下载并分析CRYAB共表达的基因,将高表达前50和低表达前50的基因制作成热图。STRING (https://cn.string-db.org/)数据库从多个来源收集证据,目的是整合蛋白质之间所有已知和预测的关联,包括物理相互作用和功能关联[16]。利用STRING数据库制作蛋白互作图,寻找关键基因。
1.5 共表达基因富集分析 从LinkedOmics网站下载共表达基因信息,筛选出高表达前200和低表达前200的基因制作成列表,再用Metascape网站(https://metascape.org/)进行下一步研究,该网站可以为用户提供全面的基因列表注释和分析资源[17],利用此功能对共表达基因进行GO和KEGG通过富集分析,寻找可能的作用机制。
2 结果
2.1 CRYAB基因在宫颈癌和癌旁组织或正常宫颈组织的表达比较 在TCGA中,CRYAB基因在宫颈癌组织中的表达水平低于癌旁组织,见图1A;在GEO数据库GSE46857数据集中,CRYAB基因在宫颈癌组织中的表达水平低于癌旁组织,见图1B;CRYAB在27种癌症中存在差异表达,差异有统计学意义,见图1C。

A:CRYAB在TCGA宫颈癌及癌旁样本中的表达情况;B:CRYAB在GEO宫颈癌及癌旁样本中的表达情况;C:CRYAB在TCGA和GTEx泛癌配对样本中的表达情况。
2.2 CRYAB的表达与宫颈癌生存期及临床病理特征的影响 GEPIA数据库分析结果显示,CRYAB的表达与宫颈癌患者的总体生存期无相关性(P=0.58),见图2A,与患者的无病生存期无相关性(P=0.23),见图2B。UALCAN数据库分析表明,从肿瘤分期来看,Ⅱ期、Ⅲ期和Ⅳ期患者的CRYAB表达低于癌旁组织,Ⅰ期患者CRYAB表达量与癌旁组织无差异,4个期别间的CRYAB表达量无差异,见图3A。从人种来看,白种人、非裔美国人和亚洲人宫颈癌患者CRYAB表达量低于癌旁组织,各人种之间的CRYAB表达无差异,见图3B。从体重来看,正常体重、超重、肥胖和过度肥胖患者CRYAB的表达量低于癌旁组织,但不同体重组之间表达量无差异,见图3C。从年龄层次来看,21~40岁组和61~80岁组患者CRYAB的表达量较癌旁组织明显偏低;41~60岁组较21~40岁组患者CRYAB的表达量偏高,而81~100岁患者的CRYAB表达量高于41~60岁组;其余各组的表达量无差异,见图3D。从组织学类型看,宫颈鳞状细胞癌、普通型宫颈管腺癌和宫颈内膜样腺癌组织中CRYAB的表达量低于癌旁组织;宫颈鳞状细胞癌组织中CRYAB的表达量明显高于宫颈腺鳞癌和普通型宫颈管腺癌;其余各组间表达无差异,见图3E。从淋巴转移的情况来看,N0期(无局部淋巴结转移)和N1期(有1~3枚腋窝淋巴结转移)组织中CRYAB表达量低于癌旁组织,但N0和N1两组间表达量无差异,见图3F。

A:CRYAB表达量与宫颈癌患者总体生存期相关性;B:CRYAB表达量与宫颈癌患者无病生存期相关性。

A:CRYAB与宫颈癌肿瘤分期的关系;B:CRYAB与宫颈癌患者人种的关系;C:CRYAB与宫颈癌患者体重的关系;D:CRYAB与宫颈癌患者年龄的关系;E:CRYAB与宫颈癌组织学类型的关系;F:CRYAB与宫颈癌患者淋巴结转移的关系。
2.3 CRYAB与宫颈癌免疫细胞浸润水平的相关性 TIMER 2.0分析显示,CRYAB在宫颈癌组织中与肿瘤细胞纯度无相关性,与造血干细胞、肿瘤相关成纤维细胞和内皮细胞呈显著正相关(P<0.01),与单核巨噬细胞、NK细胞、CD8+ T细胞和B细胞呈负相关(P<0.05)。如图4所示。

图4 CRYAB表达与宫颈癌免疫细胞浸润水平相关性
2.4 宫颈癌组织中的CRYAB共表达基因分析 利用LinkedOmics网站,筛选出与CRYAB具有正或负相关性的前50个共表达基因,并以热图的形式展示,正相关的基因按P值升序排列,前5个为:NDRG4、IVL、KRT75、SBSN和FABP12,见图5A;负相关的基因按P值升序排列,前5个为:KIAA0649、TMPRSS3、GOLM1、ALDH3B1和RHOBTB2,见图5B。STRING蛋白互作分析显示,SPRR1B、IVL、SPRR1A、SPRR3和SPRR2A相关联,CRYAB与HSPB1和HSPB2相关联,见图5C。
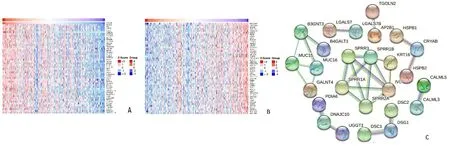
A:宫颈癌中与CRYAB表达正相关的基因;B:宫颈癌中与CRYAB表达负相关的基因;C:共表达基因互作关系。
2.5 CRYAB及共表达基因GO注释和KEGG通路富集分析 为了预测与CRYAB共表达基因的功能,下载LinkedOmics网站上的共表达基因,选取上调和下调前200个基因合并成列表,利用Metascape网站进行GO注释和KEGG通路富集。结果显示,共表达基因参与表皮发育、包膜角化、多肽交叉连接、钙离子结合和中间纤维细胞骨架组织等,见图6A,通路富集在金黄色葡萄球菌感染、黏蛋白O-聚糖生物合成、核苷酸糖的生物合成、IL-17信号通路和癌症的通路等,见图6B。

A:共表达基因GO注释;B:共表达基因KEGG通路富集。
3 讨论
宫颈癌是一种女性常见的恶性肿瘤,好发于子宫颈鳞状、柱状上皮连接处的细胞。“高致癌性”人乳头瘤病毒(HPV)感染[18]和主要遗传因素[19](包括基因和染色体的改变、抑癌因子和激活因子水平的改变)是宫颈癌发生发展的关键。中国2020年宫颈癌的新发病例为109 741例,死亡病例为59 060例。与2015年相比,新发病率基本持平,但死亡率却明显增加(约从3.4万上升至5.9万)[20]。寻找宫颈癌筛查的生物标志物和治疗靶点意义重大。随着政府宫颈癌筛查项目和HPV疫苗的不断推广,宫颈癌在一定程度上是可以预防的。但在中国,因地域及经济和文化的差异仍然较大,一些地区女性接种HPV疫苗或意愿仍然不高[21],某些地区女性宫颈癌健康素养较低[22],提高老年女性、文化程度低、农村妇女等人群的宫颈癌预防知识仍然任务艰巨。哺乳动物的晶状体蛋白可分为α、β和γ家族;作为分子伴侣,尽管它们不会使蛋白质再生并释放它们作为真正的伴侣。但是,它们结合错误折叠的蛋白质以防止蛋白质聚集,除此之外,CRYAB还有另外两个特别的功能,自身激酶活性和参与细胞骨架的稳定[23]。CRYAB的这些功能可能与肿瘤的调控相关。
本研究显示,CRYAB在宫颈癌中呈低表达,这与前人在鼻咽癌、膀胱癌和前列腺癌中的研究一致,CRYAB在这些肿瘤中可能是一种抑癌基因,而在乳腺癌、骨肉瘤等肿瘤中可能是促癌作用。这显示了调控因子功能的多样性和复杂性。CRYAB表达与肿瘤分期、人种、年龄、体重、组织学类型和淋巴转移存在一定的相关性,但结果表明,各肿瘤分期组间、不同人种组间、不同体重组间和淋巴转移数目组间CRYAB的表达量无差异。从年龄来看,年龄更大的患者,CRYAB的表达量有升高的趋势,在不同组织类型的宫颈癌中,CRYAB的表达量存在一定差异,但无明显规律。本次研究结果还显示,CRYAB的差异表达诱导肿瘤微环境中免疫细胞的改变,正相关的细胞群中差异性很大,而负相关的细胞群中差异性相对较小,这可能提示,宫颈癌中低表达的CRYAB导致造血干细胞、肿瘤相关成纤维细胞和内皮细胞大降减少,单核巨噬细胞、NK细胞、CD8+T细胞和B细胞增加,两者共同作用,促进癌症的发生发展。共表达基因GO注释发现,它们参与表皮发育、包膜角化、多肽交叉连接、钙离子结合和中间纤维细胞骨架组织等细胞功能,可以推测,CRYAB可能通过上述方式调节细胞周期,从而参与肿瘤的发生和进展。蛋白互作发现的重点分子SPRR1B和IVL已有报道与宫颈癌的调控相关[24-25],这更提供了CRYAB作为相关因子参与宫颈癌调控的依据。KEGG通路分析显示共表达基因富集在金黄色葡萄球菌感染、黏蛋白O-聚糖生物合成、核苷酸糖的生物合成、IL-17信号通路和癌症的通路等,这为今后CRYAB在宫颈癌的通路研究提供了理论基础。
综合上述,CRYAB在宫颈癌中低表达,与宫颈癌的发生发展存在一定的关系,有望成为宫颈癌的生物标志物和潜在的治疗靶点。
猜你喜欢
中国现代医生(2022年19期)2022-11-04
中国医学物理学杂志(2022年9期)2022-10-09
中国现代医生(2022年19期)2022-08-25
海峡姐妹(2020年2期)2020-03-03
保健与生活(2019年3期)2019-08-01
财经(2017年2期)2017-03-10
财经(2016年15期)2016-06-03
财经(2016年3期)2016-03-07
财经(2016年6期)2016-02-24
中国民族民间医药·下半月(2014年4期)2014-09-26
