
棘头梅童鱼肝型脂肪酸结合蛋白基因克隆及组织表达分析
2022-11-18 01:03欧阳文杰宋炜陈佳桂福坤程家骅谢正丽冯广朋
水产科技情报 2022年6期
欧阳文杰 宋炜 陈佳 桂福坤 程家骅 谢正丽 冯广朋
(1 浙江海洋大学,国家海洋设施养殖工程技术研究中心,浙江舟山 316022; 2 中国水产科学研究院东海水产研究所,农业农村部远洋与极地渔业创新重点实验室,上海 200090; 3 福建福鼎海鸥水产食品有限公司,大黄鱼育种国家重点实验室,福建宁德 352103; 4 中国水产科学研究院渔业机械仪器研究所,农业农村部渔业装备与工程技术重点实验室,上海 200092)
脂肪酸结合蛋白(fatty acid binding proteins,FABPs)属于脂结合蛋白超家族成员,是一类分子量较小而对脂肪酸有高亲和力的可溶性蛋白质,广泛分布于哺乳动物的肠道黏膜、肝脏、肾脏、心脏、肌肉、脑、脂肪组织等组织细胞中,其功能主要是参与细胞对脂肪酸的摄取、细胞内脂肪酸的转运及代谢调节,也参与调节细胞生长和分化、机体免疫等[1-4]。自1972年首次发现该蛋白以来,根据被分离出的组织及其主要结构,已鉴定出18种不同的脂肪酸结合蛋白[5-8]。不同类型的FABP基因序列之间具有较高的同源性,目前普遍认为它们是由鱼类和哺乳动物分化之前的一个共同祖先进化而来的[9]。肝型脂肪酸结合蛋白(liver fatty acid binding protein,L-FABP)是脂肪酸结合蛋白家族的重要成员,对长链脂肪酸(LCFAs)有较高的亲和力,可以介导脂肪酸由细胞膜转运到氧化和合成的部位,在LCFAs的代谢中发挥着至关重要的作用[10]。目前已在斑马鱼(Daniorerio)[11]、绿河鲀(Tetraodonnigroviridis)[12]、草鱼(Ctenopharyngodonidellus)[13]、虹鳟(Oncorhynchusmykiss)[14]和雀鳝(Lepisosteusoculatus)[15]等少数鱼类中克隆得到L-FABP基因序列。研究表明,斑马鱼和绿河鲀的L-FABP基因主要在肠道中表达,草鱼和虹鳟的L-FABP基因主要在肝脏中有表达,而雀鳝的L-FABP基因在心脏中的相对表达量最高。
棘头梅童鱼(Collichthyslucidus)隶属于鲈形目(Perciformes)、石首鱼科(Sciaenidae)、梅童鱼属(Collichthys)。该鱼适温、适盐范围广,广泛分布于我国近海和河口地区,是重要的小型底栖经济鱼类[16-17]。棘头梅童鱼肉质细嫩,味道鲜美,脂肪酸种类多样,富含鲜味氨基酸,必需氨基酸组成合理,具有较高的营养价值[18]。目前,有关棘头梅童鱼的研究主要集中在形态分类[19-20]、早期发育[21-22]、遗传结构[23-24]和生化成分[25]等方面,尚未见关于该鱼脂类营养需求及其调控方面的报道。本研究克隆得到了棘头梅童鱼L-FABP基因的cDNA序列,进行了相关生物信息学分析,并检测了其在不同组织中的表达特异性,以期为进一步研究L-FABP基因在棘头梅童鱼营养调控、脂肪酸代谢和免疫应答等方面的功能提供基础资料。

表1 棘头梅童鱼L-FABP基因克隆和荧光定量PCR检测所用引物
1 材料和方法
1.1 试验材料
试验用棘头梅童鱼为2021年9月9日从闽东海域采集的野生棘头梅童鱼,共16尾,平均体质量为(20.36±11.98)g,平均体长为(9.91±2.31)cm。解剖试验鱼并取其不同组织(胃、鳃、肝脏、肠道、眼、大脑、心脏、肌肉、脾脏、头肾和中肾),放入液氮中速冻,置于-80 ℃冰箱中保存,用于RNA提取。
1.2 RNA提取及cDNA合成
参照Trizol试剂(Beyotime)说明书提取各组织样本的总RNA。用微量分光光度计(BioTeke ND5000)检测提取RNA样品的浓度和纯度,并用1%琼脂糖凝胶电泳检测其完整性。使用ReverTra Ace qPCR RT Kit试剂盒(TOYOBO)合成互补DNA(cDNA),置于-20 ℃冰箱保存,用于后续基因克隆和荧光定量(qRT-PCR)分析。
1.3 L-FABP基因克隆
以Gan等[17]全基因组测序组装数据为基础,在Ensembl基因组数据库中获取墨西哥脂鲤(Astyanaxmexicanus)、斑马鱼、尼罗罗非鱼(Oreochromisniloticus)、绿河鲀、剑尾鱼(Xiphophorushelleri)和大黄鱼(Larimichthyscrocea)的L-FABP基因序列,进行BLAST比对(10-10),获得棘头梅童鱼L-FABP基因的含开放阅读框(open reading frame,ORF)的cDNA序列。在L-FABPcDNA序列上、下游设计验证引物(见表1),扩增产物送上海杰李生物技术有限公司进行测序。
1.4 生物信息学分析
用Vector NTI 11.5.1软件预测棘头梅童鱼L-FABP基因的ORF,推测其编码的氨基酸序列;使用在线工具ProtParam(http://web.expasy.org/protparam/)计算氨基酸分子量和等电点等理化性质;利用InterProScan在线网站(https://www.ebi.ac.uk/interpro/search/sequence-search)对蛋白结构域进行预测;利用在线工具UCL(http://bioinf.cs.ucl.ac.uk/psipred/)和SWISS-MODEL(https://swissmodel.expasy.org/interactive)预测蛋白质的二级和三级结构;用SignalP-5.0 Server在线网站(http://www.cbs.dtu.dk/services/SignalP/)预测L-FABP的信号肽;采用Clustal X软件对棘头梅童鱼L-FABP基因的氨基酸序列与其他物种进行多序列比对,并在MEGA 7软件中,使用Maximum Likelihood method(最大似然估计法)的LG+G模型构建系统发育树,设置Bootstrap为1 000。

注:方框内为起始密码子;*为终止密码子;下划线为脂质/胞质性脂肪酸结合结构域;灰色阴影为多聚腺苷酸化信号。
1.5 组织表达分析
L-FABP基因的特异性引物使用Primer Premier 5软件及NCBI数据库设计,β-actin作为内参基因(见表1)。使用cDNA为模板,选择TB Green®PremixExTaqTM(TaKaRa)作为定量反应试剂,根据Bio-Rad CFX96TM型荧光定量PCR仪进行qRT-PCR反应。qRT-PCR反应体系为20 μL:10 μL TB GreenPremixExTaq,0.4 μL ROX Reference Dye(50×),6.8 μL ddH2O,上、下游引物各0.4 μL,2 μL 10倍稀释后的cDNA模板。反应程序为:95 ℃预变性30 s;95 ℃变性5 s,55 ℃退火30 s,72 ℃延伸1 min,共40个循环;最后60~95 ℃获得熔解曲线并确认扩增引物的特异性。
1.6 数据分析
使用2-ΔΔCT方法测定不同组织中L-FABP基因的相对表达量。试验数据采用Prism 8软件进行单因素方差分析(one-way ANOVA),采用Duncan’s法多重比较检验组间差异的显著性并绘图,设P<0.05为差异显著。
2 结果
2.1 棘头梅童鱼L-FABP基因序列
克隆得到棘头梅童鱼L-FABP基因的含ORF的cDNA序列为475 bp,其中5’-UTR(非翻译区)为51 bp,3’-UTR为40 bp,在3’-UTR区域能找到明显的多腺苷酸化信号(PAS)AATAAA序列。起始密码子为ATG,终止密码子为TAA,ORF为384 bp,编码127个氨基酸,分子量为13.90 kDa,等电点(pI)值为5.59,不稳定系数(II)为22.51,判断为稳定蛋白,脂溶指数(AI)为87.32,总平均亲水性(GRAVY)为-0.346。通过蛋白结构域预测分析,找到编码蛋白在第3~126位点有脂质/胞质性脂肪酸结合结构域(见图1)。根据UCL和SWISS-MODEL预测结果,棘头梅童鱼L-FABP基因的氨基酸结构由10个β-折叠链和2个α-螺旋链构成(见图2)。SignalP-5.0 Server的结果表明,在棘头梅童鱼L-FABP基因氨基酸序列中未发现信号肽(见图3)。
2.2 系统进化分析
将棘头梅童鱼L-FABP基因氨基酸序列提交至NCBI,通过blastp比对氨基酸同源性。结果表明,棘头梅童鱼L-FABP基因氨基酸序列与大黄鱼一致性最高,相似性为96.06%。与条纹狼鲈(Moronesaxatilis)、赤锯鳞鱼(Myripristismurdjan)、白梭吻鲈(Sanderlucioperca)等物种的一致性为92.13%(见表2)。使用Clustal X软件进行多序列比对,结果表明,棘头梅童鱼L-FABP基因氨基酸序列与其他物种相比,具有41个绝对保守位点(同一性100%),其中赖氨酸(Lys)、甘氨酸(Gly)及苏氨酸(Thr)出现的频率最高(见图4)。使用MEGA 7软件绘制系统发育进化树,结果发现,棘头梅童鱼L-FABP基因可与其他物种L-FABP基因聚类,与大黄鱼有较高的同源性,置信度为81,而与绿河鲀的同源性较低(见图5)。

2.3 L-FABP基因在不同组织的表达水平
L-FABP基因在棘头梅童鱼肝脏、肠道、眼、大脑、心脏、肌肉、胃、鳃、脾脏、头肾和中肾共11个组织中均有表达(见图6),且存在组织表达差异性。棘头梅童鱼肝脏中的L-FABP基因相对表达量显著高于其他组织中的(P<0.05),肠道、肌肉组织次之,头肾、心脏、眼睛、鳃、胃和大脑中的L-FABP基因相对表达量较低,中肾和脾脏中的L-FABP基因相对表达量最低。
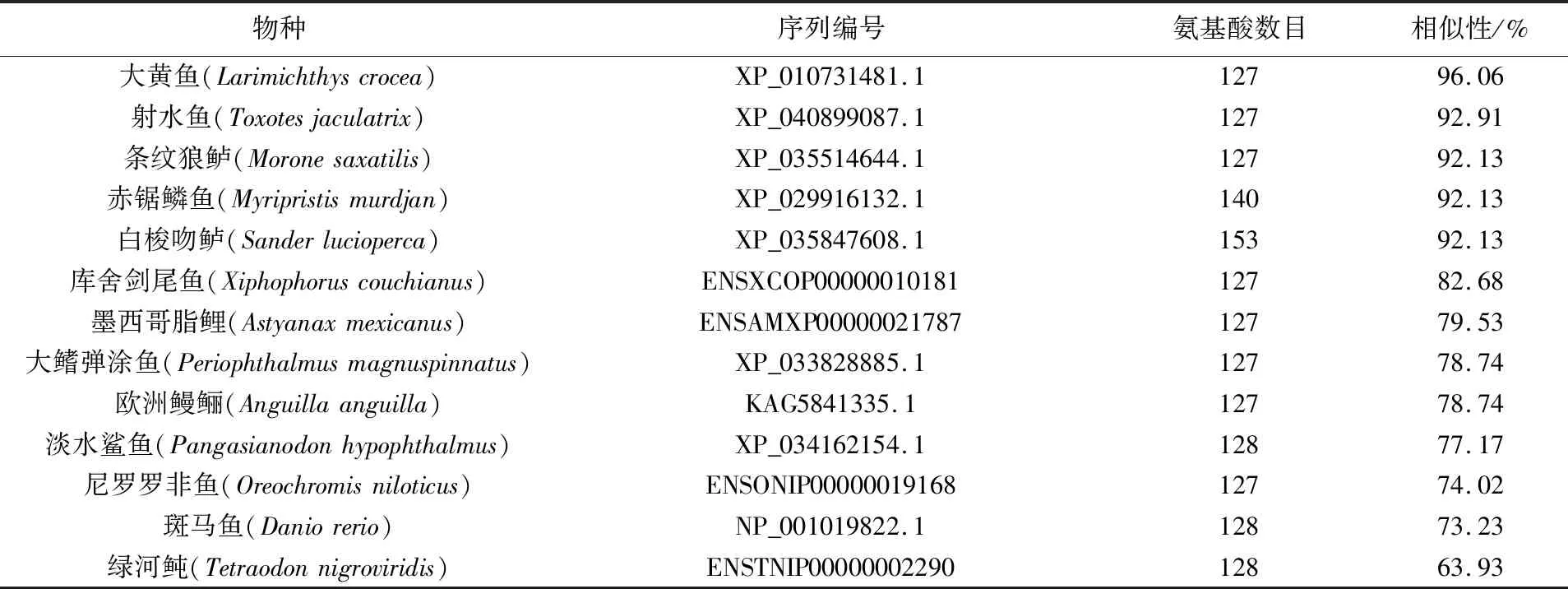
表2 棘头梅童鱼与其他物种同源L-FABP基因的相似性
3 讨论
本研究成功克隆出棘头梅童鱼L-FABP基因的cDNA序列,并对其氨基酸序列进行了分析,试验发现,棘头梅童鱼L-FABP基因含有脂质/胞质性脂肪酸结合结构域,与鱼类和其他水生无脊椎动物FABPs中所发现的保守结构域一致[26]。预测得到的蛋白质结构为10条反向平行的β-折叠链和2条短的α-螺旋链,符合传统FABPs基因家族成员的蛋白质结构特点[27]。氨基酸序列比对结果表明,棘头梅童鱼L-FABP基因的氨基酸序列与其他鱼类相比,具有41个绝对保守位点,其中Lys、Gly、Thr出现的频率最高,推测其可能与维持L-FABP基因在棘头梅童鱼进化过程中的生物学功能稳定有关。利用L-FABP进行的系统发育分析结果显示,棘头梅童鱼与大黄鱼(石首鱼科)同源性很高,与淡水鲨鱼、斑马鱼及尼罗罗非鱼同源性仅为75%左右,符合传统分类地位。
FABPs基因家族成员蛋白对脂肪酸有很高的亲和力,在脂肪酸代谢过程中起着重要作用。其主要功能是在脂肪酸进入细胞后,将其转运到不同的位点进行代谢或者储存,同时也参与调节脂肪酸氧化和代谢,并通过参与脂类合成从而影响细胞生长分化。因此,FABPs基因家族成员蛋白在各生物的不同组织,如肝脏、肠道、肌肉和心脏等广泛分布。本研究主要针对棘头梅童鱼FABPs基因家族中的L-FABP基因在不同组织中的表达进行研究。研究发现,L-FABP基因广泛分布于多种组织中,与L-FABP基因在卵形鲳鲹(Trachinotusovatus)组织中的表达模式相似[28]。肝型脂肪酸结合蛋白(L-FABP)对长链脂肪酸(LCFA)在细胞内的转运具有重要作用,且具有信号转导分子的功能,是机体能量代谢的重要环节[29]。在哺乳动物中,L-FABP基因主要分布于肝脏、肠道、肾脏和胰腺中[30]。而L-FABP基因在不同鱼类不同组织的分布差异较大,草鱼的L-FABP基因在肝脏中相对表达量最高,其次是后肠、脾脏和后肾,在前肠、中肠、心脏、头肾、肌肉和鳃中的相对表达量较低[13]。虹鳟[14]和矛尾复虾虎鱼(Synechogobiushasta)[26]的L-FABP基因也是在肝脏中相对表达量最高。斑马鱼和绿河鲀的L-FABP基因主要在肠道中富集[11-12,31]。斑点雀鳝的L-FABP基因在心脏组织中相对表达量最高[15]。本研究中,L-FABP基因在棘头梅童鱼不同组织中的表达也存在差异,在肝脏中的相对表达量显著高于其他组织中的(P<0.05),说明肝脏是棘头梅童鱼脂质合成和脂肪水解的核心部位。基因的表达功能与其在组织中的特异性表达紧密相关。脂肪转运的内源性途径(endogenous pathway)就是通过肝脏将合成的脂肪转运至机体其他组织贮藏或利用的过程,因此,肝脏是脂肪代谢的重要场所。根据棘头梅童鱼L-FABP基因在肝脏中的高相对表达量可以推断,肝脏参与合成大量的脂肪,为维持其稳态,高含量的脂肪酸会促进L-FABP基因进行转录翻译,从而将过多的脂肪酸运出肝脏,或转运至线粒体中氧化分解。因此推测,L-FABP基因在参与棘头梅童鱼脂肪酸转运和吸收,以及肝脏对脂肪酸的调节和代谢的过程中具有相关作用,该基因在肝脏中具体的生物学功能值得进一步探究。L-FABP基因在棘头梅童鱼肌肉中的相对表达量也较高,推测原因,可能是棘头梅童鱼在运动过程中的肌肉舒张和收缩需要消耗大量的能量,从而促进L-FABP蛋白转运脂肪酸至线粒体进行氧化分解,由此为机体供能;也有可能与本研究采集的棘头梅童鱼正处于性腺发育成熟期有关,说明此阶段棘头梅童鱼肌肉中脂代谢活动旺盛。


注:标注字母不同表示组间差异显著(P<0.05),字母相同表示组间差异不显著(P>0.05)。
4 结论
本研究克隆得到了棘头梅童鱼L-FABP基因的含ORF的cDNA序列,通过多序列比对和系统发育分析,证明该基因与其他鱼类的肝型脂肪酸结合蛋白基因高度相似。L-FABP基因广泛分布于棘头梅童鱼不同组织中,猜想L-FABP基因在棘头梅童鱼不同组织中均发挥作用,且在脂代谢途径中发挥不同的功能作用,这为后期功能验证创造了条件。本研究结果有助于进一步认识鱼类脂肪酸结合蛋白,也为棘头梅童鱼营养及免疫调控分子机制研究提供了基础资料。
猜你喜欢
环球时报(2022-09-20)2022-09-20
湖北农业科学(2022年11期)2022-07-18
中国生殖健康(2020年5期)2021-01-18
今日农业(2020年24期)2020-12-15
生物信息学(2020年1期)2020-05-16
中成药(2018年12期)2018-12-29
中国生殖健康(2018年5期)2018-11-06
天津科技大学学报(2016年1期)2016-02-28
小资CHIC!ELEGANCE(2015年15期)2015-09-01
中国洗涤用品工业(2015年8期)2015-02-28
