
西藏青稞穗腐病病原菌鉴定与室内药剂筛选
2022-07-29 07:03申李旖琦聂丽妍魏丽萍巩文峰
江苏农业科学 2022年14期
杜 娟, 申李旖琦, 聂丽妍, 魏丽萍, 孙 玉, 巩文峰
(1.西藏农牧学院植物科学学院,西藏林芝 860000; 2.资源与环境学院,西藏林芝 860000)
青稞(var.Hooker f.),别称裸大麦、米麦、元麦,主要分布在青藏高原的西藏和青海一带,是西藏主要的粮食作物之一,占西藏粮食作物总播种面积的61.50%。目前,随着西藏经济的快速发展以及人民生活水平的日益提高,对于青稞等的粮食需求在不断增加,农民种植青稞的生产积极性有明显升高,但随之而来的农作物病虫害成为影响粮食作物产量的阻碍,在众多病虫害中,发现一种由真菌引起的青稞穗部病害——青稞穗腐病,该病害也在玉米、水稻、小麦上普遍发生。1936年我国首次有小麦赤霉病发生的相关报道,该病主要发生在长江中下游麦区,不仅造成小麦的严重产量损失,而且病原菌产生毒素造成人畜中毒。小麦赤霉病是由多种镰刀菌引起的,报道较多的致病菌有禾谷镰刀菌复合群(species complex)、黄色镰刀菌()、串珠镰刀菌()和燕麦镰刀菌()、梨孢镰刀菌()和雪腐镰刀菌()以及其余27种镰刀菌菌种或变种,其中禾谷镰刀菌是我国小麦赤霉病的主要致病种。
近年来,青稞穗腐病是在西藏青稞种植区发现的危害逐渐严重的一种穗部病害,目前对该病害的病原研究还不清晰。由镰刀菌所引起的青稞穗腐病在2009年甘肃省甘南藏族自治州局部青稞地块首次发现,在西藏日喀则、青海省、四川甘孜州等青稞种植区域都有发现,穗腐病的危害不断加重,且流行区域总体不断扩散蔓延。同时,在病害流行区,一般青稞田块病穗率为1%~5%,局部病穗率高达10%~15%。陈琳等确定青稞穗腐病的病原是禾生指葡孢霉,本研究中也分离观察到这种病原,且发现一种新的致病菌。本研究对从西藏自治区山南市隆子县和日喀则市江孜县青稞种植区采集的发病麦穗分离病原菌,对该病原菌鉴定分析,研究碳氮源对病原菌菌丝生长的影响,选取6种不同杀菌剂进行防治试验,以期为青稞穗腐病诊断、防治和抗病机制等工作提供一定的理论依据。
1 材料与方法
1.1 试验材料
试验在2020年8月至2021年7月间于西藏农牧学院植物病理实验室进行。青稞穗腐样品采自西藏自治区山南市隆子县(28°31′35.97″N,92°15′32.37″E)和日喀则市江孜县(28°54′18.95″N,89°37′7.95″E),经纬度取县中其中1个采样地,平均海拔在4 000 m。在每个样点中用五点取样法,从100株青稞中记录发病的青稞株数,取平均值求该样品点的穗腐并发病率并记录。将采集样品分别装袋、编号、记录,置于4 ℃车载冰箱暂时保存,回到实验室,放置于4 ℃冰箱中,以待分离。
选取以下培养基进行分离、鉴定。
马铃薯葡萄糖琼脂培养基(PDA):200 g马铃薯、20 g葡萄糖、7~20 g琼脂1、1 000 mL水。
康乃馨叶片琼脂(CLA):水1 000 mL、琼脂 17~20 g、康乃馨叶片。处理方法:将新鲜的康乃馨叶片用自来水清洗干净,切成5 mm×5 mm左右的小片,放入干燥箱中,45~55 ℃烘干2 h左右,待叶片干燥且不失绿后,用环氧丙烷熏蒸灭菌2 h。将灭菌后的叶片置于灭菌培养皿中,倒入45 ℃ 1.5%~2%水琼脂,培养基冷却后,在靠近叶片处进行接种。
合成低营养琼脂培养基(SNA,用于观察小孢子,厚垣孢子):1 g KHPO、1 g KNO、0.5 g MgSO·7HO、0.5 g KCl、0.2 g葡萄糖、0.2 g蔗糖、20 g琼脂。
查氏培养基(CDA):30 g/L蔗糖、20 g/L琼脂、2 g/L NaNO、1 g/L KHPO、0.5 g/L MgSO·7HO、0.5 g/L KCl、0.01 g/L FeSO。

1.2 病原菌的分离纯化
采用菌丝顶端纯化法分离真菌。将从西藏山南市隆子县和日喀则市江孜县采集的感病麦穗首先剪去多余麦芒,用灭菌蒸馏水冲洗3~4次后用灭菌滤纸吸干其表面水分,然后将患病麦穗放置于培养皿内保湿,待有菌丝长出后进行镜检和形态鉴定。之后用接种针挑取菌丝或孢子接到灭菌PDA培养基上进行分离,待长出菌落进行纯化,并斜面保存在4 ℃冰箱待用。
1.3 形态学鉴定
将菌株接在PDA平皿上,28 ℃培养7 d,并观察其在PDA上的生长性状,如菌落形态、质地、有无色素产生。用接种针挑取菌丝(乙醇灯旁操作),放置于滴在载玻片上的水滴中,在尼康Ni-U生物显微镜下进行观察。参照Leslie的《the Fusarium Laboratory Manual》对分离的病原菌进行形态学鉴定。通过显微镜观察镰刀菌小型分生孢子的形态、大型分生孢子的分隔、弯曲程度、基孢足根以及分枝形态等特征并进行拍照、记录。
1.4 分子鉴定及进化分析
将供试病原菌接种在盛有PDB的液体培养基里,28 ℃、260 r/min振荡培养4 d,过滤收集菌丝。用CTAB法提取病原菌总DNA,利用真菌ITS通用引物ITS1(5′-T C C G T A G G T G A A C C T G C G G-3′)和ITS4(5′-T C C T C C G C T T A T T G A T A T G C-3′)以及 EF-1α 序列:EF-1H(5′-A T G G G T A A G G A A G A C A A G A C-3′)和EF-2T(5′-G G A A G T A C C A G T G A T C A T G T T-3′)进行PCR扩增。使用30 μL PCR反应体系:模板DNA 1 μL、DNA聚合酶1 5 μL、PCR Forward Primer(10 μmmol/L)1 μL、PCR Reverse Primer (10 μmol/L)1 μL、ddHO加至 30 μL。PCR程序如下:94 ℃下预变性5 min;94 ℃下变性30 s,50 ℃下退火30 s,72 ℃下延伸 1 min,35个循环;最后72 ℃下延伸7 min,4 ℃下保存。序列扩增产物经电泳检测后,胶回收得到纯化PCR产物,送公司进行序列测定。
将测序结果通过NCBI数据框(http://www.ncbi.nlm.nih.gov/BLAST/)与已报道菌株进行同源性比较,用MEGA 7.0软件进行多序列比对,采用Neighbor-Joining法和UPGMA法构建系统进化树,进行系统发育分析。
1.5 病原菌生物学特性研究
病原菌生物学特性采用菌丝生长速率法进行,用直径5 mm打孔器打取菌饼,分别接种在以查氏培养基为基础的平板上,分别以葡萄糖、果糖、麦芽糖和淀粉来替换碳源,以牛肉膏、酵母膏、氯化铵、硝酸钾和蛋白胨来替换氮源,28 ℃下培养 7 d,采用十字交叉法测菌落直径,并计算平均值,每个处理3次重复。
1.6 病原菌药剂防治的室内筛选及毒力测定
将以上6种农药统一稀释成1 g/L的农药溶液,将其加入到灭菌未凝固的PDA中,做成农药培养基,静置4 h后进行菌饼接种,在28 ℃中培养5~7 d,记录农药对菌株生长的抑制效果。
通过公式计算菌落生长速率:
菌落生长速率=(-)7。
式中:为菌落直径;为菌饼直径;7为培养天数。
通过公式计算菌丝生长抑制率:
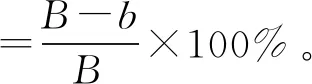
式中:为对照组菌落增长直径;为处理组菌落增长直径。
1.7 数据处理
所得数据利用SPSS 23.0进行数据统计,并采用Duncan’s新复极差法进行多重比较,用OriginPro 2021作图。
2 结果与分析
2.1 危害症状及形态学鉴定
田间采集青稞穗腐病发病症状主要有2种不同的类型,一种是具有粉红色霉层:麦穗从孕穗期表现出症状,包裹麦穗的壳出现病斑。发病部位中央有粉红霉层,四周出现褐色病斑,空穗且发病麦粒的麦芒呈现白化,存在畸形现象。另一种症状是无粉红色霉层:从孕穗期开始发病,初期表现为麦穗上部及麦芒发病,呈现黄褐色病斑,后期发病部位扩散至整个麦穗,黄褐色逐渐转变为白色,严重时出现空穗,发病麦粒的麦芒逐渐白化(图1)。
由表1可知,山南市隆子县的3个样点的穗腐病发病率达5.53%;日喀则市江孜县的穗腐病发病率6.4%。

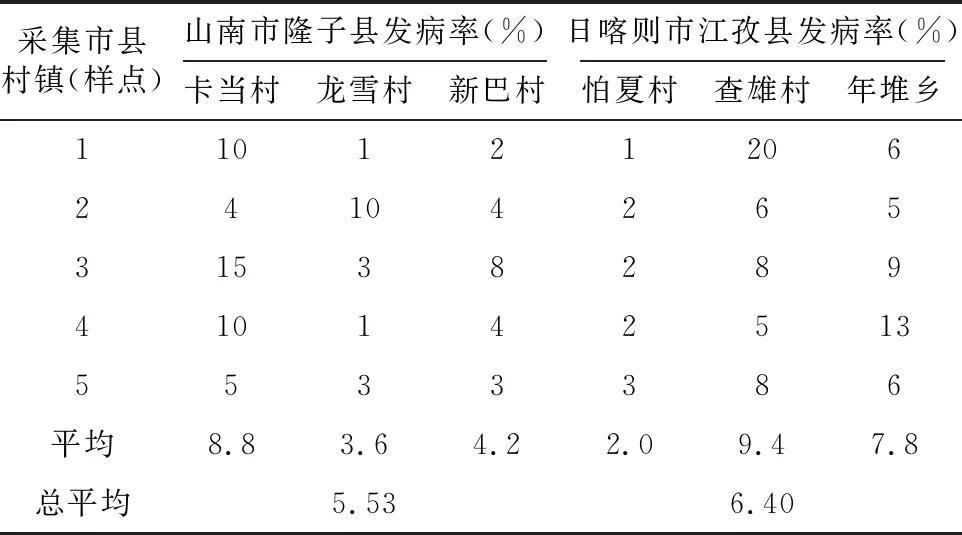
表1 采集点调查青稞发病率
青稞穗腐病病原菌在PDA培养基28 ℃下培养7 d后进行镜检。经过观察,在西藏江孜、隆子2个地方采集的发病麦穗上都是同一类镰刀菌,所以进行形态学和分子鉴定中将不再区分地方。通过在培养基上所观察到的菌落性状为菌落近圆形,菌丝体白色、毛絮状、生长旺盛,而外围一圈的菌丝体生长稀疏,且底部能产生少量红色或黄色色素(图2-C)。通过生物显微镜观察发现,小孢子多,呈卵形、棍棒状,分隔为0~1隔,多数1隔(图2-B)。小型分生孢子的弯曲程度不一,多数较直,分隔多数1~3隔,基孢足根明显(图2-A)。结构情况主要是单瓶梗产孢,厚垣孢子球形,单个顶生(图2-D)。综上,待测菌株与燕麦镰刀菌()形态相似。

2.2 分子鉴定
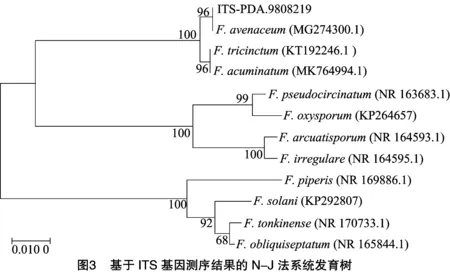
将所采集的青稞发病麦穗上提取的菌株送公司检测,得到DNA测序结果分别进行与NCBI数据库BLAST比对结果,用MEGA 7.0软件进行多序列比对,构建系统进化树。由图3可知,在1 000次重复检验的前提下,分支上侧数字为大于50%的Bootstrap检验值,遗传距离为0.01,供试菌株与登录号为MG274300.1的菌株燕麦镰刀菌处于同一分支,亲缘关系较近,同源性达96%。但该菌株与登录号为KT192246.1的三线镰刀菌()和登录号为MK764994.1的锐顶镰刀菌()也处于同一分支上,且相似性相同。因此ITS基因测序的系统发育树无法区分开该菌株种类,需通过EF-1基因测序结果进一步分析区分。
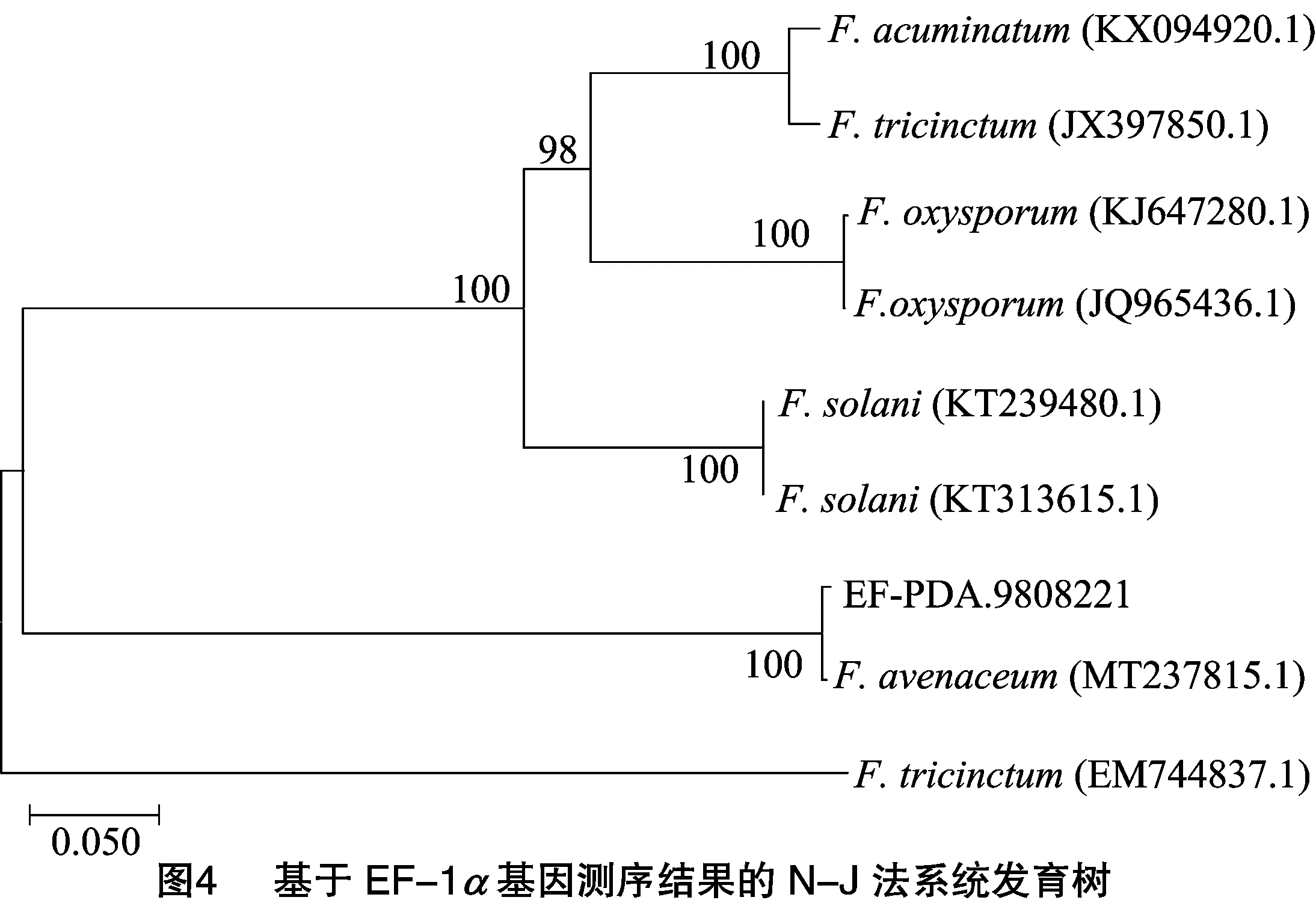
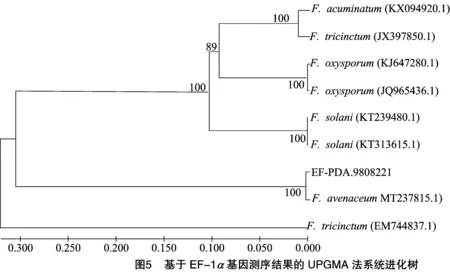
经EF-1测序序列进行比对分析,Bootstrap为1000次检验时,遗传距离为0.05(图4),供试菌株与登录号为MT237815.1的菌株燕麦镰刀菌处于同一分支,且可信度高达100%,亲缘关系更近。进一步分析发现UPGMA法所作系统发育树与N-J法结果一致(图5)。综合以上3种分析结果以及形态学鉴定结果,可以初步得出该穗腐病病原菌为镰刀菌属中的燕麦镰刀菌()。
2.3 不同碳源、氮源对菌丝生长的影响
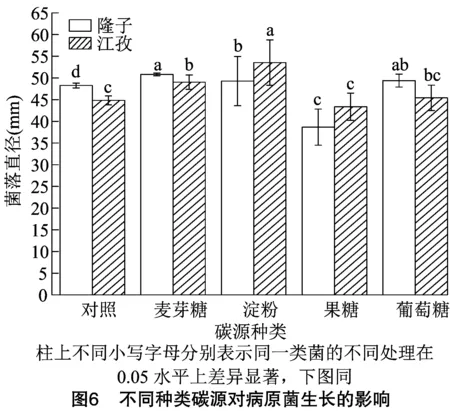
试验设置4个碳源处理,3次重复,在28℃下培养7 d,采用十字交叉法测量病原菌菌落直径结果。结果表明,同一类菌在不同碳源培养基上生长有差异,不同地方采集的同一类菌在同一碳源培养基上生长也有差异。在隆子采集的病原菌最适宜生长的碳源是葡萄糖和麦芽糖;在江孜采集的病原菌最适宜生长的碳源是淀粉(图6)。
试验设置5个碳源处理,3次重复,在28 ℃下培养7 d,采用十字交叉法测量病原菌菌落直径。结果表明,同一类菌在不同氮源培养基上生长有差异,不同地方采集的同一类菌在同一氮源培养基上生长也有差异。在隆子采集的病原菌最适宜生长的氮源是硝酸钾;在江孜采集的病原菌最适宜生长的氮源是蛋白胨(图7)。值得注意的是,氯化铵对于燕麦镰刀菌的作用是抑制的,与其他氮源有明显的不同。

2.5 药剂防治室内筛选
2.6 药剂防治的毒力测定

表2 不同药剂防治室内毒理测定结果
3 讨论与结论
本研究以从西藏山南市隆子县、日喀则市江孜县采集到的青稞感病麦穗上分离到的病原物为对象,研究该病原物的形态和分子鉴定,并初步研究其生物学特性和毒力测定。综合该病原物的形态学鉴定与ITS/EF-1基因序列分子鉴定结果,可初步确定该病原物属于镰刀菌属的燕麦镰刀菌。在生物学特性研究中发现该病原菌能利用葡萄糖、麦芽糖等多种单糖、多糖作为碳源;能利用有机氮蛋白胨和无机氮硝酸钾作为氮源,符合燕麦镰刀菌的特性。这与龚弘强等的研究结果一致。引起麦类穗腐的病原一般都是由多种病原引起的,如河南省小麦的穗腐病病原由5种镰刀菌组成,而在大麦上有11种镰刀菌可引起大麦赤霉病,早在1951年Bakshi认为燕麦镰刀菌()是主要危害大麦的病原物,本研究也主要分离到了燕麦镰刀菌。目前还没有明确燕麦镰刀菌是否是导致青稞穗腐病发生的优势致病病原菌,为进一步验证,还需要经过柯赫氏法则验证。
在室内毒力测定试验中发现,药剂筛选结果与毒力测定结果大致相似,初步确定苯醚甲环唑对燕麦镰刀菌有抑制作用,且抑制效果较其他种类农药好,可能成为防治首选药剂。苯醚甲环唑的田间防效评价试验是接下来的试验方向,为防治青稞穗腐病的农药种类提供参考。
但由于农药在农业中的广泛使用,其毒性和环境污染问题越来越受到人们的关注。苯醚甲环唑是一种广谱杀菌剂,属于三唑类化学物质。苯醚康唑是典型的三唑类杀菌剂之一。这种杀菌剂以种子处理和叶面喷洒的形式对真菌群非常有效。由于其广谱活性,苯醚康唑已被广泛用于各种农作物。三唑类杀菌剂通过抑制细胞色素P450酶(CYP450)活性来抑制麦角甾醇的生物合成,最终抑制真菌的生长。因此,它被广泛用于防治水果、蔬菜和人类的真菌病害。Wang等研究了苯醚甲环唑对人体细胞的毒性,通过对DNA损伤、细胞凋亡等进行了测试。DNA损伤会导致衰老、神经退化、癌症和其他相关疾病。根据DNA损伤的起源,主要包括2个方面:一是环境影响引起的外源性DNA损伤;另一种是自发损伤引起的内源性DNA损伤。农药引起的DNA损伤属于第2类。虽然推荐使用苯醚甲环唑杀菌剂,但现在不提倡大量使用农药进行防治,笔者后续就穗腐病的根际微生物和内生菌进行相关研究,从中可以发现对燕麦镰刀菌有明显抑制效果的拮抗菌株,进行生防制剂的研究。
青稞是西藏地区主要的粮食作物,青稞的产量和品质直接影响当地的经济和百姓的生活水平。燕麦镰刀菌所导致的青稞穗腐病此前在我国西藏地区还未见报道,青稞穗腐病原组成多样性、致病性和致病力目前还不清楚,该病原菌的侵染循环、致病机理以及该病原菌是否是由种子带菌引起的等问题是未来进一步研究的方向。
猜你喜欢
中老年保健(2022年5期)2022-11-25
星星·诗歌原创(2022年1期)2022-04-16
现代畜牧科技(2021年10期)2021-11-19
今日农业(2021年7期)2021-07-28
翠苑(2019年4期)2019-11-11
延河(2017年7期)2017-07-19
西藏科技(2016年8期)2016-09-26
中国酿造(2016年12期)2016-03-01
中国酿造(2014年9期)2014-03-11
食品工业科技(2014年9期)2014-03-11
