
10 年禁牧未改变藏西北高寒荒漠植物水氮利用效率
2022-08-24 05:50牛伟玲陈辉侯慧新郭晨睿马娇林武建双
草业学报 2022年8期
牛伟玲,陈辉,侯慧新,郭晨睿,马娇林,武建双*
(1. 中国农业科学院农业环境与可持续发展研究所,北京 100081;2. 河北师范大学地理科学学院,河北省环境演变与生态建设实验室,河北 石家庄 050024;3. 西藏自治区畜牧总站,西藏 拉萨 850000)
草地是西藏自治区最大的陆地生态系统,总面积约8.8×107hm2,占其陆地面积的72.1%[1]。在气候变化和过度放牧影响下,西藏自治区约1.96×107hm2的天然草地发生了不同程度的退化[1]。草地退化不仅影响畜牧业发展和农牧民生计[2],也威胁着生态系统结构和功能的稳定性[3-4]。因此,探明退化草地恢复机理已成为当前高寒生态学的研究热点[5]。
围栏封育是退化草地生态修复的常用措施之一[6-7],但学术界对其有效性存有争议。有研究发现:围栏封育可以改善植物物种和功能群组成[8-9],提高草地群落水平生产力[10],甚至可通过植物资源利用效率来影响生态系统功能的稳定性[11-12]。但也有研究发现:短期围栏封育不能有效改善群落结构,甚至会提高杂类草的相对优势度[3]。关于围栏封育如何影响植物资源利用效率也尚无定论。碳是构成植物干物质的元素,对植物生长起重要作用。氮是植物生长必要的营养元素,影响叶绿素的合成,与植物光合作用紧密相关[13]。叶片碳含量(leaf carbon content,LCC)和叶片氮含量(leaf nitrogen content,LNC)反映了植物体内的生长代谢状况。稳定碳同位素组成(stable carbon isotopic composition,δ13C)和稳定氮同位素组成(stable nitrogen isotopic composition,δ15N)可表征植物的水分利用效率(water use efficiency,WUE)和氮素利用效率(nitrogen use efficiency,NUE)[14-15]。
水分和氮素作为植物生命活动不可或缺的资源,制约着植物个体、种群、群落乃至生态系统的发育、发展和演替[16]。因此,研究草地植物LCC、LNC、δ13C 和δ15N 对于理解植物WUE、NUE 以及群落演替具有重要意义。许雪贇等[17]研究发现围栏封育可以增加青藏高原植物LCC,但对LNC 无显著影响。勒佳佳等[18]在新疆天山高寒草原研究发现,围栏封育可显著降低建群种羊茅(Festuca ovina)的LCC 和LNC,增加退化期代表性植物鹅绒委陵菜(Potentilla anserina)的LCC。丁小慧等[19]发现,呼伦贝尔草原在群落尺度上放牧和围栏封育样地之间的植物LCC 和LNC 并无显著差异;但围栏封育在种群尺度上显著降低了植物LNC。吕广一等[12]发现,放牧可以降低草甸草原和典型草原LNC 和δ15N,但对荒漠草原植物无显著影响,其将这种差异归因于放牧样地土壤总氮(total nitrogen,TN)和δ15N 的贫化。此外,王亚婷等[20]从植物叶片δ13C 和碳氮比(C/N)的角度探讨了放牧对植物WUE和NUE 的影响,研究发现围栏封育会降低一年生植物的WUE,并提高其NUE。因此,围栏封育效果的差异可能是不同功能群植物的资源利用策略造成的。
西藏自治区是我国五大牧区之一,草地资源丰富,地理条件和气候特征独特。高寒草地对气候变化和人类活动响应敏感[21]。当前研究多利用δ13C 来探究围栏封育对植物WUE 的影响,且集中在温性草原和高寒草甸[20,22]。利用δ15N 来研究植物物种和群落水平的NUE,尤其是在极端干旱的荒漠草原的研究仍然比较缺乏。因此,本研究选择藏西北阿里地区高寒荒漠草原为对象,拟通过LCC、LNC、δ13C 和δ15N 等指标在围栏封育内外对比评价围栏封育对高寒荒漠草原生态系统水分和氮素利用效率的影响。
1 材料与方法
1.1 研究区概况
阿里地区位于西藏自治区西北部,是羌塘高原的核心地带,地势高亢,海拔在4500 m 以上。作为藏西北传统的高寒牧区,阿里地区以高寒荒漠草原为主,群落结构单一,以紫花针茅(Stipa purpurea)或沙生针茅(Stipa glareosa)等寒、旱生植物为主,其也面临着畜牧业发展与草地保护的双重压力[23]。阿里地区属高原干旱气候区,气候寒冷干燥,日照充足。由于远离水汽源地且受喜马拉雅山和冈底斯山脉阻挡,阿里地区降水稀少,多年平均降水量不足400 mm。阿里地区面积3.45×107hm2,草地占其土地面积的70%以上,高寒荒漠草原占其草地总面积的18%[1]。土壤类型以亚高山和高山荒漠草原以及荒漠土为主,质地粗糙、养分贫瘠。
1.2 野外调查
本研究在植物生长旺季2017 年8 月中旬选取3 个典型荒漠草原样地进行调查取样(表1)。自2006 年起,所选样地已经连续围栏封育10 年。样地所在地年均温低于2.5 ℃,生长季平均温度11 ℃;多年平均降水量为260~400 mm,超过85%的降水发生在每年5-9 月植物生长季。

表1 采样点信息Table 1 Sampling location information
在每个样地围栏封育内外选择地势平坦、植物长势均一的区域,分别设置1 个100 m×100 m 的采样区。沿采样区对角线,间隔20~30 m 布设5 个1 m×1 m 的样方。在每个样方中分物种刈割收获植物样品,存放在纸质信封或档案袋中保持干燥。在每个样方内取表层20 cm 土壤,过筛(2 mm)去除植物根系和砾石。土壤样品分为两份:一份自然风干保存,用于测定土壤养分;另一份在野外及时称取鲜重(soil fresh weight,SFW),带回实验室后105 ℃下烘干24 h 后称取干重(soil dry weight,SDW),计算土壤重量含水量(soil water content,SWC):

植物样品带回实验室后在65 ℃下烘干48 h 至恒重,称量并计算地上生物量。叶片样品用去离子蒸馏水清洗,再次烘干后研磨成细粉末,储存在棕色玻璃瓶中用于同位素分析。本研究共采集105 个植物叶片样品,其中豆科植物11 个,禾本科29 个,莎草科13 个,杂类草52 个(表2)。禾本科植物在样地中占绝对优势,其AGB 比例超过70%。围栏封育增加了禾本科和莎草科AGB,却减少了豆科和杂类草AGB。但围栏封育对不同功能群植物AGB 的影响不显著(P>0.05,图1)。

图1 围栏封育内外不同功能群植物地上生物量占比之间的比较Fig. 1 Comparisons of aboveground biomass fraction of different plant functional groups(PFGs)between grazed and fenced sites

表2 不同功能群植物叶片碳、氮含量和同位素组成及其地上生物量Table 2 Leaf carbon and nitrogen content(LCC and LNC),leaf stable carbon and nitrogen isotopic composition(δ13C and δ15N),and aboveground biomass of different plant functional groups(PFGs)
1.3 气象数据
气候数据来源于国家气象科学数据中心(http://data.cma.cn)。首先,从该中心下载2017 年间西藏境内各个气象台站逐日气温和降水资料,将植物生长季定义为每年的5-9 月。然后,计算各气象台站年均温(mean annual temperature,MAT)、年 降 水(mean annual precipitation,MAP)、生 长 季 气 温(growing season temperature,GST)和 生 长 季 降 水(growing season precipitation,GSP)(表1)。使用ANUSPLIN 4.3 薄板样条函数进行气象数据插值。最后,在ArcGIS 10.4 中根据经度、纬度和海拔提取样地匹配的气候数据。
1.4 测定指标与方法
采用重铬酸钾-外加热法测定土壤有机碳(soil organic carbon,SOC)含量。使用2 mol·L-1KCl 溶液浸提,采用双波长紫外比色法测定土壤铵态氮(NH4+-N)和硝态氮(NO3--N)的含量。采用1.0∶2.5(m∶v)土水比,悬液酸度计(PHS-3C 型,上海)测定土壤pH。在中国科学院地理科学与资源研究所理化分析中心采用Flash EA1112 HT 元素分析仪(Flash EA1112 HT,意大利CARLO-ERBA 公司)以及Delta V Advantage 同位素质谱联用仪(Delta V Advantage,美国/赛默飞世尔公司)测定植物LCC、LNC、13C/12C 和15N/14N 的自然丰度比,用δ表示,单位为‰,计算公式[24]如下:

式中:Rsample代表叶片样品13C/12C 或15N/14N;Pee Dee Belemnite(PDB)作为δ13C 的国际标样,出自美国南卡罗来纳州的碳酸盐陨石物质,其RPDB=0.0112372,δ13CPDB=0。大气氮气作为δ15N 的国际标样,其Rair=0.0036765,δ15Nair=0[25]。
1.5 数据处理与分析
群落加权平均值(community-weighted mean,CWM)能很好地反映群落水平植物功能性状的聚合度[26],是常用的性状多样性研究指标。本研究计算每个样方植物LCC、LNC、δ13C 和δ15N 的CWM,公式如下:

式中:Pij是样方j中物种i的权重(以相对优势度计算,即该物种AGB 占样方内所有物种AGB 的百分比);Tij是样方j中物种i的叶片性状具体值;CWMj是样方j中植物叶片性状的群落加权值。
首先,采用t-检验对比围栏封育内外群落水平LCC、LNC、δ13C 和δ15N 的差异。其次,采用双因素方差(Twoway ANOVA)分析检验高寒荒漠草原不同功能群植物LCC、LNC、δ13C 和δ15N 在围栏封育内外是否存在明显分化。然后,利用双变量回归分析探究群落水平LCC、LNC、δ13C 和δ15N 随环境因子的变化规律。最后,采用多元回归模型和方差分解法,量化评估气候和土壤因子对群落水平LCC、LNC、δ13C 和δ15N 的相对影响。根据MASS包中逐步赤池信息准则(step akaike information criterion,step AIC)和pedometrics 包中的逐步方差膨胀系数法(step variance inflation factor,step VIF),采用后向筛选法(backward selection)确定最优模型。采用R 4.1.0 进行数据分析与绘图。
2 结果与分析
2.1 围栏封育对群落水平植物叶片碳氮含量和同位素组成的影响
在群落水平上,围栏封育对LCC、LNC、δ15N 和δ13C 均无显著影响(图2,P>0.05)。围栏封育内LCC(49.25%)略高于围栏封育外(47.24%)。围栏封育内LNC、δ13C 和δ15N(1.65%,-26.53‰,3.13‰)比围栏封育外(1.69%,-26.25‰,3.27‰)略低,但均不显著。

图2 围栏封育对群落水平植物叶片碳氮含量和同位素组成的影响Fig. 2 Effects of livestock exclusion on community weighted means(CWMs)of leaf carbon and nitrogen content(LCC and LNC),leaf stable carbon and nitrogen isotopic composition(δ13C and δ15N)
2.2 围栏封育对不同功能群植物叶片碳氮含量和同位素组成的影响
相比LUT,PFGs 对LCC、LNC、δ13C、δ15N 的影响更大。除LCC 外,围栏封育对LNC、δ13C、δ15N 均无显著影响(P>0.05,表3)。PFGs 和LUT 的交互也仅对LNC 影响显著(P<0.05,表3)。

表3 土地利用和植物功能群对群落水平叶片碳氮含量以及同位素组成的影响Table 3 Effects of land use types(LUT,fenced vs grazed)and plant functional groups(PFGs)on the community-weighted means of leaf carbon and nitrogen content(LCC and LNC)and stable carbon and nitrogen isotopic composition(δ13C and δ15N)
不同PFGs 之间LCC、LNC 和δ15N 差异明显(图3,P<0.05),但同一PFGs 在围栏封育内外无显著差异(P>0.05)。就LCC 而言,禾本科(49.06%)和莎草科(49.36%)植物显著高于杂类草(44.72%)和豆科植物(42.18%)(P<0.05)。就LNC 而言,豆科植物(2.87%)显著高于杂类草(1.87%)、莎草科(1.66%)和禾本科(1.58%),后三者之间仅围栏封育内禾本科植物(1.54%)显著低于杂类草(1.94%),其他两两之间无显著差异。高寒荒漠植物的叶片δ13C 为-28.40‰~-23.15‰,PFGs 之间无显著差异,且围栏封育对其无显著影响。就δ15N 而言,豆科植物(-0.71‰)最小,显著低于杂类草(4.10‰)、莎草科(3.31‰)和禾本科(3.09‰),后三者之间及其在围栏封育内外无显著差异。
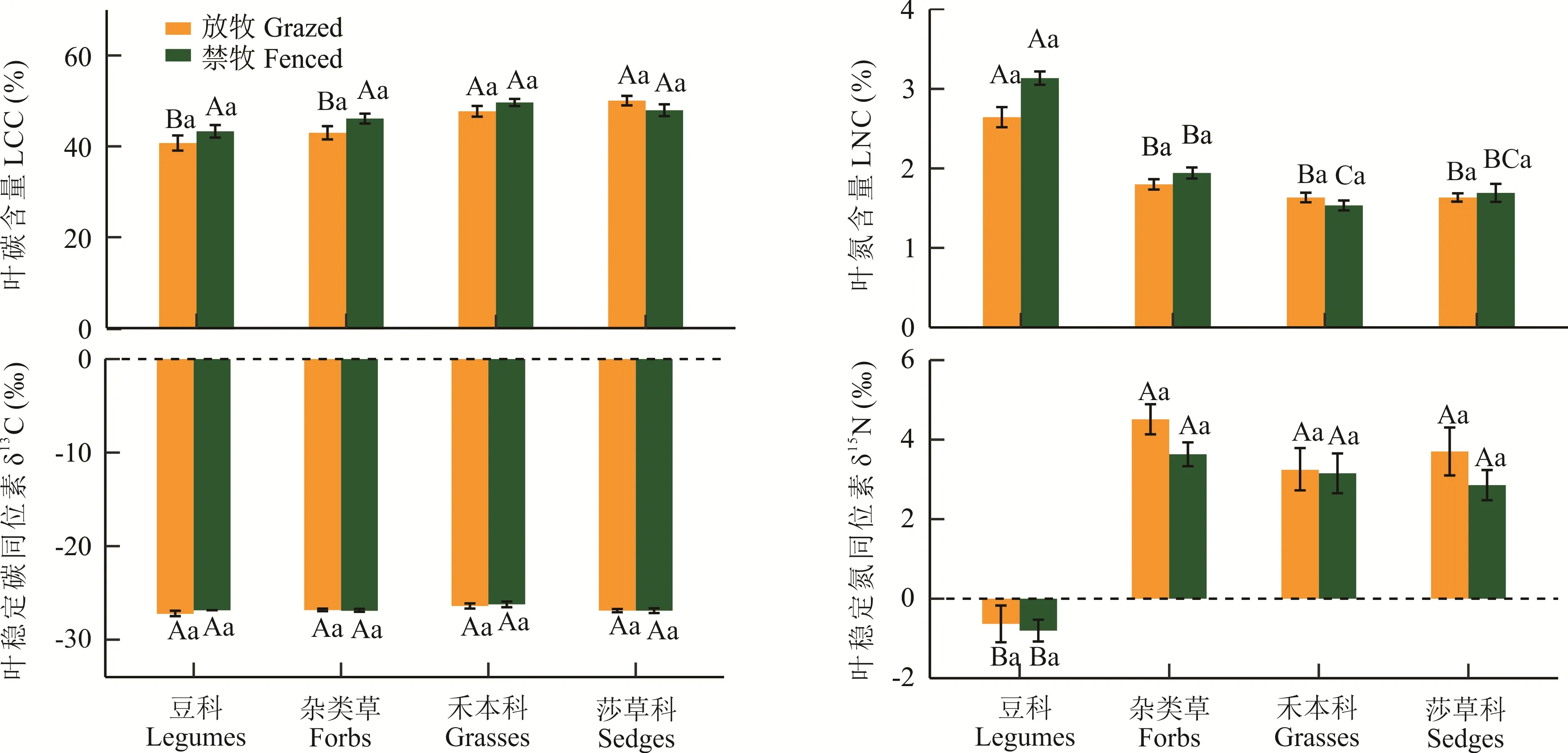
图3 围栏封育对不同功能群植物叶片碳氮含量和同位素组成的影响Fig. 3 Effects of livestock exclusion on leaf carbon and nitrogen content(LCC and LNC)and stable carbon and nitrogen isotopic composition(δ13C and δ15N)of different functional group(PFGs)
2.3 群落水平上植物叶片碳氮含量和同位素组成之间的关系
群落水平上,在围栏封育内植物叶片δ15N 随LCC 增加呈先减小后增加的变化趋势(P<0.05);在围栏封育外放牧样地,则呈先增加后减小的变化趋势,但不显著(P>0.05)。除此之外,其他变量之间均无显著相关关系(P>0.05,图4)。

图4 群落水平上植物叶片碳氮含量和同位素组成之间的关系Fig. 4 Relationship among community weighted means(CWMs)of leaf carbon and nitrogen content(LCC and LNC)and stable carbon and nitrogen isotopic composition(δ15N and δ13C)
2.4 群落水平上植物叶片碳氮含量和同位素组成随环境的变化
除叶片δ15N 与GST 相关性较高外(r=0.87),LCC、LNC 和δ13C 群落加权平均值与环境因子的相关性较小(|r|<0.5)。GSP 与土壤硝态氮,SWC 与SOC,土壤硝态氮和pH 之间共线性较强(|r|>0.8)(表4)。

表4 群落水平叶片碳氮含量和同位素组成与环境变量之间的相关矩阵Table 4 Correlation matrix of community weighted means(CWMs)of leaf carbon and nitrogen content(LCC and LNC)and stable carbon and nitrogen isotopic composition(δ13C and δ15N)as well as environmental variables
群落水平上,叶片δ13C 和LNC 与GST 和GSP 均无显著相关性(图5)。叶片δ15N 随GST 增加呈线性递增,随GSP 增加呈先增加后减小的变化趋势(P<0.01)。在围栏封育内LCC 随GST 增加呈先减小后增加的变化趋势,随GSP 增加呈先增加后减小的变化趋势(P<0.01),这种相关性在放牧样地发生改变。

图5 群落水平叶片碳氮含量和同位素组成与气候因子之间的关系Fig. 5 Relationship among community weighted means(CWMs)of leaf carbon and nitrogen content(LCC and LNC)and stable carbon and nitrogen isotopic composition(δ13C and δ15N)and climate factors
在围栏封育内叶片δ13C 和LCC 分别与SWC 和SOC 呈单峰曲线关系(图6);围栏封育外叶片δ13C 随SWC 增加呈先减小后增加的变化趋势(P<0.05),叶片δ15N 随硝态氮含量增加呈先增加后减小的变化趋势(P<0.1)。在围栏封育内叶片δ15N 随铵态氮含量增加呈线性递增(P<0.05),LCC 分别随SWC 和硝态氮含量增加呈线性递增(P<0.05),叶片δ13C 和LCC 随土壤pH 增加呈线性递减(P<0.05);在围栏封育外叶片δ15N 随土壤pH 增加呈线性递增(P<0.01)。其他变量与土壤因子均无显著相关性。

图6 群落水平叶片碳氮含量和同位素组成与土壤因子之间的关系Fig. 6 Relationship among community weighted means(CWMs)of leaf carbon and nitrogen content(LCC and LNC)and stable carbon and nitrogen isotopic composition(δ13C and δ15N)and soil factors
在群落水平上,气候和土壤因子均不能较好地解释LNC 的变化(P>0.05,表5)。GSP 和硝态氮是影响LCC的主要因子(P<0.05),硝态氮对LCC 变化的解释度大概是GSP 的1.7 倍(表5 和表6)。土壤pH 是调控叶片δ13C 的主要因子,单独解释了其变化的31.9%。叶片δ15N 主要受GST、GSP 和硝态氮的影响,GST 对其变化的解释度最大,大概是GSP 和硝态氮共同作用的11 倍(表5)。

表5 环境因子对群落水平叶片碳、氮含量和同位素组成的相对影响Table 5 Effects of environmental variables on community weighted means(CWMs)of leaf carbon and nitrogen content(LCC and LNC)and stable carbon and nitrogen isotopic composition(δ15N and δ13C)

表6 环境因子对群落水平叶片碳含量和同位素组成相对影响的多元线性最优模型Table 6 Effects of environmental variables on community weighted means(CWMs)of leaf carbon content(LCC)and stable carbon and nitrogen isotopic composition(δ15N and δ13C)in multivariate linear optimal models(MLOM)
3 讨论
3.1 围栏封育对植物叶片碳氮含量的影响
围栏封育对LCC 和LNC 均无显著影响,与丁小慧等[19]在呼伦贝尔草地和鄢燕等[27]在藏北高寒草原的研究结果基本一致。本研究发现,莎草科和禾本科植物LCC 显著高于豆科和杂类草,豆科植物的LNC 显著高于其他物种,但围栏封育对其并无显著影响(图3)。在群落尺度上,围栏封育增加了莎草科和禾本科植物的AGB 占比,降低了豆科和杂类草AGB 比重,但并未改变禾本科植物的绝对优势,对群落组成无显著影响(图1)。这可能是功能群和群落水平上LCC 和LNC 对围栏封育无响应的原因之一。另外,本研究和丁小慧等[19]在群落水平上的研究均是基于植物群落中物种重要值及其养分和同位素指标进行加权计算所得的群落加权均值,植物元素含量存在种间差异且群落结构不同,可能造成群落加权均值无明显差异。群落作为一个整体,需要较长时间才能对环境变化做出响应[28-29]。此外,碳是植物生物量组成的基本元素,与其他元素相比在植物体内所占比例较大(约45%)且较稳定,因此,在放牧干扰下LCC 也能保持相对恒定。
但是,范月君等[30]研究发现,围栏封育可显著提高三江源高寒草甸禾本科和莎草科植物LCC 和C∶N。敖伊敏等[31]在内蒙古典型草原也发现,在群落水平上放牧可降低植物地上部分碳含量。Liu 等[32]研究发现,禁牧对青藏高原中部高寒草原豆科植物LNC 无影响,但可以显著降低非豆科植物LNC,并把这种差异归因于禁牧减少了土壤硝态氮含量,本研究结果与此存在一定差异。与敖伊敏等[31]和Liu 等[32]的研究相比,阿里地区相对更干燥,群落结构更简单,以寒旱生植物为主,单位面积放牧率更低,植物受放牧干扰的程度和几率较小。此外,寒旱生植物的抗牧、耐牧的性能更强,这可能是造成上述差异的原因。
3.2 围栏封育对植物叶片碳氮同位素组成的影响
围栏封育内外叶片δ13C 和δ15N 均无显著差异。本研究发现,同一功能群,叶片δ13C 和δ15N 在围栏封育内外无差异(图3)。豆科植物的δ15N 显著低于其他物种(图3),且其AGB 占比较少(图1),对群落的贡献率较低,不足以引起群落水平δ15N 变化。在不同功能群之间叶片δ13C 也无差异(图3)。因此,在群落尺度上,围栏封育对叶片δ13C 和δ15N 无显著影响(图2)。Wu 等[33]发现在物种水平上围栏封育可能不会影响植物WUE,但可能通过改变不同功能群相对优势度来影响群落水平WUE,本研究结果与此一致。但姚鸿云等[22]对比了我国7 种类型草地放牧前后δ13C 的差异,发现狗娃花属、冰草属、委陵菜属和针茅属植物的δ13C 在放牧区显著升高,点地梅属和羊草的δ13C 在放牧区显著降低。朱国栋等[34]在内蒙古草原的研究发现,放牧显著降低了C3植物叶片δ13C。牲畜采食和践踏破坏草地,会减小植物叶面积,降低叶片气孔导度,减弱CO2和水汽的扩散能力,从而使得细胞间12CO2浓度相对较高[35],进而影响叶片δ13C。但Raven[36]研究发现,在荒漠草原区围栏封育对植物细胞间CO2浓度无显著影响,这可能也是围栏封育对本研究区植物WUE 无影响的原因之一。
吕广一等[12]在内蒙古的研究发现,群落水平上,放牧对荒漠草原植物叶片δ15N 无显著影响。但An 等[37]在宁夏半干旱荒漠草原的研究发现,放牧显著增加了杂类草的δ15N,而降低了豆科植物的δ15N,本研究结果与此不同。造成这种差异的原因可能是不同研究区物种组成存在差异。本研究发现,禾本科植物在荒漠草原群落中占有绝对优势,而豆科植物处于劣势(图1)。围栏封育虽然增加了禾本科和莎草科等可食性牧草AGB 的比重,减少了豆科和杂类草等不可食性牧草,但总体上围栏封育并未显著改变群落物种组成(图1)。
3.3 环境因子对植物水分和氮素利用效率的影响
植物WUE 主要受SWC 和pH 值的调控。植物叶片δ13C 随SWC 先增大后减小(图6)。在干旱条件下,植物根部缺水,导致细胞内溶质减少,水势增高。植物为了减少细胞失水,使得部分气孔关闭,增加了WUE,从而增大了叶片δ13C[38]。随着SWC 增加,叶片气孔开放,植物的蒸腾速率的增加幅度大于净光合速率,植物WUE 减小[39]。姬兰柱等[40]对干旱胁迫下水曲柳(Fraxinus mandshurica)光合特征的研究发现,在中度水分胁迫下水曲柳的WUE 达到最大值,在重度水分胁迫下其WUE 反而下降,本研究结果与其一致。植物叶片δ13C 随pH 值线性递减。研究表明,土壤碱化程度越高,使土壤团粒结构不断消失,导致土壤通透性降低,从而影响植物的水肥供应,对植物生长不利[41]。此外,随着土壤pH 值增大荒漠草原土壤有机碳和全氮含量均降低[42-43]。植物氮主要来自土壤,并参与叶绿体内叶绿素的合成,与植物光合作用中碳的固定密切相关[13],进而影响叶片δ13C。
GST 是植物NUE 的主要驱动因子。本研究发现,GST 单独解释了叶片δ15N 变化的75.3%(表6)。植物叶片δ15N 随着GST 增加逐渐增大,随GSP 增加先增大后减小(图5)。这与前人在全球[44-45]及青藏高原[46]的研究一致,表明干热的环境下氮循环更加开放[44]。此外,Wu 等[46]、Yang 等[47]和Zhou 等[48]的研究也发现,气候因子是影响青藏高原高寒草地生态系统氮循环的主要环境因子。在干旱或极端干旱的地区,温度升高可促进氮挥发,土壤中14N 由于质量较轻优先从土壤中散失,从而使得15N 在土壤中更加富集[49]。植物主要从土壤中获取氮源,植物δ15N 也因此增大。此外,放牧样地叶片δ15N 随GST 的变化率更快(P<0.01)。放牧通过牲畜的采食、践踏和排泄等行为加快净氮硝化和矿化速率,使得氮循环速率加快[32]。
本研究发现,植物LNC 和δ15N 与δ13C 值均无显著相关性(P>0.05,图4),表明高寒荒漠植物WUE 和NUE之间并不存 在权衡(trade-off)关 系。Peri 等[25]在阿根廷森林生态系统的研究和Lopes 等[50]对小麦(Triticum aestivum)的研究均发现,叶片δ15N 与δ13C 存在显著的正相关性。熊鑫等[51]在鼎湖山森林生态系统的研究发现,主要优势树种叶片δ13C 与C∶N 呈极显著正相关关系。以上研究均表明植物生长并非受WUE 和NUE 的补偿制约。而Chen 等[52]和Field 等[53]在草原和森林生态系统的研究均发现,具有较高WUE 的植物其NUE 较低,两者存在补偿机制,这主要取决于植物叶片的生理特性。本研究结果与上述结果均不一致,表明植物对氮素的利用方式不同,其资源利用策略有所差异,WUE 和NUE 并非简单的竞争或权衡(trade-off)关系。
4 结论
围栏封育是一种人为管理草地的措施,有助于科学平衡资源利用与生态系统生产力之间的关系。本研究基于LCC、LNC、δ13C 和δ15N,分析了围栏封育对藏西北高寒荒漠草原生态系统WUE 和NUE 的影响。研究发现,在群落和功能群水平上,围栏封育对植物LCC、LNC、δ13C 和δ15N 均无显著影响。群落水平上,SWC 和pH 值是影响叶片δ13C 的主要因子,表明植物内部WUE 主要受土壤因子的调控。GST 是影响叶片δ15N 的最主要的环境因子,表明气候因子是调控青藏高原高寒草地生态系统氮循环的主要因子。此外,围栏封育并未显著改变物种和功能群的生理指标和功能性状,且荒漠草原植物WUE 和NUE 之间无权衡关系。因此,从长远来讲,短期围栏可能对荒漠草原生态系统无显著影响。
猜你喜欢
当代水产(2022年8期)2022-09-20
中国音乐学(2022年2期)2022-08-10
红蜻蜓(2020年12期)2020-06-20
学校教育研究(2020年7期)2020-04-09
好孩子画报(2019年8期)2019-09-19
分析化学(2019年3期)2019-03-30
发明与创新·大科技(2018年2期)2018-03-17
中学生理科应试(2017年6期)2017-09-27
分析化学(2017年5期)2017-06-21
中学生理科应试(2017年2期)2017-04-01
