
茄科植物黄萎病研究进展
2022-07-29 07:03陈恩发叶夕苗刘聪聪范士杰
江苏农业科学 2022年14期
周 帅, 卢 扬, 陈恩发, 叶夕苗, 阳 腾, 刘聪聪, 范士杰
(贵州省农业科学院生物技术研究所,贵州贵阳 550006)
茄科(Solanaceae Pers)共包含有75~80个属,2 000 多个种,我国大约有16个属,70个种。其中茄子(L.)、番茄(L.)是重要的经济类蔬菜作物,马铃薯(L.)更是当今世界上最主要的非谷类粮食作物;因此,它们的安全生产对于保障蔬菜粮食安全和经济发展至关重要。由大丽轮枝菌等引起的黄萎病是茄子、番茄、马铃薯等茄科作物病害中危害较为严重的真菌性病害之一,该病害通过土传发生在作物生长关键期,危害严重,可导致番茄、茄子、辣椒减产20%~60%。
在过去的20年中,随着寄主及黄萎病病原菌基因组序列的测定和基因注释的完成,黄萎病致病基因、作物抗病基因的定位与克隆以及作用机理成为研究热点。本文从病原菌和寄主茄科作物番茄、茄子、马铃薯入手,系统地总结了近年来茄科作物黄萎病研究进展,对抗病资源筛选和抗病品种的选育进行梳理,以期为茄科作物培育高抗品种提供理论依据。
1 黄萎病病原菌
1.1 黄萎病的发生与危害
黄萎病为典型的土传兼种传病害,由于此病害在田间主要呈现为叶片黄色斑驳和萎蔫的症状,故称为黄萎病。我国黄萎病历史最早可追溯至20世纪40年代,起因于从美国引进了带黄萎病病原菌的“斯字棉 4B”棉花种子。90年代后期黄萎病在我国各作物产区大暴发,至今已超过100种作物受到黄萎病的危害(表1),包含但不限于花卉果树、油料纤维和蔬菜粮食,对蔬菜粮食中茄科作物生产造成的负面影响尤为明显。例如,全国7省(区)因马铃薯黄萎病而发生病害的种植田占调查的70.9%,最高发病率达95.8%,导致马铃薯大幅减产,由于抗黄萎病种质资源缺乏和防治不力,黄萎病的发生可致番茄减产约50%,茄子减产约40%。

表1 受黄萎病危害的部分植物
1.2 黄萎病病原研究
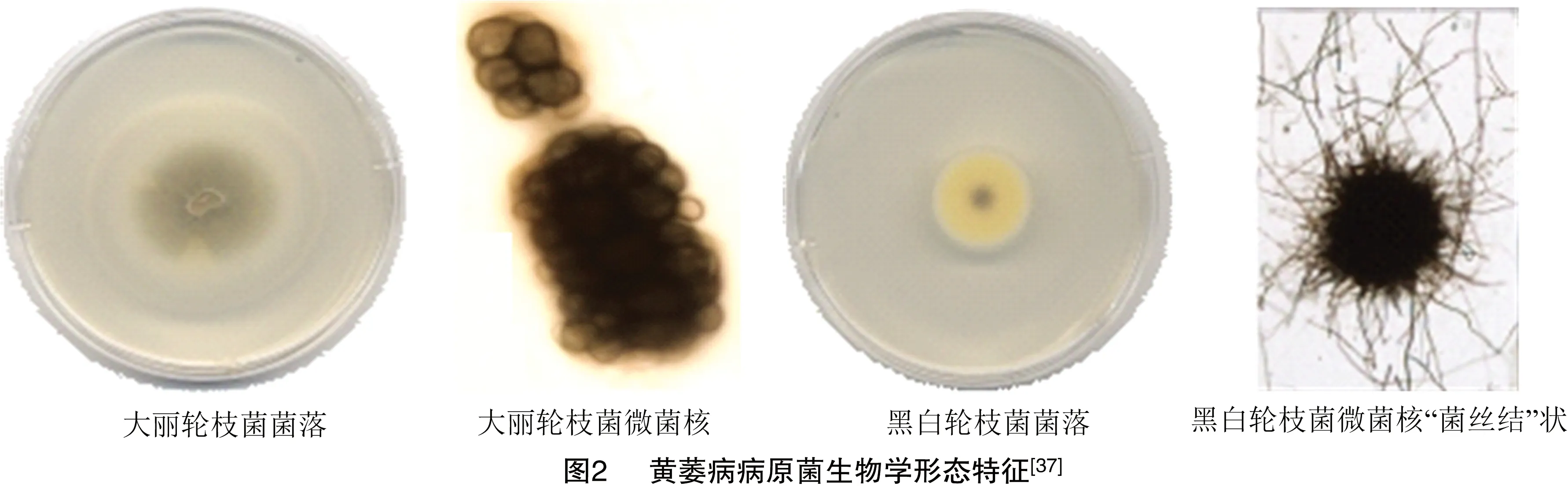
1.2.1 病原菌分类及特征 引起黄萎病病害的病原属黏菌门、丝孢纲、轮枝菌属(图1)。导致植物致病的种主要有大丽轮枝菌()、黑白轮枝菌(-)、三体轮枝菌()、云状轮枝菌()等。大丽轮枝菌()、黑白轮枝菌(-)对作物的危害较为严重。大丽轮枝菌由1条至数条菌丝相互分隔,细胞壁增厚、隆起,再向不同方向生长最终形成黑色微菌核,在30 ℃条件下大丽轮枝菌生长良好,培养初期无色,后期菌落主要呈灰褐色,菌丝呈绒毛状或干絮状。而黑白轮枝菌萌发前形态一般是“菌丝结”,在室温条件下就可生长较好,最初菌落颜色为白色,后期菌落呈黄色或橘色,菌丝呈絮状且表面光滑。大丽轮枝菌宿主比较普遍,对温度的变化和高温的适应能力也更强,黄萎病病原菌以大丽轮枝菌为主(图2)。

国内外研究者依据不同划分原则对大丽轮枝菌进行划分,结果稍有差异。国外学者从病原菌的致病力分化程度和生理形态进行鉴别,将其区分为落叶型和非落叶型2个生理小种,其中落叶型菌系的危害更普遍。根据从各省棉花上分离鉴定到的不同致病力的黄萎病原菌,国内研究者将其分为强、中、弱3个主要生理类型,强致病型以陕西径阳菌株为典型代表。
1.2.2 黄萎病病原菌基因组 随着高通量测序技术的蓬勃发展,大丽轮枝菌VDL.s17和苜蓿轮枝菌VaMS.102的基因组先后被测序并发布出来。通过比较2个轮枝菌的基因组发现,它们共编码蛋白为 8 699 个,VDL.s17特有的编码蛋白比VaMS.102要多,为1 357个,VaMS.102约1 100个。VDL.s17中某些特定基因家族拷贝数量较VaMS.102也显著增多,比如编码碳水化合物活性酶类和分泌蛋白酶类等。VDL.s17的染色体中包含区分种系的片段(lineage-specific regions,LS),在LS区域中鉴定得到2个高度同源基因:VDAG_04894.1、VDAG_04836.1,基因被报道与其寄主多样性相关。基因组分析表明黄萎病病原菌寄主复杂性程度与菌系的遗传多样性有密切关系。
1.2.3 黄萎病病原菌致病机理
1.2.3.1 病原菌侵染的生理机理 黄萎病病原菌一般以微菌核形态寄存在土壤之中,通过识别并利用寄主植物的根部分泌物促进自身萌发,萌发后的菌丝快速生长并入侵植物根系组织,致使寄主植株提前衰老死亡后,菌丝又产生大量微菌核存在于植株坏死组织中,而这些微菌核会随着寄主残留碎片回归到土壤中并进行新的侵染循环。
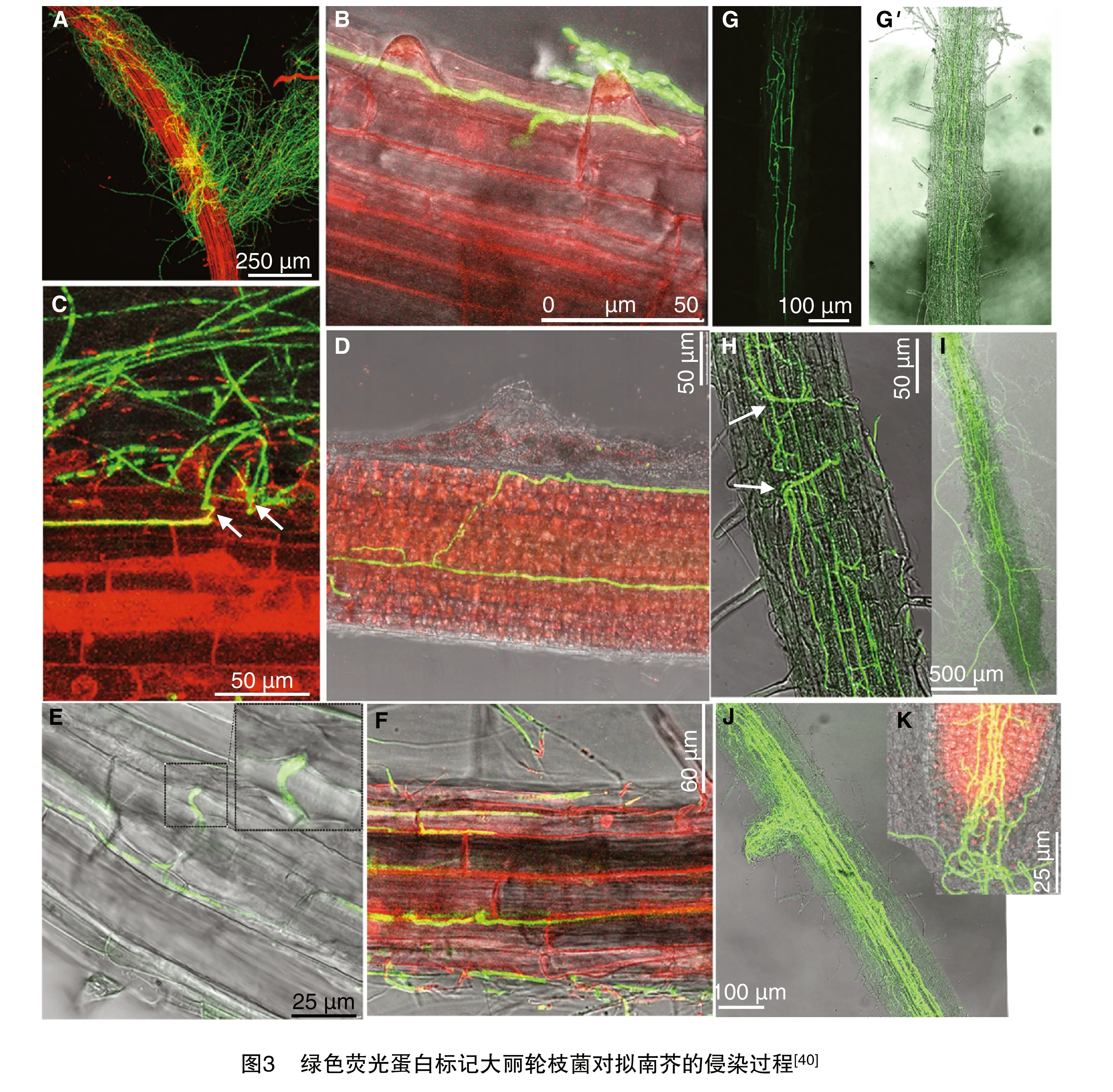
科研人员采用荧光蛋白分子标记技术对大丽轮枝菌菌株进行处理,揭示了黄萎病病原菌对拟南芥根部的侵染机理。由图3可知,当接种了大丽轮枝菌2 d后,可以观察到寄主植物根部被大量的菌丝覆盖包裹,并且部分分生孢子在根部组织中萌发,形成芽管和菌丝,沿根部组织纵向生长,一直延伸到细胞间的表皮细胞(图3-A、图3-B)。3 d后,菌丝通过形成侵染钉穿过表皮细胞迅速双向平行生长(图3-C、图3-D)。4 d后,菌丝则向维管组织扩展并迅速生长,在导管中结成一张菌丝网(图3-G)。10 d后,菌丝沿木质部导管向上延伸到地上组织,同时也沿着维管组织扩展到侧根(图 3-I、图3-J)。12 d后,菌丝向根尖区扩展导致根冠塌陷丧失吸收水分和营养的功能(图3-K)。感病材料叶片呈黄化萎蔫症状,抗病材料植株则表现正常(图4)。
1.2.3.2 病原菌致病基因的克隆 病原菌全基因组数据的发布为鉴定和分离致病相关基因、研究病原菌致病机理提供了重要的数据支撑。由于苜蓿轮枝菌的寄主范围小,对作物危害较轻。研究者多基于大丽轮枝菌测序数据对黄萎病致病基因开展鉴定研究工作。目前,已鉴定得到的致病基因主要与病原菌菌核、黑色素、降解酶、毒蛋白的形成过程密切相关,或参与调控寄主防御信号传导和病原菌效应蛋白表达等过程(表2)。
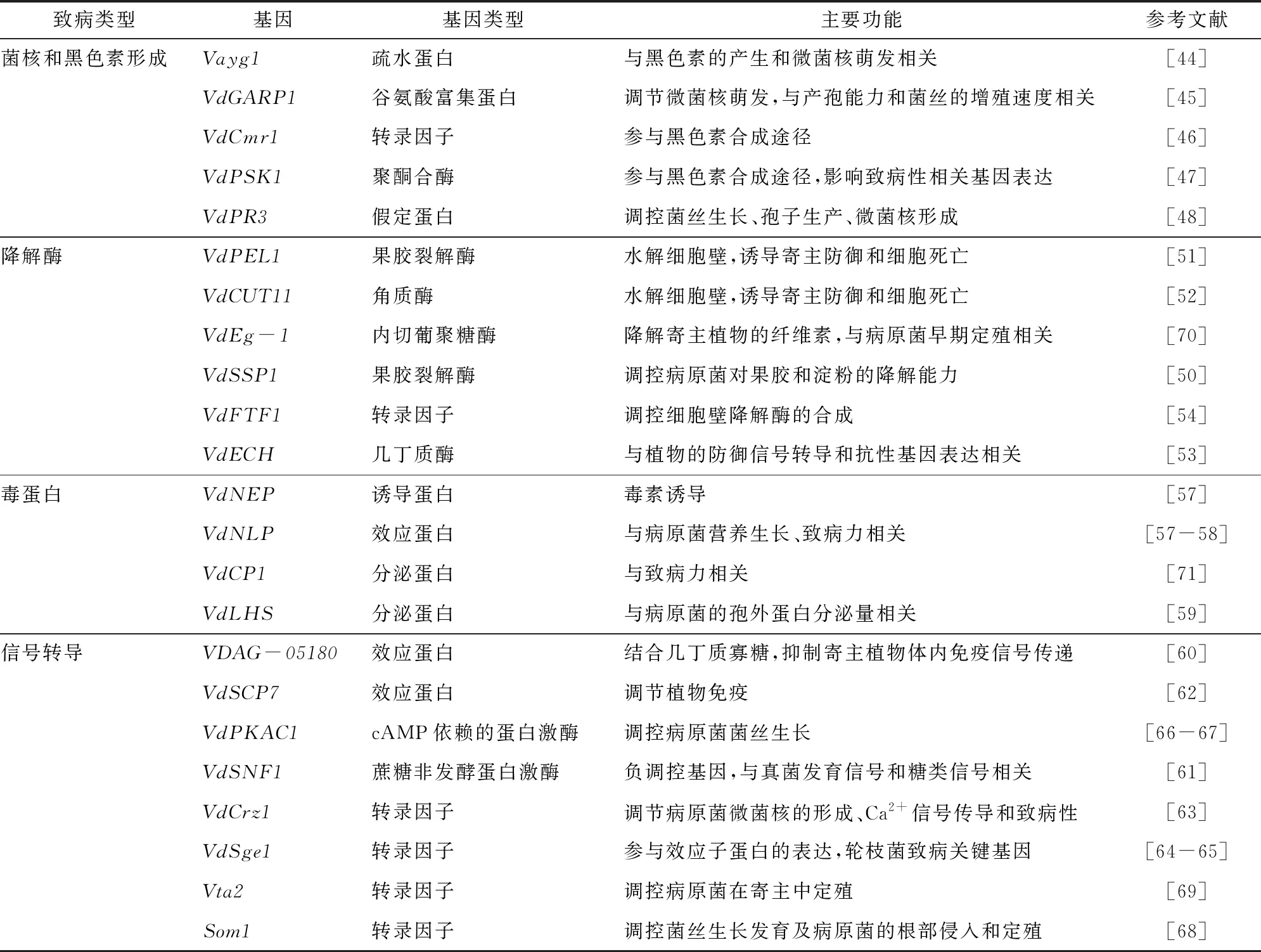
表2 大丽轮枝菌致病相关基因
病原菌一般以微菌核的形式在土壤中保留下来作为下次侵染的侵染源,微菌核产生的数量和存活率将很大程度影响黄萎病的发生水平。大丽轮枝菌侵染时须经过侵染钉附着于寄主细胞表面,这与稻瘟病病菌附着孢类似,附着孢的生长和成熟会累积大量黑色素,继而累积形成黑色素层,这是病原菌进行侵染的重要物质基础。基因是烟曲酶的同源基因,编码疏水性蛋白质,当缺失或突变会使大丽轮枝菌黑色素合成相关的基因表达下调,黑色素的积累受到负面调控,导致微菌核的萌发被抑制。编码谷氨酸富集蛋白的基因突变后会明显抑制微菌核的产生,致病菌的产孢力和增殖速度显著降低。近年来,研究并报道的与微菌核萌发和黑色素合成相关的基因还有、、。
细胞壁是植物对抗大丽轮枝菌的第1道防线,可以有效阻止其侵入到植物体内。植物细胞壁主要成分是果胶多糖多聚体。真菌侵入宿主植物后会产生并分泌细胞壁水解酶(CWDE),使寄主细胞壁中的果糖多聚体水解从而突破宿主表面的物理屏障,为继续侵染做准备。编码果胶裂解酶蛋白的是一种重要的细胞壁水解酶合成基因,缺失后植物细胞壁果胶和淀粉水解效率会降低。另一种果胶裂解酶还作为重要的致病因子参与诱导宿主免疫反应,敲除后的病原菌菌株在棉花寄主上的致病能力明显减弱。除了果胶裂解酶外,胶质酶、几丁质酶也可以水解细胞壁成分,并刺激植物的防御信号转导和抗性基因表达,相关编码蛋白的基因已被克隆。某些基因不直接合成水解酶,而是在调控细胞壁水解酶的合成上起重要作用,如。

“毒素假说”是黄萎病致病的重要学说之一。大丽轮枝菌在次级代谢中会分泌出一类被称为毒素的蛋白。毒素蛋白会破坏宿主组织细胞完整性,引起细胞膜通透性变化,使得细胞内离子外渗,最终导致植株发生萎蔫反应。是编码毒素蛋白的重要基因,通过诱导VdNEP毒素蛋白分泌使植物萎蔫坏死。病原菌营养生长情况对病原菌毒素蛋白的合成有重要影响,在基因缺失下菌株的营养生长会发生改变。主动调节毒素蛋白分泌量是病原菌发挥致病效力的另一途径,研究者对克隆到的稻瘟病LHS同源基因展开研究,发现该基因缺失后大丽轮枝菌的毒素蛋白分泌量降低70%以上,致病能力也显著降低。
在病原菌侵入致害与植物免疫防御的“博弈”过程中,病原菌会通过合成释放转录因子等产物或与靶向膜受体结合,阻断植物体内免疫信号的传递以抑制寄主防御反应。如、[]、、;或与靶基因位点结合后在效应蛋白表达()、菌丝生长发育(、、)以及病原菌侵入(、)等过程中发挥重要作用。
2 茄科植物抗黄萎病研究进展
2.1 抗病基因遗传规律和相关基因定位
从黄萎病在茄科作物上被发现以来,已有众多学者在番茄、茄子、马铃薯上相继开展黄萎病抗性遗传规律、相关抗性基因分离等研究(表3)。

表3 茄科作物黄萎病抗性基因克隆
2.1.1 番茄 雷娜等利用以父本051355(感病品种)和母本05046(抗病品种)构建的杂交群体,观察到F代单株表型均对黄萎病有抗性,从而推断母本05046中具有对黄萎病产生直接抗性作用的基因,再通过构建F群体进行表型鉴定,卡方测试显示符合单显性基因遗传规律,这证明显性单基因是决定番茄黄萎病抗性的重要基础。但是无法排除染色体组的其他位置存在抗性基因修饰因子。
基因起始于Rick等在1959年的研究报道,他们认为基因位于番茄第4条染色体。Kerr等通过筛选与基因连锁的RAPD标记,在第12条染色体上得到1个与其紧密连锁的标记,遗传距离为3.54 cM。 Diwan等通过设计RFLP基因标记进行基因制图,最终发现基因在番茄的9号染色体,与GP39标记高度连锁,负责编码细胞表面类受体激酶受体蛋白。根据基因信息比对,在类番茄茄中发现高度同源的和基因(登录号为AY262016和AY282580)。雷娜等利用1个SSR标记和3个AFLP标记围绕构建连锁遗传图谱,标记与基因的平均遗传距离为16.9 cM。应用蛋白质组学和代谢组学分析可以为抗性相关基因的定位提供便利,基于此方法,研究者发现参与防御代谢相关以及细胞壁强化相关的酶,在赋予番茄对大丽轮枝菌抗性中起关键作用。
2.1.2 茄子 茄科(Solanaceae)茄属()野生种托鲁巴姆(Swartz)对黄萎病高抗或近乎免疫,刚果茄等对黄萎病中抗。在对茄子抗黄萎病遗传规律研究中,井立军等以自交多代且抗性水平差异显著的6个株系作为亲本,轮配法配制15个组合,分析后代群体接种大丽轮枝菌后的抗病指数,显示抗性受2对及以上显性基因调控,且应当存在上位性效应。
根据NCBI数据库同源序列比对获得刚果茄的和(登录号分别为AY615303、AY590144)。Fei等基于基因序列信息采用同源序列克隆法分离得到基因,史仁玖等采用类似方法分离到了托鲁巴姆()抗黄萎病基因。除此之外,野生茄子中也含有丰富的抗病基因资源,研究者从野生茄子中克隆到、、、、。miRNA参与调控茄子对黄萎病抗性的研究也取了一些进展。Yang等用大丽轮枝菌诱导茄子后,miR482、miR93等6个 miRNA 家族基因显著差异表达。其他参与调控抗性的miRNA有miR395、miRm0002。
2.1.3 马铃薯 尽管在南美个别地区习惯以二倍体马铃薯作为栽培种,但是就全世界范围而言,马铃薯栽培种以四倍体最为常见。由于四倍体马铃薯群体复杂的分离比会产生庞大的群体统计工作,在四倍体水平上进行遗传规律解析难度巨大,因此关于黄萎病抗病遗传规律研究和报道多集中于二倍体水平上。早期研究表明马铃薯对黄萎病抗性属于数量性状。对黄萎病具有显著抗性水平差异的二倍体野生种、、等相互杂交后代群体进行表型鉴定,后代群体对黄萎病表现出连续抗性,表明抗病遗传方式为复杂的多基因遗传,Corsini等的研究结果也证实了马铃薯对黄萎病的抗性由多基因控制。也有研究表明,马铃薯对黄萎病的抗性是质量性状。Lynch等采用二倍体野生种为亲本构建F和BC群体,接种病原菌后群体抗、感病出现了 1 ∶1 的分离比,这一研究结果证明抗性由显性单基因控制。Jansky等使用2个对黄萎病均有很高抗性水平的二倍体种间杂种C545和C287作为亲本杂交材料,对分离群体接种病原菌后发现抗、感病比例为3 ∶1,基于此提出双基因控制抗性的观点。在此研究背景下,肖睿通过利用感病后代V67与抗病亲本C545回交构建BC1群体,表型鉴定感、抗病分离比为1 ∶1,进一步构建混池测序分析,在5号染色体上检测到极端单峰,也证实马铃薯对黄萎病的抗性是由显性单基因控制。
Simko等以四倍体马铃薯为试材,借助番茄基因成功定位了1个QTL位点,进一步对该位点进行序列分析,发现其可编码10个以上LRR类蛋白。与其他性状相关的基因也可能参与调控抗黄萎病性状,例如,在9号染色体上发现的QTL位点与块茎发育相关基因有显著交互作用,该位点能明显下调基因表达。本文作者通过构建BC群体结合标记开发,在验证前人研究结果可靠性的基础上,将抗性基因定位于5号染色体前端约为1.76 Mb区间内,通过测定该区间内多个NBS-LRR在接种病原菌后表达量水平,初步推测在10个NBS-LRR中可能存在真正的抗病基因。
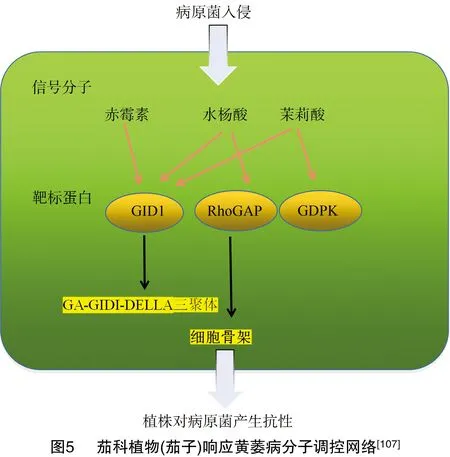
2.2 基于抗病基因解析茄科植物抗黄萎病机理
对茄科植物研究最为透彻的黄萎病抗性基因是、。介导抗性通路依赖于EDS1(enhanced disease susceptibility 1)和NDR1(non-race specific disease rResistance 1)信号途径,通过编码细胞外亮氨酸重复(leucine-rich repeat,LRR)的类受体结合蛋白,放大靶标蛋白信号来增强植物对大丽轮枝菌与黑白轮枝菌的抗性能力,也可以通过促进防卫相关的基因(PR基因)的表达提高植物的基础防御水平,并与水杨酸(SA)、茉莉酸(JA)、赤霉素(GA)等代谢通路共同组成抗病调控网络(图5)。磷脂酰肌醇特异性磷脂酶中的SIPLC6会介导对大丽轮枝菌的抗性反应,进一步研究表明,是以提高植物体内HO的浓度激活植物免疫通路的方式产生抗性。对于基因,后续研究发现该基因并不为植物提供抗黄萎病功能。
茄子的基因与番茄基因相似且编码相同的细胞表面糖蛋白,通过特异性识别病原菌产物激发植物产生一系列代谢防御反应。在野生茄子中,基因会通过激发莽草酸途径酶活性对黄萎病病菌的侵染发挥抗性作用,和则是通过调控代谢解毒物质、抗病物质等合成通路提高对大丽轮枝菌的抗性。水茄子中分离到的属于“信号转导”类型基因,编码550个氨基酸蛋白,经黄萎病病原菌处理后该基因相对表达量水平明显提高。茄子中会激活植物自身进行抗氧化防御并诱导特定抗病基因表达。
miRNA是一类不参与编码的小RNA分子,能通过调控靶基因表达参与植物生长发育和防卫反应。例如,miR93负调控靶基因参与泛素降解途径,miR482上调靶向NBS-LRR(nucleotide binding site-leucine-rich repeats)基因表达提高植物对黄萎病病原菌的防卫反应,发挥类似作用的还有miR395、miRm0002。
3 抗病品种选育进展
3.1 抗黄萎病茄科作物资源的筛选与利用
长期生产实践证实避免遭受黄萎病危害最节本增效的手段就是抗病育种。筛选并利用好高抗黄萎病材料对加快推动抗病育种工作十分重要。
尤海波对98份番茄材料进行抗病性鉴定,鉴定到抗黄萎病材料51份,耐病水平的材料21份,抗(耐)病材料占比大。王明耀等调查了117份番茄种质材料在病田的表现,发现番茄对黄萎病的抗性可分为5个类别,包含免疫、高抗、中抗、耐病和感病等不同水平,其中高抗水平材料最多,为33份。
在茄子抗黄萎病资源鉴定筛选中,大量近缘野生茄和野生茄种质资源对黄萎病抗性良好。胡建坤等采取人工接种方法,共鉴定获得茄子抗性资源11份,占鉴定材料的3.67%,近缘野生茄和野生茄种质资源筛选到的抗病材料占所有鉴定材料的3.33%,栽培种中筛选到的抗病材料占所有鉴定材料的0.33%。张鸿燕等对135份茄子种质资源进行了黄萎病抗性水平调查和评价,结果表明,80.00%抗性材料来源于近缘野生茄。因此,针对野生茄、近缘野生茄种质资源抗性基因挖掘可能是加快茄子黄萎病抗病育种进程的新途径。
在马铃薯抗黄萎病种质资源筛选过程中,将生育期纳入筛选指标可能有助于抗病品种培育。Simko等对283份材料进行抗性评价,结果显示,早熟品系对大丽轮枝菌的敏感程度比晚熟品系更高,即使调整生育期,抗性差异仍然显著。国内外对马铃薯抗黄萎病资源筛选方面的研究相对较少。
3.2 抗黄萎病茄科作物品种的选育
目前,抗黄萎病茄科作物品种的选育已经取得较好的工作基础,很多抗性品种已经成功走向市场。刘厚忠等以日本墨染茄为母本、黑龙江龙茄 1 号为父本育成同时具有长势强、抗倒伏和抗黄萎病特点的品种哈茄V8。杨爱国等利用 Ly06-6和Ly09-1为父母本选育的洛茄5号青茄品种,在田间种植表现良好,抗多种真菌病和病毒病。常见的主栽品种长茄1号、吉茄1号、鲁茄1号、辽茄3号、长茹3号、驻茄11号等品种均较为抗病。
各国栽培的番茄品种中,对田间黄萎病有较好抗性水平的品种有阿莫尔(荷兰)、法宝008(荷兰)、866258(西班牙)等。由国内种业公司选育的T20314、HG1908、HTC750355等品种也对黄萎病具有抗性。
据国外报道,马铃薯品种Alpha、A81473-2、Cal White、Chipteta、Gemchip等对黄萎病均有较好抗性或耐性。我国推广的部分自育品种对黄萎病也有良好的抗/耐性。中薯系列和陇薯系列11个品种(系)在黄萎病病田中不感病或呈轻微症状,表现出高度抗性或耐性,内蒙古自治区主栽品种中克新1号、云薯401、合作88较于其他品种有较好的抗性。
4 展望
近年来,黄萎病在我国发生猖獗,造成茄子、番茄、马铃薯20%~60%不等的产量损失,发病程度可能呈扩大趋势,严重威胁产量。茄科作物抗黄萎病研究已在遗传规律、基因定位、资源筛选、品种选育等方面取得明显进展。但是,要在生产上建立行之有效的黄萎病综合防治体系,仍需在抗病资源筛选、抗病基因挖掘、抗病品种选育等方面加大研究力度。
积极对接茄科作物资源丰富的国家或地区,引入抗病性状综合表现良好的茄科作物资源,运用基因组学等技术手段针对黄萎病进行抗源种质筛选。对马铃薯而言,尤其要突出对野生二倍体资源搜集、鉴定。针对抗黄萎病种质资源开展遗传定位研究,开发有效标记、定位功能位点。在番茄、马铃薯中先后被鉴定出476、1 126个NBS-LRR类抗病基因,与抗黄萎病相关的抗病基因有待进一步验证。高效利用已克隆到的抗性基因资源可以加快抗病品种的选育进程。如从野生秘鲁番茄(Mill.)获得的抗黄萎病基因,已成功转入马铃薯栽培种中,大大提高了马铃薯栽培种对黄萎病的田间抗性水平。随着现代分子生物学技术的发展,对黄萎病病原菌致病机理、茄科作物抗病机理的研究会越来越深入,这将积极推动抗黄萎病新材料创制工作,育成更多具备良好抗性和推广价值的茄科品种将成为可能。
猜你喜欢
环境昆虫学报(2023年4期)2023-10-19
中学生百科·小文艺(2020年4期)2020-01-13
铁道通信信号(2019年7期)2019-10-08
生物技术通报(2018年4期)2018-03-31
广东农业科学(2017年5期)2017-08-29
蔬菜(2017年12期)2017-01-31
