
尾巨桉与红锥混交对林下植被和土壤性质的影响
2022-09-02 08:46王磊温远光周晓果朱宏光孙冬婧
生态环境学报 2022年7期
王磊,温远光, ,周晓果,朱宏光, ,孙冬婧
1.广西大学林学院/广西森林生态与保育重点实验室,广西 南宁 530004;2.广西科学院生态环境研究所,广西 南宁 530007;3.广西友谊关森林生态系统定位观测研究站,广西 凭祥 532600
桉树人工林是当今全球生长速度最快、产量最高、用途最广泛、经济效益较好的人工林之一(Turnbull,1999;Forrester,2013),全球桉树人工林面积已达2500×104hm2,年产木材约占世界人工林的37%(温远光等,2018)。近年来中国桉树人工林面积不断扩大,到2018年,全国桉树种植面积已达 546.74×104hm2,主要分布在广西、广东、海南、福建等沿海省区,以广西的种植面积最大,达256.05万 hm2(国家林业和草原局,2019)。虽然桉树人工林面积仅占中国森林面积的 2.48%,其每年提供的木材却占全国木材产量的30%(中国林学会,2016)。发展桉树人工林对保障国家木材安全、促进天然林保护、应对气候变化以及助力实现碳中和目标均具有不可替代的重要作用(温远光等,2020a)。然而,由于长期以来桉树人工林采取单一种植、短周期经营和高代次连栽方式,导致林下植被退化、植物多样性降低、植物功能群改变、外来植物入侵加剧、土壤性质持续恶化等问题(Wen et al.,2010;温远光等,2005;温远光等,2018),高代次连栽还导致林分生产力显著降低(温远光等,2020b),严重影响着桉树人工林的可持续经营,危及桉树种植区的生物安全、土壤安全和生态安全(温远光等,2018;温远光等,2020b),从而引起国际社会的普遍担忧和广泛关注(温远光,2008;温远光等,2020b)。
植被与土壤是一个有机整体,林下植被是人工林生态系统的重要组成部分,在增加生物多样性(Wu et al.,2011)、改善立地条件(Rothe et al.,2001;黄宇等,2005)、促进养分循环与能量流动(Rivaie,2014)、维持生态系统功能稳定性(周晓果,2016)等方面发挥着不可替代的作用。国内外学者对桉树人工林与林下植被和土壤理化性质的关系开展了大量的研究,主要认为桉树人工林生态问题产生的根源是短周期、高代次纯林连栽的传统营林方式,而林下植被退化、功能群改变、植物多样性显著降低进一步引起土壤理化性质恶化和林分生产力降低(Zhou et al.,2020;李朝婷等,2019;温远光等,2020b)。尾巨桉(Eucalyptusurophylla×E.grandis)是广西最主要的桉树造林树种之一,近年来人们探索通过生态营林、营造混交林、降低林地干扰、延长经营周期等多种措施来权衡桉树木材生产与林下植被和土壤性质的关系,以实现桉树人工林的可持续经营,取得了显著的成效(温远光等,2020a;温远光等,2020b;陈秋海等,2022;邵文哲等,2022),但就尾巨桉与乡土珍贵树种混交对林下植被物种多样性、土壤理化性质和生物量的综合研究较少。因此,本研究以尾巨桉纯林、红锥(Castanopsishystrix)纯林和尾巨桉-红锥混交林为对象,探究尾巨桉与红锥混交对林下植被物种多样性、生物量和土壤理化性质的影响,旨在为桉树人工林的可持续发展提供科学依据。
1 研究区域概况与研究方法
1.1 研究区概况与试验设计
研究区域位于广西凭祥市,中国林业科学研究院热带林业实验中心青山实验场(21°57′47″—22°19′27″N、106°39′50″—106°59′30″E)。低山丘陵地貌,海拔130—1045.9 m,土壤主要为砖红壤和赤红壤。地处南亚热带季风气候区,年均气温20.5—21.7 ℃;极端最低气温-1.5 ℃,极端最高气温40.3 ℃;雨热同季,干湿季分明,年均降雨量1200—1500 mm,4—9月为雨季;年均蒸发量1261—1388 mm,相对湿度80%—84%;为南亚热带半湿润-湿润气候。地带性森林植被为南亚热带季雨林,已被破坏殆尽,现存森林植被主要是马尾松(Pinus massoniana)、杉木(Cunninghamialanceolata)、桉树(Ecalyptusspp.)、红锥、格木(Erythrophleum fordii)等为优势的各种热带亚热带人工林。
选择青山实验场67林班为试验区域,原立地植被为35年生的马尾松人工林,总面积45 hm2。2011年冬季对马尾松人工林采伐后进行人工清理和带状整地,带宽1 m,种植穴规格为50 cm×50 cm×30 cm,种植前7 d,每穴施250 g复合肥作为基肥。2012年3月,采用随机区组设计,共5个实验区组,每个区组分别营造尾巨桉纯林(PEU)、红锥纯林(PCH)和尾巨桉-红锥混交林(MEC),每种林分实验面积为 2 hm2。尾巨桉、红锥纯林的造林密度均为 1428 plant·hm-2,设置宽窄行,窄行行距为2 m,宽行行距为5 m,株距为2 m。为了保持桉树人工林的木材产量,混交林中,尾巨桉按纯林规格造林,红锥种植于宽行之间,株距为2 m,密度为714 plant·hm-2,混交比例为 2∶1。对所有林分均采用“三低三高”的生态营林方式(温远光等,2020b)。2012年和2013年秋季进行人工除草抚育。2013年秋季过后,停止人为抚育和干扰。据2012年对5个实验区组的土壤采样测定,0—20 cm 土层土壤有机碳变化于 18.75—23.52 g·kg-1、土壤全氮为 1.70—2.10 g·kg-1、全磷为 0.31—0.42 g·kg-1、全钾为 16.70—18.30 g·kg-1、pH 为 4.5—5.0,不同区组各指标差异不显著(周晓果,2016)。
1.2 样地设置与群落调查
2019年5月,在上述3种林分中选择立地条件基本一致的代表性区域,分别随机设置5个20 m×20 m的调查样方,共计15个样方。不同林分样方概况见表 1。在对样方内林木进行每木调查的基础上,分别选取尾巨桉、红锥标准木25株和15株(按林分平均木选取,混交林每个样方4株,其中桉树3株、红锥1株;桉树和红锥纯林每个样方各2株),伐倒标准木后,按分层切割法测定地上部分生物量;将根系全部挖出,测定地下部分生物量;并按干材、干皮、树枝、叶和根分别取样500 g,带回实验室于85 ℃下烘干至恒质量,测定含水率,进而计算乔木层生物量(温远光等,2020b)。在每个400 m2样方中随机设置 3个 5 m×5 m的灌木层样方和 3个 2 m×2 m的草本层样方,分别对样方内物种进行全面调查,记录种名、个体数、高度和盖度等。采用全收获法测定样方内所有木本、草本植物地上部分和地下部分的生物量,称鲜质量,采集各部分样品500 g,带回实验室,置于85 ℃下烘干至恒质量,计算单位面积干物质的质量。
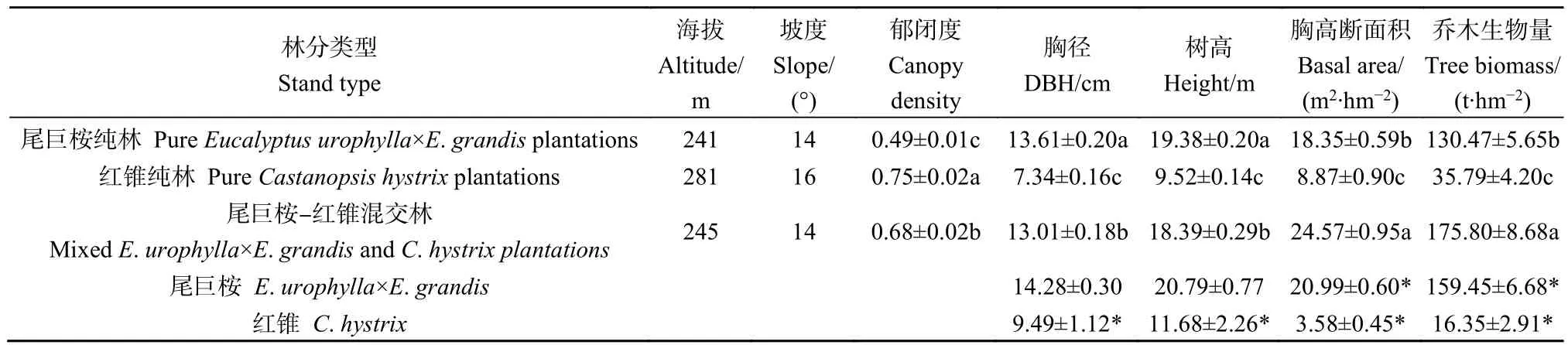
表1 不同林分试验样方概况Table 1 Characteristics of the experiment plots in different stands
1.3 土壤样品采集与测定
在每个400 m2样方中随机选择3个土壤剖面,按 0—20、20—40、40—60、60—80、80—100 cm采集土壤样品1 kg,去除植物根系及石砾后各层分别混合均匀,过2 mm孔径筛,用于测定土壤理化性质。
采用环刀法测定各土层的土壤容重,土壤含水量采用烘干法测定,土壤pH值采用pH计(starter 2100,Ohaus,USA)测定(m(soil)∶V(water)=1 g∶2.5 mL),有机碳采用 K2Cr2O7-H2SO4外加热法测定,全氮和有效氮(铵态氮和硝态氮)分别采用H2SO4-混合催化剂(m(K2SO4)∶m(CuSO4)=10∶1)消煮、0.2 mol·L-1KCl溶液提取后于连续流动分析仪(AA3,Bran Luebbe公司)测定。全磷采用H2SO4-HClO4消煮、钼锑抗比色法测定,速效磷采用双酸(HCl-H2SO4)浸提、钼锑抗比色法测定。全钾采用NaOH熔融、火焰光度法测定,速效钾采用NH4OAc浸提、火焰光度法测定(鲍士旦,2018)。
1.4 数据处理与统计分析
根据样方数据,计算各林分的物种丰富度指数R、Shannon-Wiener指数H、Simpson指数D、Pielou均匀度指数J,计算公式如下:


其中:
S——样地出现的物种数,;
ni——样地内某种的个体数;
N——样地内总个体数。
在SPSS 24.0 for Windows中,采用单因素方差分析(One-way ANOVA)检验不同林分林下植被群落物种多样性、生物量、土壤理化性质的差异显著性,采用LSD法进行多重比较,显著性水平设置为P<0.05。采用冗余度分析确定影响林下植被群落物种多样性变异的主控因子,在R3.5.2的vegan程序包(Dixon,2003)中,通过向前选择(forward selection)法和蒙特卡洛检验(Monte Carlo tests,循环次数999)筛选影响显著(P<0.05)的生物及非生物因子,获得最优RDA模型。在rdacca.hp程序包中采用层次分割理论分解 RDA最优模型中每个解释变量的解释率(Lai et al.,2022)。采用vegan程序包的varpart函数对土壤理化因子、生物量因子进行方差分解分析,并用韦恩图展示两类因子的纯效应和共同效应。
2 结果与分析
2.1 不同林分林下植被群落物种多样性指数
图1显示,3种林分的物种丰富度指数存在显著差异,混交林(MEC)和尾巨桉纯林(PEU)灌木层的物种丰富度显著高于红锥纯林(PCH),前两者无显著差异;PCH和MEC草本层的物种丰富度显著高于PEU,PCH与MEC差异不显著;MEC林下植被群落的物种丰富度显著高于两种纯林,两种纯林间差异不显著(图1)。3种林分林下植被群落的Shannon-Wiener指数、Simpson指数均以PCH的最高,MEC最低,PEU居二者之间,但差异不显著;而 3种林分林下灌木层的 Shannon-Wiener指数、Simpson指数均存在显著差异,MEC显著高于PCH,与PEU差异不显著;PEU灌木层的Shannon-Wiener指数显著高于PCH,但Simpson指数无显著差异。3种林分草本层的 Shannon-Wiener指数、Simpson指数变化与林下植被群落一致。3种林分林下植被群落和草本层的均匀度指数存在显著差异,而灌木层的均匀度指数却无显著差异;PCH林下植被群落和草本层的均匀度指数显著高于MEC,MEC与PEU差异不显著。
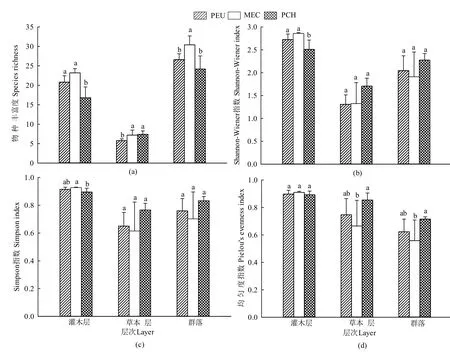
图1 不同林分林下植物多样性指数Figure 1 Plant diversity indices in the understory of different stands
2.2 不同林分林下植被生物量
不同林分林下植被生物量呈现 PEU (4336.37 kg·hm-2)>MEC (1848.43 kg·hm-2)>PCH (1197.11 kg·hm-2),方差分析表明,PEU显著高于 MEC和PCH,后两者差异不显著(图 2)。3种林分灌木层地上部分生物量的变化规律与林下植被群落的相似,也呈现出PEU显著高于MEC和PCH,而MEC和PCH差异不显著;灌木层地下部分生物量为 PEU显著高于 MEC和 PCH,MEC显著高于PCH。PCH草本层地上部分生物量显著高于MEC和PEU,后两者差异不显著;MEC和PCH草本层地下部分生物量显著高于PEU,MEC与PCH差异不显著(图2)。
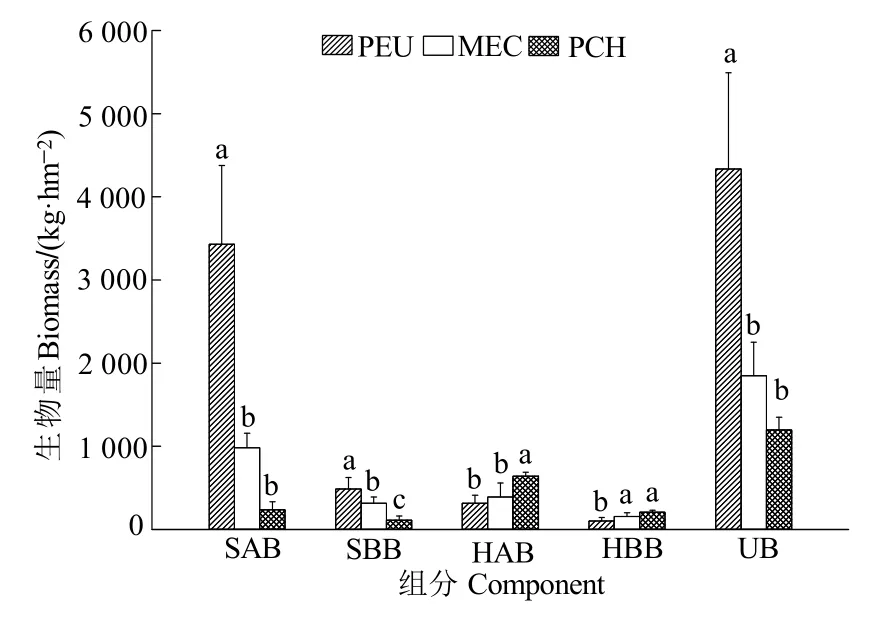
图2 不同林分林下植被的生物量Figure 2 Biomass of understory vegetation in different stands
2.3 不同林分的土壤理化性质
表2显示,MEC 0—100 cm土层的pH值显著高于PEU和PCH,PCH 0—20 cm土层的pH值高于PEU,20—100 cm土层则相反,为PEU大于PCH,其中20—40、40—60、80—100 cm土层达到显著差异。3种林分的土壤含水量,除了MEC 0—20 cm土层显著高于 PEU、MEC和 PCH 60—80 cm、80—100 cm显著高于PEU外,其余土层差异不显著。3种林分的土壤容重以PEU较高,其中,除了MEC 20—40 cm与40—60 cm土层差异不显著外,其余土层均为PEU显著高于MEC和PCH(表2)。3种林分的土壤有机碳含量,除了个别土层(MEC 80—100 cm)外,MEC和PCH各土层的土壤有机碳含量均显著高于PEU。PCH 40—60、80—100 cm土层的全氮显著低于 PEU和 MEC同一土层,PEU和PCH 60—80 cm显著低于MEC,其余林分和土层均无显著差异。PEU和MEC各土层的全钾含量显著高于PCH,前两者差异不显著;PCH 60—80、80—100 cm土层的速效钾显著低于其他两种林分,其余各林分和土层间差异不显著。3种林分各土层的全磷含量均无显著差异,而 MEC各土层有效磷含量显著高于两种纯林(PCH 40—60 cm除外),PEU 40-60 cm土层的有效磷与PCH差异不显著,其余土层均表现为PCH显著高于PEU(表2)。
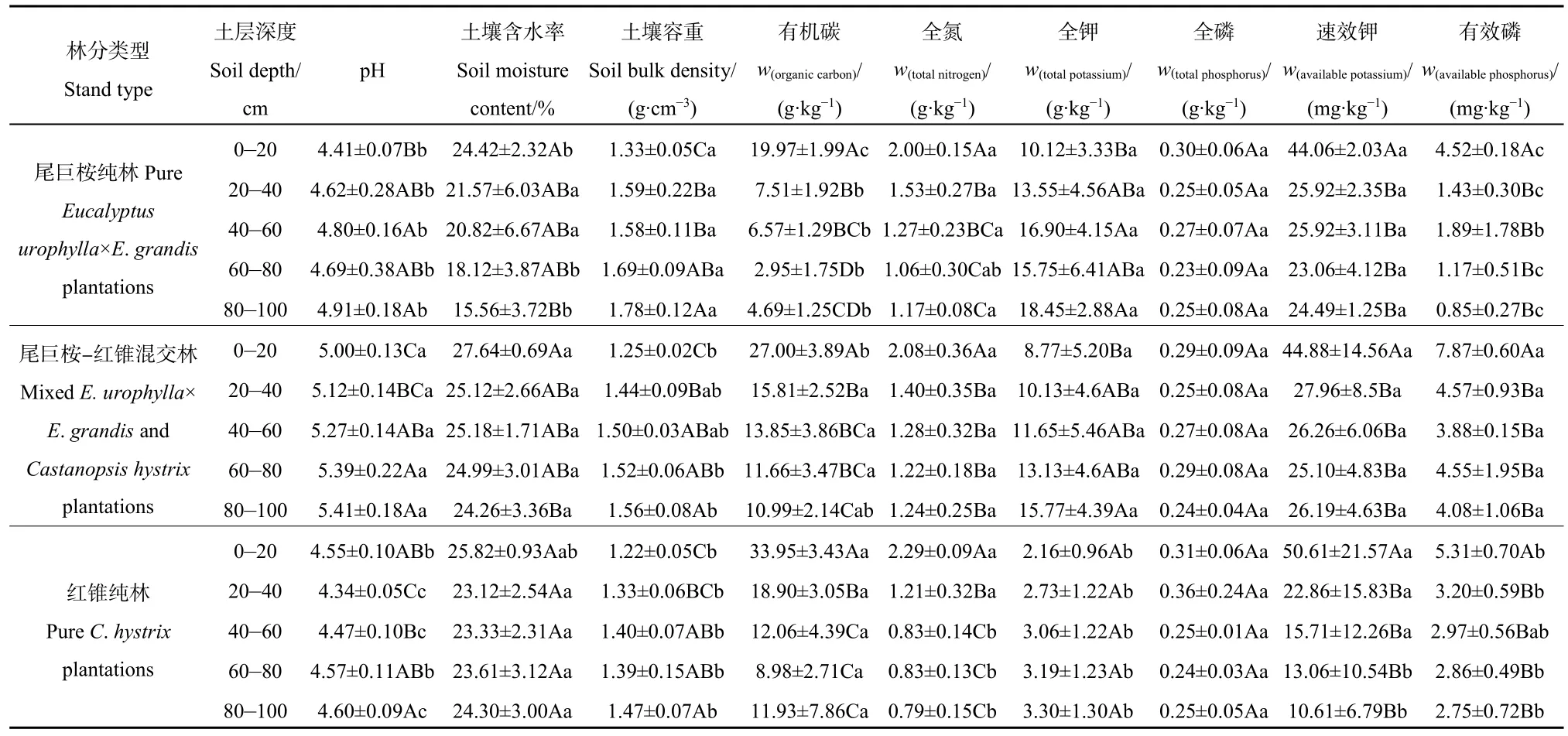
表2 不同林分的土壤理化性质Table 2 Soil physicochemical properties in different stands
2.4 不同林分林下物种多样性与生物量和土壤理化性质的关系
林下植物群落物种多样性与土壤理化性质和生物量的冗余度分析结果表明,RDA第一、二轴分别能解释林下植物群落多样性变异的71.90%、11.31%(图 3a),累计解释率为 83.21%,占总解释率的96.55%。置换检验显示,林下植物群落物种多样性与土壤、生物量因子在RDA轴1和所有RDA轴分别为显著(P=0.014)和极显著相关(P=0.007),表明 RDA排序结果能较好地解释林下植被物种多样性与土壤、生物量因子间的关系,且其相关性主要由第一排序轴决定。
影响林下植物群落物种多样性的主控因子为灌木层地下部分生物量(SBB)、草本层地上部分生物量(HAB)、乔木层地上部分生物量(TAB)、土壤C∶N、土壤全氮(TN)、土壤N∶P和pH(表3)。其中,TAB、SBB、TN、pH、N∶P与第一排序轴呈显著负相关(图3a),单独分别能解释林下植被物种多样性变异的12.9%、9.9%、13.9%、8.5%和6.2%(图3b),与PEU和MEC林下植物群落、灌木层的物种丰富度呈显著的正相关关系。HAB、C∶N与第一排序轴显著正相关(图3a),单独分别能解释林下植被物种多样性变异的 19.6%、9.2%(图3b),与PCH草本层的物种丰富度呈显著的正相关关系。灌木层、草本层及林下植物群落Shannon-Wiener指数、Simpson指数和均匀度指数主要受SBB、HAB的显著影响(图3a)。将影响林下植物群落物种多样性的主控因子进一步归为土壤理化性质和生物量因子进行方差分解分析(图3c),结果显示,土壤理化性质、生物量因子对林下植物群落物种多样性变异的解释率分别为12.2%、21.3%,两者能够共同解释林下植物群落物种多样性变异的34.2%。
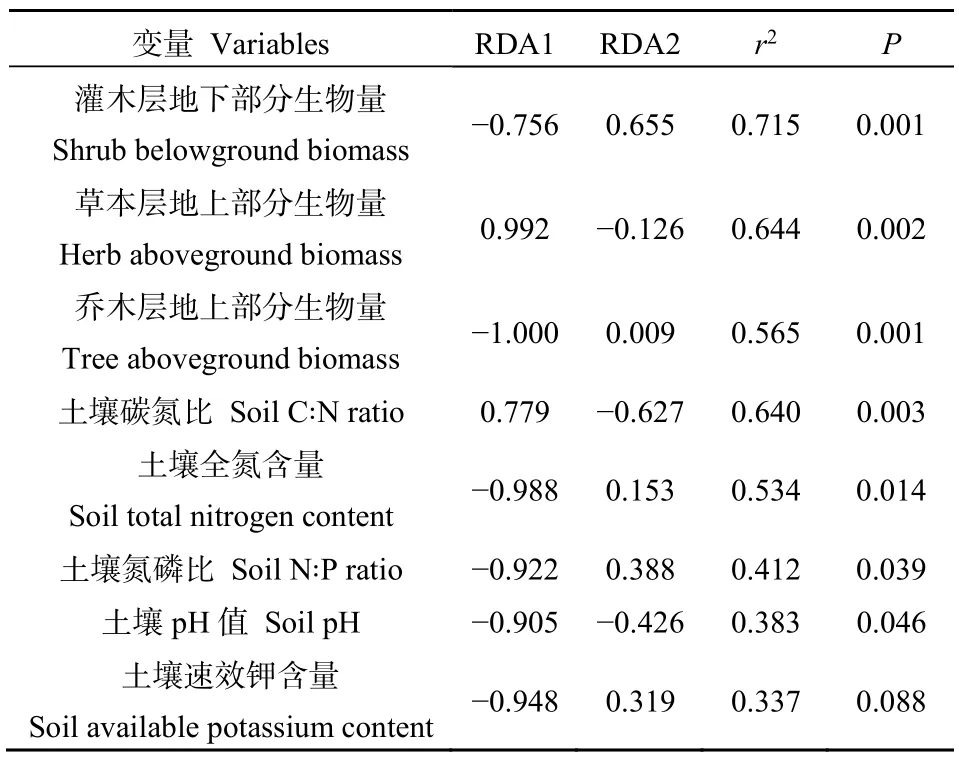
表3 土壤及生物量因子与排序轴的相关性Table 3 Correlation between soil, biomass factors and ordinate axes
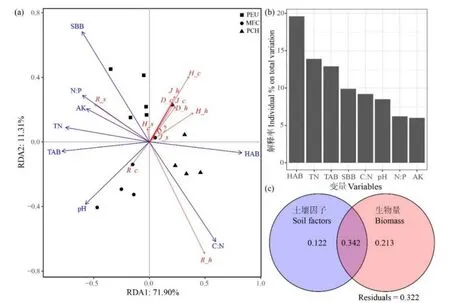
图3 林下植被物种多样性与土壤因子和生物量的冗余度分析(a)及影响因子的方差分解(b、c)Figure 3 Redundancy analysis of understory species diversity, soil factors and biomass (a) and variance partitioning of the influencing factors (b, c)
3 讨论
3.1 尾巨桉与红锥混交对林下植物多样性的影响
林下植物群落的物种多样性是人工林可持续经营的重要内容,也是认识群落组成结构和组织水平的基础。林下植被群落的物种多样性受诸多因素的影响,常因树种、林分类型或经营管理水平不同而异(周晓果,2016;温晶等,2019;舒韦维等,2021)。一般情况下,混交林通过改变树种组成和林分结构,使林分结构得以调整和优化,促进生态位的分化,增进物质循环,改善林地土壤理化性质,有利于更多的物种定居,从而提高林下物种多样性(Levick et al.,2015;孙冬婧等,2015;陈科屹等,2017)。本研究结果与上述结论基本一致。本研究发现,尾巨桉与红锥混交显著提高林下植被群落、灌木层和草本层的物种丰富度,对灌木层的Shannon-Wiener指数、Simpson指数也有一定的正效应,而对草本层却存在负效应;混交林对林下植被群落、灌木层和草本层的物种均匀度指数也存在负效应。同时还发现,两种纯林都存在降低灌木层多样性、提高草本层多样性的趋势。究其原因,可能是因为PEU的林分郁闭度较小(0.49),林下光照充足,在生态营林方式下,林分抚育强度明显减弱,灌木层中阳性的大型木质藤本,如羽叶金合欢(Acaciapennata)、钩藤(Uncariarhynchophylla)等特别发育,有的缠绕在桉树的树干上高度达15 m以上,阴生或中生性的物种受到限制,导致林下植被群落物种多样性的降低;与之相反,PCH林分郁闭度较高(0.75),林下光照明显减少,阳性的植物种类受到抑制,但阴生性的植物种类,特别是耐阴的蕨类植物,如乌毛蕨(Blechnopsisorientalis)、金毛狗(Cibotiumbarometz)等在草本层占居明显优势,从而增加草本层物种多样性;MEC林分郁闭度适中(0.68),混交形成的双层林冠结构既显著地遏制阳性大型木质藤本的生长,显著降低其重要值,同时也能为阳性与阴性物种提供适宜的生态位和资源,使得林下植被的植物物种数量最丰富,达77种,分别比PEU(61种)和PCH(60种)提高26.23%和28.33%。本研究结果说明,在南亚热带区域,桉树人工纯林采取的“双龙出海”模式和宽窄行配置,使得宽行内的光照特别充足,同时又采取生态营林方式经营,对林分的抚育强度明显减弱,导致宽行内的阳性大型木质藤本大量定居和繁茂生长,加剧了桉树与大型木质藤本的资源竞争,从而降低桉树的生长量和生物量(表1)。然而,采取“双龙出海+珍贵树种”的生态营林方式,可以较好地调节林分的光照强度和分布,有利于遏制阳性大型木质藤本生长,增加生态位的分化,改善凋落物的数量和质量,增进物质循环,改善林地土壤理化性质,有利于更多的物种定居,增强生物多样性,提高生态系统稳定性。这与中性理论是一致的(Catford et al.,2012)。
3.2 尾巨桉与红锥混交对林下植被生物量的影响
林下植被生物量是评价森林群落物质生产及能量平衡的基础,也是植物资源利用、生态位占有、种内种间竞争能力的体现(Tran et al.,2013)。林分的郁闭或者稀疏,都会直接影响林下植被的生长发育,从而引起灌木层、草本层生物量的差异(Lai et al.,2022)。本研究中,不同树种或林分类型对林下植被生物量的影响存在差异,PEU显著提高林下植被灌木层生物量、降低草本层生物量;PCH显著降低灌木层生物量、显著增加草本层生物量;而MEC则居二者之间,在较高产量水平上维持灌草生物量的相对平衡。本试验区位于南亚热带,光照和热量充沛,PEU林冠较稀疏,林下光照充足,在生态营林降低林下植被抚育情况下,林下出现大量的阳性大型木质藤本和阳性灌木及小乔木,如羽叶金合欢、钩藤、阔叶瓜馥木(Fissistigmachloroneurum)、黄毛榕(Ficusesquiroliana)、毛稔(Melastoma sanguineum)等,而且生长繁茂,生物量高,PEU林下植被灌木层生物量(3917.27 kg·hm-2)占林下植被生物量的 90.33%;灌木层地上部分生物量(3430.81 kg·hm-2)是地下部分生物量(486.46 kg·hm-2)的7.05倍,根冠比(地下与地上生物量的比例关系)为0.14,远低于中国森林生态系统的根冠比中值(0.23)(王丽美等,2020)。究其原因,与林下出现大量的阳性大型木质藤本,如羽叶金合欢、钩藤、阔叶瓜馥木等有关。这些物种的地上部分生长十分繁茂,有的攀援生长的高度达15 m,导致地上与地下部分的生物量比例严重失衡。尾巨桉林下灌木层的高生物量增长强烈的遏制了草本层生物量的发育,使得草本层的生物量显著下降(419.10 kg·hm-2),成为3种林分中草本层生物量最少的类型。
PCH林冠郁闭度较大,抑制了林下阳性大型木质藤本和阳性灌木植物的定居和生长,导致灌木层生物量下降,仅为348.98 kg·hm-2,是3种林分中最低的;相反,PCH林下草本植被特别发育,以中生性的高草植物为优势,草本层生物量 848.13 kg·hm-2,明显高于灌木层生物量,成为3种林分中草本层生物量最高的类型。这与林下光照条件明显不足有关。
MEC林下植被生物量,无论是灌木层还是草本层都是处于两种纯林之间,灌木层生物量显著低于PEU,则显著高于PCH;相反,草本层生物量显著高于PEU,却显著低于PCH。这是因为MEC的郁闭度居两种纯林之间,为0.68,比较适中,既能够较好的遏制阳性大型木质藤本的生长,也能让中生性灌草植物良好生长,通过增加物种丰富度来提高林下植被的生物量;此外,MEC灌木层和草本层根冠比比较稳定,变化在0.32—0.40之间。这也说明混交林内水分和养分资源比较稳定。
3.3 尾巨桉与红锥混交对林地土壤理化性质的影响
树种或林分的不同使得林下植被种类组成和多样性发生改变,必然导致生境条件的变化,进而引起土壤理化性质的差异。许多研究表明,与人工纯林相比,混交林可以改变凋落物的数量和质量,提高凋落物分解速率,增加养分的归还量,因此,树种混交能有效的维持和改善土壤质量(Forrester et al.,2005;Huang et al.,2014)。本研究发现,尾巨桉与红锥混交对林地土壤理化性质有显著影响。首先,MEC 0—100 cm土层的土壤pH值显著高于PEU和 PCH,说明尾巨桉与红锥混交可以缓解 PEU和PCH引起的土壤酸化;林下植被去除试验表明,林下植物功能群与土壤 pH值密切相关,去除木本植物功能群时土壤pH值的降幅最大(周晓果,2016),MEC林下的木本植物功能群最丰富,因而有利于提高土壤的 pH值。其次,尾巨桉与红锥混交降低土壤容重,有利于促进林木生长;对PEU和PCH根系生物量研究发现,尾巨桉为深根性树种,其主根深至土壤1.5 m以上,而红锥为㳀根性树种,其主根生长在1 m土层内,根系主要分布在0—70 cm土层,桉树与红锥混交显著提高根系的数量,因而能够有效的改善土壤结构,降低土壤容重。第三,尾巨桉与红锥混交提高土壤有机碳和全钾含量,其土壤有机碳显著高于PEU,而土壤全钾含量显著高于PCH;虽然3种林分的土壤全磷含量无显著差异,但 MEC显著提高土壤有效磷的含量,其土壤有效磷含量显著高于两种纯林;研究表明,混交林不仅能充分利用地上地下空间及资源(Sagar et al.,2003;汪殿蓓等,2001),还能提高凋落物的数量与质量,促进土壤养分的归还量,提高养分的有效性和利用率(耿玉清等,1999;李茂金等,2012)。树种混交增强物种丰富度,同时会改变凋落物的数量和质量,并通过光照、温湿度等环境因素影响凋落物的分解速率,增加养分的归还量(Forrester et al.,2005;Huang et al.,2014);植物根呼吸及养分元素的吸收存在种间差异,促使土壤动物群落发生改变,进而增加凋落物分解,大量的磷元素归还于土壤(董玉梁等,2018);混交林林下植被物种多样性较高,林下植被生物量、盖度高,凋落物不断积累,从而丰富了土壤的有机质(王慧敏等,2013)。
有研究发现,土壤理化性质与植物群落物种组成及多样性的关系密切(周晓果,2016)。据同一区域的研究报道,灌木层 Shannon-Wiener指数、Patrick丰富度指数与全氮、水解氮、有效磷、有机质之间呈显著正相关关系,认为水解氮、有效磷、有机质是影响该地区杉木林下植被生长分布的主要因子(Dixon,2003)。在本研究中,影响林下植物群落物种多样性的主控因子为灌木层地下生物量、草本层地上生物量、乔木层地上生物量、全氮、C∶N、N∶P、pH。其中,乔木层地上生物量、灌木层地下生物量、全氮、pH、N∶P与PEU和MEC林下植物群落、灌木层的物种丰富度呈显著的正相关关系。草本地上生物量、C∶N与红锥纯林草本层的物种丰富度呈显著的正相关关系。灌木层、草本层及林下植物群落Shannon-Wiener指数、Simpson指数和均匀度指数主要受灌木层地下生物量、草本层地上生物量的显著影响。这可能与生物多样性和生物量的关系决定的(温远光等,2008)。
4 结论
在生态营林显著减少林下植被抚育强度和频率的情况下,“双龙出海”和宽窄行配置模式经营的PEU降低物种多样性,导致林下灌木层阳性大型木质藤本的强势生长,地上生物量显著增加,降低尾巨桉生物量,并导致土壤容重增加、土壤 pH显著下降、有机质和有效磷的显著减少。在生态营林方式下,PCH降低灌木层多样性、增加草本层多样性和生物量,并具有增加土壤含水量、降低土壤容重、增加土壤有机碳的作用,但显著降低土壤全钾含量、早期林分生长缓慢,生物量低。而在“双龙出海+珍贵树种混交”的生态营林方式下,尾巨桉-红锥混交林显著提高了林下植被的物种丰富度,在较高水平上维持了灌草生物量和地上地下生物量的相对平衡,并有改善土壤结构,缓解土壤酸化,降低土壤容重,提高土壤有机碳、全钾、有效磷含量等作用,有效提升混交树种和混交林分的生物量。因此,建议在南亚热带地区推广应用此营林方式。
猜你喜欢
青年文学家(2021年16期)2021-08-05
少儿美术(2020年8期)2020-11-06
防护林科技(2020年6期)2020-08-12
诗潮(2019年10期)2019-11-19
绿色科技(2019年6期)2019-04-12
绿色科技(2019年6期)2019-04-12
中央民族大学学报(自然科学版)(2018年4期)2018-03-19
诗潮(2017年2期)2017-03-16
绿色科技(2014年4期)2015-01-06
安徽农学通报(2014年9期)2014-06-23
