
金黄色葡萄球菌表面蛋白及致病性相关研究进展
2022-12-15 09:58王晓宁景双艳
世界最新医学信息文摘 2022年69期
王晓宁,景双艳
(甘肃中医药大学,甘肃 兰州 730000)
1 金黄色葡萄球菌
金黄色葡萄球菌(Staphylococcus aureus,S.aureus)在我们生活中无处不在,它可以引起食物中毒、机体的局部皮肤感染,严重时会导致肺炎、败血症甚至死亡。
1.1 金黄色葡萄球菌的生物学特征
金黄色葡萄球菌是一个在显微镜下观察呈葡萄串状、球型的革兰阳性菌,无鞭毛,无芽胞,大多数无荚膜。
对于生长条件,金黄色葡萄球菌的营养要求不高,需氧或兼性厌氧,在普通的MH(Mueller-Hinton agar plate)平板上就能生长,最适温度是37℃,最适PH为7.4,科研实验中一般将其在血平板和胰蛋白胨大豆琼脂培养基(Tryptic Soy Agar,TSA培养基)平板上进行培养,在以上平板上的金黄色葡萄球菌菌落形状呈圆形凸起,表面光泽,在血平板上会出现完全透明的β溶血环,这也是鉴定金黄色葡萄球菌的标志之一。液体培养基的选择一般肉汤(LB)培养基和胰酪大豆胨液体(TSB)培养基,37℃过夜培养后,发现金黄色葡萄球菌呈均匀混浊生长。
金黄色葡萄球菌的生化反应中[1]:触酶(+)、葡萄糖(+)、麦芽糖(+)、蔗糖(+)、7.5%NaCl甘露醇(+)、棉子糖(-)、水杨苷(-)、明胶(+)、血浆凝固酶(+)、DNA酶(+)、七叶苷(-)、A蛋白和凝集因子(+)。
1.2 金黄色葡萄球菌的流行病学
金黄色葡萄球菌是一种人兽共患病原菌,它存在自然界各个地方,比如土壤、空气和水中;在人的皮肤、鼻咽部等也会发现金黄色葡萄球菌的存在。据文献查阅,健康人携带金黄色葡萄球菌的比率为20%-30%,上呼吸道感染患者的鼻腔中检测发现带菌率高达83%,化脓性感染患者的感染部位也有发现金黄色葡萄球菌的存在。在美国的疾病控制中心报道,由金黄色葡萄球菌引发的感染仅次于大肠杆菌,占第二位[2];在我国由金黄色葡萄球菌引发的食物中毒事件占25%左右,位居第四位[3]。
由于金黄色葡萄球菌自身的一些特征从而形成了如下几个流行病学特点:第一,季节分布,在春夏季节, 由于气候呈现温热潮湿,在这种条件下,更加利于金黄色葡萄球菌的生长繁殖,导致春夏季节成为金黄色葡萄球菌感染的高发时间。第二,引发食物中毒,由于金黄色葡萄球菌存在自然界的各个角落,就连食物表面也不例外,比如奶、肉、蛋、鱼及其制品,这些食物被金黄色葡萄球菌感染后,如果没有进行灭菌处理则会导致细菌性食物中毒,严重者会引起机体紊乱,甚至引起死亡。第三,人兽共患,在上呼吸道感染患者的鼻腔中也发现金黄色葡萄球菌的存在,这应引起我们的高度重视。
近年来,随着大量抗生素应用于临床治疗,金黄色葡萄球菌出现了大量的耐药菌株,其中最为严重的是甲氧西林耐药金黄色葡萄球菌(MRSA),MRSA又分为医院获得性MRSA(HA-MRSA)和社区获得性MRSA(CA-MRSA),两者的区别是患者是否在门诊或入院48h之内分离得到的MRSA菌株。CAMRSA通常只对β-内酰胺类抗生素耐药,但HA-MRSA可对多种药物耐药。
2 金黄色葡萄球菌的耐药机制
目前对于β-内酰胺类抗生素耐药的耐药性机制主要分为两种:一种是产生β-内酰胺酶,它可以在抗生素作用之前使β-内酰胺抗生素失活或降解。另一种是青霉素结合蛋白2a(PBP2a)的产生,该蛋白与β-内酰胺类抗生素的亲和力低,从而绕开了抗生素的活性[4]。CA-MRSA耐药机制主要是由于青霉素结合蛋白(PBPs) 发生变化所致,它产生的PBP2a可以导致与β-内酰胺类抗生素(所有头孢菌素和半合成青霉素)的亲和力下降。因此PBPs会受到抑制而不能发挥作用,但PBP2a仍能够发挥PBPs所有的功能,并参与细菌细胞壁的合成,使细菌得以生存,从而表现为对β-内酰胺类抗生素耐药[5]。PBP2a由mecA基因编码,而MRSA的种属可以根据mecA基因类型分类[6]。
3 金黄色葡萄球菌的表面蛋白
当前金黄色葡萄球菌需要找到新的抗生素,需要寻找到对于细菌新的靶点,金黄色葡萄球菌表面蛋白正因为与宿主有着特异性粘附能力,成为对抗金黄色葡萄球菌感染疫苗开发的新热点。目前金黄色葡萄球菌表面蛋白主要为细胞外结合基质蛋白(extracellularmatrix-binding proteins,ECMBP),细胞外基质(extracellular-matrix,ECM)在脊椎动物的组织和器官中都会发现,通过大量实验研究发现,它可以支持、增殖及调节基因表达[7]。它们与细胞相互作用,又参与生理和病理的过程,例如帮助机体伤口的愈合以及癌细胞的转移。细胞表面也存在大量的ECM受体,它们被称为整合素(integrins)家族[8-9],病原性细菌表面的ECM受体被称为黏附素或者是可以识别粘附基质的细胞表面成分(Microbial Surface Components Recognizing Adhesive Matrix Molecule, MSCRAMM),它们被认为是细菌感染的必要条件之一[10-11],因为如果没有这些成分,细菌无法定植及繁殖,更不能释放外毒素(exotoxin),就不会引起感染。不仅如此,当细菌完全被机体ECM 所包被后,机体的免疫系统不会将其识别,从而进一步引发感染。
目前金黄色葡萄球菌中有5个表面蛋白的分子结构已研究清楚(图1),分别是葡萄球菌蛋白A(SpA)、胶原蛋白结合(Cna)蛋白、纤连蛋白结合(FnBP)蛋白、纤维蛋白结合(ClfA)蛋白和弹性蛋白结合蛋白(Ebps)。其中SpA不属于MSCRAMM,它可以结合IgG的Fc区域来抑制吞噬作用[12],它还可以介导血管假性血友病因子(von Willebrand)与金黄色葡萄球菌结合。
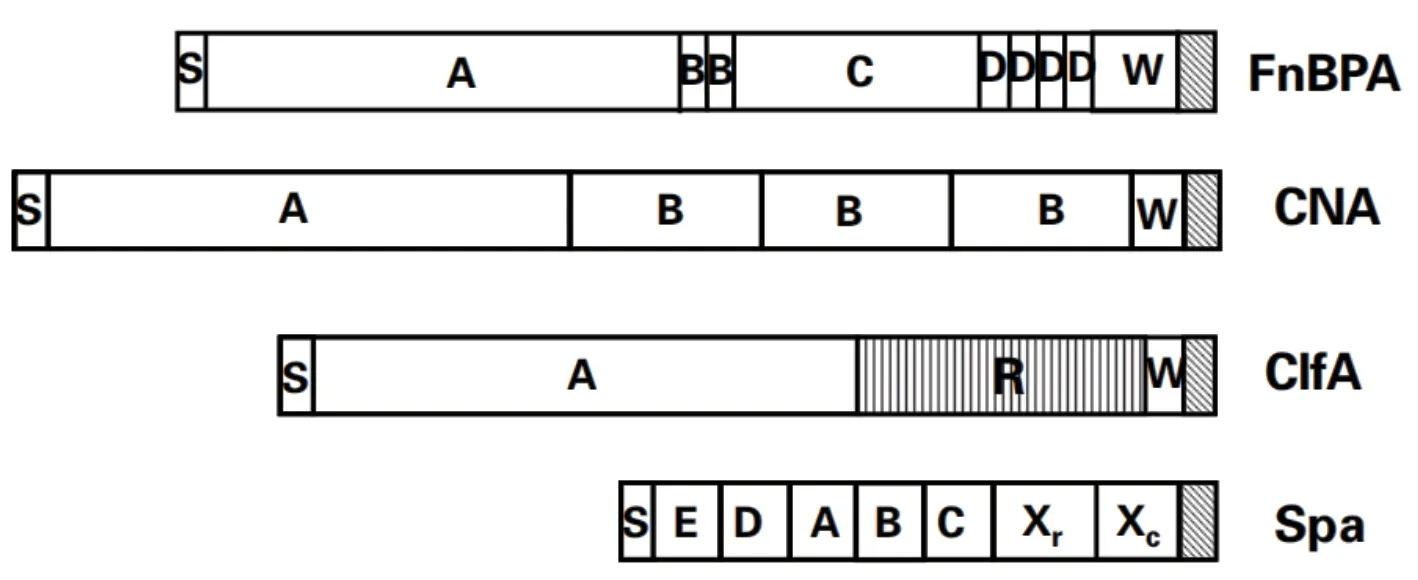
图1 MSCRAMM蛋白的结构[13]
3.1 识别纤连蛋白的FnBP
细菌中的FnBP蛋白可以分为FnBPA和FnBPB,它的相对分子质量约为110 000,它的结构由七部分组成,分别是处于N端的信号肽(S)区,之后为非重复序列的伸展区(A),接下来是含有2个重复单元的(B)区域,C区之后有4个重复的D区,它们是由38个氨基酸残基模块构成的结构域,紧接着是细胞壁延展区域(W),最后是相对保守的LPXTG基序。
3.2 识别胶原蛋白的Can
细菌中的Can蛋白使细菌粘附至胶原蛋白、组织及软骨上。它的结构由五部分组成,分别是信号肽(S)区,紧跟着分子质量为55 000的A区,之后为3个重复序列的B区,接下来是细胞壁延展区域(D)和LPXTG基序。其中B区经研究发现,这段重复序列对于Can蛋白为非必需的。在A子区域发现两个反向平行的β折叠以及两个α螺旋结构,它们共同组成“果冻状”折叠结构,此区域可以抑制细菌与胶原蛋白结合[14-15]。
3.3 识别纤维蛋白原的ClfA
细菌中的ClfA蛋白决定着金黄色葡萄球菌与含纤维蛋白基质作用[16],ClfA蛋白出现在细菌生长的每一个阶段,它与其它表面蛋白结构不同之处在于,含有一个独特打R区域,这个区域由Asp-Ser的二肽重复组成,前后分别是A区和细胞壁延展W区,此区域可以促使A区在细菌表面与配基发生作用[17]。
4 金黄色葡萄球菌表面蛋白Ebh
金黄色葡萄球菌表面蛋白具有很多功能,它可以使细菌粘附和定植,进一步形成生物被膜,一方面可以躲开免疫系统和免疫细胞的“追杀”,另一方面又阻碍了抗生素的杀菌作用[18]。最终导致机体免疫能力下降,发生感染从而致病。Kuroda等利用生物信息学技术在金黄色葡萄球菌细胞表面发现了一种新的蛋白—Ebh蛋白,这是一种功能未知的细胞外基质结合蛋白,定位于C末端[19]。有研究发现,在已确诊的金黄色葡萄球菌感染的患者血清中出现了抗Ebh抗体,定位研究显示Ebh蛋白与细胞包膜有关[20]。深入研究Ebh蛋白对金黄色葡萄球菌生物被膜形成及耐药机制有重大的意义。
主办券商声誉机制对新三板挂牌企业会计信息质量影响的实证研究..................................................................................................................孙 翯 张文松 孟 为(23)
4.1 Ebh蛋白的特性
Ebh蛋白是在金黄色葡萄球菌中发现的一个巨蛋白,约1.1-MDa蛋白,与其他ECMbinding protein具有同源性。它拥有几个结构域,包括一个由126个氨基酸组成的52个不完全重复序列的大中心区域。通过电子显微镜直接观察包含大量重复单元的Ebh蛋白结构,发现Ebh是由高度柔韧的丝状分子组成的;它有高度重复的结构和独特的结构灵活性,即与细胞壁的糖结合,这种结合导致细胞内壁相互交错,紧密联系在一起,使得细胞变得更加坚固[21],同时也避免了在高渗条件下发生质壁分离,促进结构内稳态。Ebh蛋白的结构分析还显示它是一个320nm棒状分子,在模块连接处具有高塑性[22]。
前期课题组首次从医院临床分离的48株金黄色葡萄球菌中发现19株Ebh基因阳性菌株,通过mecA基因检测,鉴定19株Ebh基因阳性菌株中11株为MRSA,8株为MSSA,两者中Ebh基因的携带率差异无统计学意义。利用脉冲凝胶电泳(PFGE,Pulsed Field Gel Electrophoresis)对Ebh基因阳性菌株进行分型,结果显示菌株主要为A型流行株,并对菌株标本来源进行统计,发现大多数Ebh基因阳性菌株来自烧伤病区患者,我们进一步推测Ebh基因可能与其黏附宿主细胞外基质有关[23]。
4.2 Ebh蛋白的致病性研究
4.2.1 金黄色葡萄球菌ArlRS通路调控Ebh基因的表达
金黄色葡萄球菌是非常普遍的一种细菌病原体,引起人类很多严重的疾病。抗生素的频繁使用导致这些疾病更加难以治疗,因此人们开始寻找新的治疗工具。有研究表明,金黄色葡萄球菌在凝集过程中可以结合人基质蛋白形成稳定的团块,这些团块有助于逃避宿主防御并建立感染,他们通过不同的技术来研究金黄色葡萄球菌的凝集机制,最后发现ArlRS双组分调节系统通过调节编码巨型葡萄球菌表面蛋白(GSSP)的Ebh基因的表达来控制凝集;这也在败血症和感染性心内膜炎的兔模型中得到证实,证明ArlRS是毒力的重要调节剂[20]。Fournier和Hooper最初确定并将ArlRS定性为自溶调节因子。然而,Memmi等人进一步研究发现ArlRS在自溶中的调节作用仅限于对甲氧西林敏感的金黄色葡萄球菌(MSSA)菌株,MRSA中的自溶作用受一种独特但尚未确定的机制控制。研究过程中发现Ebh基因参与凝集过程,Ebh基因大小约为33kbp,是金黄色葡萄球菌染色体上最大的开放阅读框,有效地包含了其基因组[24]的1%;Ebh基因编码的Ebh蛋白,也称为GSSP,预计是膜锚定的并以纤维状方式从细胞表面突出[25]。为了进一步研究ArlRS双组分调节系统如何影响细菌凝集,分别在USA300(LAC-WT)菌株中构建了arlRS基因敲除菌株(LAC △arlRS)、arlRS基因及Ebh基因双基因敲除菌株(LAC △arlRS △Ebh),并通过实时荧光定量PCR (qPCR)来检测基因的表达变化情况。研究结果发现,当ArlRS基因敲除后,Ebh基因表达上调近50倍;又通过凝集实验发现,LAC△arlRS不能发生凝集反应,而LAC △arlRS △Ebh能够发生凝集;最后通过光学显微镜观察三种细菌(LAC-WT、LAC △arlRS和LAC △arlRS △Ebh)的形态学结构是否有所差异,结果显示三者都会形成紧密簇,但观察LAC△arlRS菌株发现其细胞分布比较松散,进一步证实了Ebh基因影响了细菌凝集,如何影响凝集的过程还需要进一步的研究。
4.2.2 Ebh蛋白在稳定SCV细胞中的表达
金黄色葡萄球菌发病机制的一个未确定的特征是其持续存在然后疾病复发。这可以通过改用其他生活方式来解释,主要是生物被膜或小菌落变种(SCVs)[26]。金黄色葡萄球菌SCV现已明显与不同的疾病相关,重要的是这些疾病包括慢性感染,如囊性纤维化,慢性鼻窦炎,心内膜炎和慢性骨髓炎患者[27-29]。SCV被定义为产生直径≥1mm的菌落的变体,具有降低的色素沉着和溶血活性[30]。研究发现当金黄色葡萄球菌转化为SCV的时候,会形成一种细胞外基质,它是由蛋白质和DNA组成的,通过qPCR来检测相关基因的表达变化,结果发现Ebh基因发生显著上调,并推测它参与了金黄色葡萄球菌的致病过程[31],另外,由于此基因编码的蛋白过于巨大,推测Ebh蛋白可能会通过干扰肽聚糖的生物合成进一步影响细胞壁的合成[25]。最后,他们从分批培养的细胞和恒化器衍生的稳定SCV细胞制备膜蛋白组分,并对这些细胞进行分析和比较,在这种膜蛋白质组学中,与批量生长细胞相比,Ebh蛋白显示稳定SCV几乎增加了5倍,其他膜蛋白的表达也发生变化,这些蛋白也可能形成稳定的SCV基质的一部分。
5 金黄色葡萄球菌与生物被膜的关系
5.1 生物被膜形成过程
MRSA形成生物被膜的过程(图2)十分复杂,主要分为以下几个部分:①细菌粘附在机体表面;②细菌聚集与增殖;③形成生物被膜;④细菌脱落并转移。当生物被膜内环境不能满足细菌的时候,会通过分泌胞外水解酶,使胞外多糖发生分解,从而成为浮游菌,然后继续粘附、聚集、增殖、形成新的生物被膜,就这样周而复始,生物被膜变得更加坚固。成熟的生物被膜内部会形成一个液体充足的通道,内部的细菌可以经过通道来传递营养物质。在分子学角度上,参与生物被膜形成的分子包括①自溶素A:促使细菌粘附至宿主细胞表面;②多糖胞间黏附素(Polysaccharide Intercellular Adhesion,PIA):可以促进生物被膜成熟,由ica基因编码合成,但ica基因并不是形成生物被膜的必需基因,大量实验研究发现,一些细胞外基质蛋白比如Bap、Embp、FnbpA、FnbpB等,都能够使细菌通过ica非依赖途径形成生物被膜[32];③细胞外DNA(Extracellular DNA,eDNA):eDNA由细菌通过水解胞外多糖产生,它可以促使其它金黄色葡萄球菌进行聚集,从而形成生物被膜。大量实验数据表明金黄色葡萄球菌生物被膜形成还与培养细菌的营养条件、温度和时间有密切联系。

图2 细菌生物被膜形成过程[33]
5.2 群体感应系统对生物被膜形成的影响
5.3 生物被膜的致病性与耐药性及其机制
生物被膜形成之后,机体并不会发生严重的炎症反应,这是因为生物被膜阻碍了细菌与定植部位的接触,这使得细菌与机体发生共存;另一方面,这也阻断了抗生素和机体防御系统对细菌的杀伤,生物被膜内细菌还可以通过脱落继续形成新的生物被膜,导致机体不断发生感染。它的致病机制主要分为2个方面:①生物被膜为细菌定植及感染提供了屏障:生物被膜阻挡了抗体及炎性细胞进入膜内,从而使得无法清除细菌;②损伤免疫细胞:中性粒细胞由于无法进去细胞被膜内部只能聚集在其周围,细胞被膜又阻碍了机体特定信号的传递,最终导致这些中性粒细胞释放细胞毒素、蛋白酶等,使周围组织发生损伤[34]。
6 展望
金黄色葡萄球菌是一种革兰阳性病原体,是医院和社区获得性感染的主要原因,包括致病疾病,如菌血症、感染性心内膜炎和败血症[35]。在金黄色葡萄球菌感染个体的血清样本中检测到抗ebh免疫球蛋白G,表明Ebh蛋白在感染过程中表达。此外,最近的研究已经在筛选人类感染期间在体内表达的基因中鉴定出了这种蛋白[36]。以绿色荧光蛋白为报告源,将Ebh蛋白定位于整个细胞表面,这表明,Ebh蛋白可能通过在细胞壁、细胞壁和细胞质膜之间形成桥梁,以避免在高渗条件下发生质分裂,从而促进结构内稳态[24]。通过电子显微镜观察到很多扭曲的杆状分子,这些分子都具有相同的序列,但分子的形状却不相同,这表明结构域存在广泛的灵活性[21]。金黄色葡萄球菌为什么会产生像Ebh蛋白这样大的蛋白质还不清楚。鉴于Ebh蛋白的巨大规模,人们很容易推测出它可以形成一种细胞粘附的特殊表面结构[24]。
细菌细胞的表面显示出与肽聚糖共价锚定的多种蛋白质,它们具有许多功能,包括与宿主细胞和组织的粘附,非吞噬细胞的侵入,以及先天免疫反应的逃避。基于结构和功能分析,蛋白质已被分类为不同的类别。许多表面蛋白是多功能的,细胞壁锚定的蛋白质在共生状态期间和侵入性感染期间执行支持存活和增殖的基本功能;细胞壁锚定蛋白结合脱皮上皮细胞的能力在定植期间是重要的,并且与纤维蛋白原的结合在发病机理中具有特别重要[37]。
现在仍然有很多金黄色葡萄球菌表面蛋白的功能尚未发现,需要我们去发掘及深度研究,为解决细菌耐药性提供新的靶点,更好应用于临床治疗。对于表面蛋白的研究不能仅仅针对于其结构,可以在分子水平上进行研究。目前随着科学技术的飞速发展,越来越多的新技术应用于科研之路上,其中包括对细菌进行全基因组测序、基因敲除、转录组测序、蛋白组学测序等。基因组测序主要分为一代测序、二代测序和三代测序;基因敲除主要针对某个基因功能的研究;转录组测序主要针对细胞转录后的RNA进行分析研究;蛋白组测序则是针对转录翻译后的蛋白质进行机制及通路方面的研究。这些技术将为我们研究新的表面蛋白奠定基础。
猜你喜欢
九江学院学报(自然科学版)(2022年2期)2022-07-02
草地学报(2022年3期)2022-03-28
今日农业(2021年11期)2021-11-27
食品安全导刊(2021年20期)2021-08-30
中国食用菌(2020年11期)2021-01-18
科研成果与传播(2019年3期)2019-09-10
创新作文(1-2年级)(2018年2期)2018-09-13
学习报·教育研究(2017年33期)2017-08-31
食品工业科技(2014年23期)2014-03-11
食品工业科技(2014年23期)2014-03-11
