
肾小球系膜细胞中TGF-β受体1的反式激活调控了CTGF的增量表达①
2022-07-19 13:32王耀坤刘双春
黑龙江医药科学 2022年3期
王耀坤, 刘双春
(1.佳木斯大学医学部,黑龙江 佳木斯 154007;2. 台州市立医院输血科,浙江 台州 318000)
肾小球毛细血管压力升高 (glomerular capillary pressure, Pgc) 是不同病因诱导的慢性肾脏疾病进展中主要致病决定因素。利用机械应激的实验方法,可以体外模拟肾小球系膜细胞(glomerular mesangial cell, GMC)高Pgc的传递,在结构方面恰好为肾小球毛细血管簇提供了必要的支持[1]。在体外,循环应用真空与可变形孔板模型拉伸效应。在基质上生长的MC经循环拉伸/松弛后增加了基质蛋白的合成[2],此方法恰恰为MC中机械应激诱导信号的研究提供了一个模型系统。另外,基质蛋白合成和肾小球纤维化的两个关键介质是促纤维化细胞因子转化生长因子β1 (Transforming factor β1, TGF-β1)和结缔组织生长因子(connective tissue growth factor, CTGF)。CTGF至少介导了TGF-β1的部分促纤维化作用,但它也对基质上调有独立和叠加的作用[3]。CTGF的上调与肾纤维化的严重程度密切相关,抑制其表达对MC具有明显对的保护作用[4]。机械拉伸是否能够上调CTGF的表达,其分子机制如何目前还不清楚。本研究利用原代培养的GMC作为研究对象,利用机械拉伸的实验方法分析 CTGF的表达变化及TβR1在此过程中的作用。研究结果可以为临床慢性肾病的有效治疗提供必要的理论和实验支持。
1 材料与方法
1.1 实验材料
SD大鼠肾小球系膜细胞(GMC),由加拿大 Joan Krepinsky教授(McMaster University)惠赠。DMEM 培养液(美国Gibco公司);胎牛血清(美国Sigma公司);Trizol(美国ThermoFisher公司); 抗CTGF抗体(mouse,美国Santa Cruz公司);抗Tubulin抗体(rabbit, 美国Cell Signaling公司);抗P-Smad3 S423/S425 抗体((rabbit,美国Cell Signaling公司);TβR1阻断剂SB431542 (5 mM, 30 minutes)(美国Cell Signaling公司),细胞培养中需要的其他耗材均购自Corning 公司。
1.2 细胞培养
SD大鼠,雌性,乙醚麻醉后用事先冷藏的生理盐水反复灌洗肾脏,观察肾脏颜色变白后按照常规细胞原代培养方法进行[5,6]。6~15代的GMC饥饿24h后进行机械拉伸处理。
1.3 机械应激(Stretch)实验操作方法
6~15代GMC细胞,制备单细胞悬液后铺在一种特殊的6孔板上,底部有弹性,涂有牛I型胶原蛋白(购置于Flexcell International公司,美国)。当细胞融合度达到90%~95%时饥饿24h。机械应激实验前先加入TβR1阻断剂SB431542 (5 mM, 30 minutes),然后将铺满细胞的6孔板置于一种机器控制的真空中(Flexercell 4000, Flexcell International Corp.),连续循环拉伸/放松,每循环0.5秒的拉伸和0.5秒的放松,总共60个循环/min。
1.4 Wester Blot方法检测细胞中CTGF和P-Smad3蛋白表达
取对数生长期的GMC,调整其浓度为2.5×105/mL,每孔1mL,接种于直径6cm 细胞培养皿中,置入37℃、5% CO2培养箱培养,细胞融合度达到80%~85%时饥饿24h,分为正常对照组、Stretch 1h组、Stretch 3h组、Stretch 6h组,固定时间点加入细胞裂解液,冰上裂解30min,14000r /min,4℃离心10min收集上清。采用SDS-PAGE检测目的蛋白的表达,各泳道蛋白上样量为50μg。再将凝胶上的蛋白转印到0.45μm硝酸纤维素膜( NC膜) 上,NC 膜用脱脂奶粉进行封闭,然后与一抗孵育4℃过夜,TBST 洗涤后再与过氧化物酶标记的二抗室温孵育2h,TBST再次洗涤,电化学发光( electrochemiluminescence,ECL) 检测免疫印迹信号,发光成像仪拍照记录,Image J图像分析软件进行条带分析。
1.5 统计学方法
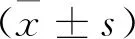
2 实验结果
2.1 机械应激增量调节了CTGF的表达
GMC细胞通过Stretch处理1h、3h和6h后CTGF的表达明显增加,并与作用时间呈正相关(P<0.05),说明外源性机械应激可以明显激活GMC细胞CTGF的表达。见图1。

图1 机械应激上调了GMC细胞CTGF的表达与对照组相比,*P<0.05。
2.2 机械应激增量调节了P-Smad3的表达
GMC细胞通过Stretch处理30S、1h和3h后P-Smad3的表达明显增加,并与作用时间呈正相关(P<0.05),说明外源性机械应激可以明显激活GMC细胞P-Smad3的表达。见图2。

图2 机械应激实验上调了GMC细胞P-Smad3的表达与对照组相比,*P<0.05。
2.3 TβR1阻断剂低调节了机械应激诱导的CTGF增量表达
GMC细胞通过Stretch处理1h后CTGF的表达明显增加,TβR1阻断剂则明显降低了CTGF的表达(P<0.001),说明外源性机械应激激活GMC细胞CTGF的表达依赖TβR1。见图3。

图3 TβR1阻断剂降低了机械应激诱导的GMC细胞CTGF增量表达与对照组相比,P<0.001。
2.4 TβR1阻断剂低调节了机械应激诱导的P-Smad3增量表达
GMC细胞通过Stretch处理1h后P-Smad3的表达明显增加,TβR1阻断剂则明显降低了P-Smad3的表达(P<0.01),说明外源性机械应激激活GMC细胞P-Smad3的表达依赖TβR1。见图4。

图4 TβR1阻断剂降低了机械应激诱导的GMC细胞P-Smad3增量表达与对照组相比, P<0.01。
3 讨论
更深入地了解调节基质生成的分子机制,对于发掘治疗慢性肾脏疾病的新的潜在治疗靶点至关重要。我们目前的研究提出了几个新发现。首先我们确定了机械性拉伸实验以一种配体独立反式激活方式激活TβR1发挥促纤维化作用。也就是说在没有TGF-β1存在的情况下,GMC处于一种机械应激状态下TβR1可以被激活,发挥诱导基质生成促纤维化的作用。事实上,大量文献已经证实,虽然Stretch可以诱导TGF-β1分泌水平增加,但均发生与Stretch后48~72h[7]。而本实验中发现Stretch 1h后 TβR1已经活化。GMC中TGF-β1的分泌比信号通路的激活和CTGF的上调晚得多。
体外研究证实,CTGF诱导的GMC改变,主要涉及到了肾脏病变程度的进展[8]。CTGF的作用机制主要涉及到增量调节了纤维连接蛋白和纤溶酶原激活物抑制因子的合成,结果引起了GMC外基质产生的增加和积聚。TGF-β1与CTGF有着及其相似的功能。但是CTGF的siRNA或者RNAi均能抑制TGF-β1的这种作用。慢性肾脏疾病发生发展过程中,无论是基质扩张,还是GMC肥大,病理改变均发生于系膜区,由此说明TGF-β1是肾脏疾病病理损伤中的决定性因子。在此过程中CTGF则是促进TGF-β1作用发挥并增强其效应的重要调节子[9]。相关文献证明尽管机械应激导致GMC中CTGF上调,但其上调的机制尚不清楚[10]。本实验结果发现,TβR1在机械应激诱导的Smad3的磷酸化和CTGF表达上调中是绝对需要的。有趣的是,Smad3的激活是由配体独立的TβR1激活介导的。相关文献证明,Smad3也参与了内半月板细胞机械应激诱导的CTGF表达[11]。综上所述,我们发现TGF-β1是GMC机械应激诱导CTGF上调的重要诱导子,Smad3作为TGF-β1下游蛋白也是诱导CTGF增量表达所必须的。我们的研究为评估TGF-β1抑制剂作为一种慢性肾脏疾病的新型抗纤维化策略提供了强有力的基础。
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中国现代医生(2022年21期)2022-08-22
传染病信息(2022年3期)2022-07-15
天津医科大学学报(2021年4期)2021-08-21
临床肾脏病杂志(2020年6期)2020-07-17
陕西农业科学(2020年4期)2020-07-06
医学研究杂志(2015年9期)2015-07-01
医学研究杂志(2015年12期)2015-06-10
天津体育学院学报(2015年6期)2015-06-01
老友(2010年9期)2010-09-16
