
增温对青藏高原高寒沼泽草甸不同时期土壤酶活性的影响
2023-02-22 12:25高钰雍少宁王俊峰
生态科学 2023年1期
高钰, 雍少宁, 王俊峰
增温对青藏高原高寒沼泽草甸不同时期土壤酶活性的影响
高钰1, 2, 雍少宁3, 王俊峰1,*
1. 中国科学院西北生态环境资源研究院, 冻土工程国家重点实验室, 兰州 730000 2. 中国科学院大学, 北京 100049 3. 甘肃省交通运输厅, 兰州 730000
为研究青藏高原多年冻土区土壤酶活性对气候变暖的响应特征, 选择北麓河地区高寒沼泽草甸为研究对象, 利用开顶式生长室(OTC)进行模拟增温试验, 在7月份、12月份分别对增温和对照样地土壤进行分层取样, 并对不同深度土壤脲酶、蔗糖酶、过氧化氢酶和磷酸酶的活性进行分析测定。结果表明: 在夏季7月份, 增温作用下近地表月平均气温升高4.55 ℃, 不同深度的土壤脲酶、蔗糖酶、过氧化氢酶和磷酸酶活性均有降低趋势, 但未达到显著水平(>0.05)。土壤蔗糖酶活性与土壤水分和根系生物量呈显著相关关系; 在冬季12月份, 增温作用下近地表月平均温度升高6.64 ℃, 不同深度的土壤脲酶、蔗糖酶、过氧化氢酶和磷酸酶活性均有所提高, 且在20—50 cm深度的土壤蔗糖酶和磷酸酶活性和40—50 cm深度的土壤脲酶和过氧化氢酶活性变化均达到显著水平(<0.05)。四种土壤酶活性与土壤温度和水分均呈显著相关关系。在深度变化上, 夏季增温样地四种土壤酶活性总体随土层深度增加而降低, 而冬季随土层深度增加而显著升高(<0.05)。增温处理下, 不同土层深度冬季四种土壤酶的活性总体均大于夏季, 且在40—50 cm深度上差异显著(<0.05)。增温对高寒沼泽草甸土壤酶活性的影响冬季大于夏季。夏季土壤酶活性受土壤水分、根系生物量降低等因素的影响, 对增温的响应不显著。冬季土壤酶活性受不同深度土壤水分、温度的影响, 在土壤温度、水分含量均较高的土层表现出较高的活性。
土壤酶活性; 增温; 高寒沼泽草甸; 青藏高原
0 前言
土壤酶是一种具有生物催化能力的高分子活性物质, 参与土壤中各种生物化学过程, 在土壤系统有机质分解和养分循环过程中发挥着重要作用[1]。土壤酶活性受土壤微生物、土壤理化性质、季节变化、土壤类型及植物多样性等多方面影响[2]。温度作为影响土壤酶活性的重要因子, 一方面, 通过在一定温度阈值内提高酶动力学常数, 来提高土壤酶的活性; 另一方面, 通过影响土壤的水热条件、土壤微生物数量、土壤养分含量等因素直接或间接的影响土壤酶的活性[1]。
由于土壤酶活性对增温的复杂响应过程, 目前有关增温对土壤酶活性影响的研究结果存在较大差异。不同土壤深度处土壤酶活性对增温表现出不同的响应特征。郑蔚等[3]对亚热带杉木幼树增温处理发现, 与对照样地相比, 增温使0—20 cm土壤β-葡萄糖苷酶、纤维二糖水解酶、酚氧化酶和过氧化物氧化酶活性提高, 而20—40 cm土壤中除酚氧化酶其余胞外酶的活性均无显著影响或有一定的抑制作用。因为增温增强了表层土壤中微生物和植物的养分竞争, 需提高土壤酶活性来获取微生物所需养分, 深层土壤中, 增温提高了土壤可溶性有机质含量, 给微生物提供更多底物, 进而减少了对土壤酶活性的需求。
同时, 土壤酶活性变化也受季节的影响。有研究表明, 微生物的产酶活性可能在生长期的早期或晚期受到温度的限制, 而这种限制在生长期中期并不连续[4]。李红琴等[5]研究也表明, 转化酶、脲酶、多酚氧化酶、过氧化氢酶、中性磷酸酶和淀粉酶活性的最大值出现在4月和9月, 而非植物生长旺盛的夏季。4月份土壤解冻初期, 微生物和植物的生长刚刚开始, 各种营养物质输入不足, 从而导致酶活性增强。
IPCC预计到21世纪末气温将会升高1.8—4 ℃[6], 高寒地区对气温变化的响应尤为敏感, 其升温速率及幅度远高于低海拔地区[7]。随着全球温度的升高, 高寒生态系统的低温限制效应将会被打破, 土壤温湿度条件也会相应产生改变, 土壤中酶的活性也将随之改变。前述研究工作表明, 不同生态系统类型、不同土层深度及不同季节下各类土壤酶活性对温度变化的响应特征明显且存在显著差异, 而土壤酶活性在土壤养分循环中起着重要作用。因此, 酶活性的变化将直接影响区域生态系统中碳、氮等元素的分解、存储以及其他生态系统过程[8]。受青藏高原冬季气候条件恶劣、监测工作困难等限制, 有关青藏高原高寒草地生态系统酶活性的研究大多集中在生长季, 对冬季酶活性的研究较少。结合前人的研究, 我们假设: (1)青藏高原高寒沼泽草甸土壤酶活性主要受土壤温度和水分变化的影响, 不同季节对增温的响应不同; (2)在空间分布上, 土壤酶活性也会受到不同土层温度和水分的影响而变化。为验证假设, 本研究以青藏高原北麓河地区的高寒沼泽草甸为研究对象, 利用开顶式生长室增温, 对不同深度、不同取样时间土壤酶活性对增温的响应特征进行了系统研究, 并探讨高寒沼泽草甸不同土壤酶活性的温度响应机制, 可为进一步分析全球气候变暖背景下高寒沼泽草甸土壤养分及其生态变化过程提供一定的理论依据和数据支撑。
1 材料和方法
1.1 研究样地概况
研究样地位于青藏高原北麓河高寒沼泽草甸分布区(34 ° 49 ′ 35.1 ″ N, 92 ° 55 ' 37.9 ″ E), 海拔4642 m。该样地处于青藏高原腹地多年冻土区, 地势较为平坦, 地形稍有起伏[9]; 属亚寒带半干旱气候, 年平均气温-3.8 ℃, 年均降水量290 mm, 年均蒸发量1316.9 mm; 冻结期10月至翌年4月[10]; 优势种为藏嵩草()、青藏苔草()和矮火绒草()等, 植被盖度约80%[11]。基于青藏高原气候条件的特殊性, 本研究中季节划分参照范思睿等[12]的方法, 春、夏、秋、冬四季起始日期分别为: 4月24日、7月3日、8月2日和10月11日。
1.2 研究方法
1.2.1 样地设置和环境因子监测
增温采用国际冻原计划(ITEX)模拟增温的方法, 于2010年在研究样地设置开顶式增温系统(Open-Top Chamber System, OTCs), 至2019年已连续增温9年, 增温样地内土壤微生物群落组成、土壤理化过程和植物生理生态过程对增温胁迫已基本适应, 各指标已趋于稳定[13]。OTCs系统顶部开口为60 cm, 框架为正六面体结构, 各边与地面倾角为60°, 六面均为透明有机玻璃板, 透光率达到95%, 考虑青藏高原高寒沼泽草甸植物矮小, 将高度设为0.8 m。研究区内随机设置6组增温样方(OTC)与对照样方(CK), 每个增温样方与相同面积的对照组样方相距5—10 m。OTCs的增温原理是: 在有机玻璃板的阻挡下, 生长室内风速降低, 空气流动减弱, 使热量不易散失, 太阳辐射又对透明有机玻璃板的穿透性较好, 使生长室内部温度升高, 从而在对植物生长环境干扰极小的条件下实现增温[14]。利用自记式空气温湿度传感器(U23-002, ONSET HOBO, 美国)对近地表气温进行监测, 利用土壤温度水分传感器(5TM, Meter Group美国)分别对地下5 cm, 20 cm, 50 cm处的土壤温度和水分含量进行测定, 土壤温度分辨率为0.1 ℃, 体积含水量分辨率为0.08%。样地的土壤温度和水分数据均由数据采集仪(EM50, Decagon美国)自动采集获得, 采集频率为每30分钟采集一次。
1.2.2 土样与植被生物量采集
于2019年7月17日和12月1日, 用土钻分别钻取夏季和冬季增温和对照样地的土壤样品, 采样深度分为0—10 cm、20—30 cm、40—50 cm三层, 分别在6组增温室(OTC)和对照样方(CK)内取样, 即6个重复, 每个重复样地中随机取3个样混合而成。每个样方的样品放入自封袋中密封做好标记, 随后用便捷式冰箱带回实验室风干, 去除植物根系和石块, 过100目筛用于土壤酶活性的测定。
植被根系生物量的测定采用直接收获法[15], 将植物地上部分剪取后, 用直径7 cm的根钻钻取根土样品, 每隔10 cm取一次, 取样深度为地下50 cm, 后用60目筛网在流水中冲洗掉泥土, 并根据表面和断面的颜色筛选出各层活根, 将植物样品装入信封中带回实验室, 烘箱内105 ℃杀青后, 80 ℃烘至恒重, 称干物质重量。
1.2.3 土壤酶活性的测定
土壤脲酶和土壤蔗糖酶分别在土壤氮素和碳素循环中起着重要作用; 土壤过氧化氢酶活性可以表征土壤腐殖质化强度大小和有机质转化的速度; 土壤磷酸酶活性影响着土壤中有机磷的分解转化[1-2]。对这四种土壤酶活性变化的研究, 能为分析土壤养分的变化提供一定的理论依据。土壤酶测定参照郑蔚[3]、陈康惋[16]使用的方法。取土壤样品1 g, 加入9 mL磷酸缓冲盐溶液, 将制备好的10%的匀浆液离心15分钟, 取上清液测定。在酶标包被板上标准品孔加样50 μL, 待测样品孔中先加样品稀释液40 μL, 再加待测样品10 μL(样品最终稀释度为5倍), 最后将空白孔除外的每孔加酶标试剂100 μL。封板后37 ℃下温育60分钟。弃液体甩干, 加洗涤液, 静置30秒弃去, 重复5次。每孔加显色剂100 μL, 37 ℃避光显色10分钟。加终止液50 μL, 终止反应。15分钟后用酶标仪测定样品的吸光度(即值)。
1.3 数据统计
采用重复测量方差分析(Repeated-analysis ANOVA)检验增温、土壤深度、取样时间以及各因子交互作用对土壤脲酶、蔗糖酶、过氧化氢酶和磷酸酶活性的影响。采用t检验进行假设检验, 分析相同取样时间下, 同一深度增温处理和对照下四种土壤酶活性的差异显著性。采用单因素方差分析(One-way ANOVA)和LSD法检验同一取样时间下, 同种处理不同土壤深度对四种土壤酶活性的影响。采用Pearson相关系数评价土壤温度、土壤水分和夏季根系生物量与土壤酶活性的相关关系。所有统计分析均采用SPSS 21.0完成, 且界定<0.05为显著水平。
2 结果与分析
2.1 OTC增温效能
2019年7月1日—2020年1月1日期间, 研究区域近地表空气温度、土壤温度及土壤含水量变化特征如图1、图2所示。2019年7月, 增温处理后近地表月平均气温提高4.55 ℃; 地下5 cm、20 cm以及50 cm处月平均土壤温度在增温后分别提高1.97 ℃、1.29 ℃和0.85 ℃, 月平均土壤含水量在增温后分别比对照低9.82%、0.35%和4.78%; 2019年12月, 增温处理后近地表月平均气温提高6.64 ℃, 地下5 cm、20 cm以及50 cm处月平均土壤温度在增温后分别提高5.72 ℃、4.66 ℃和3.23 ℃, 月平均土壤含水量在增温后分别比对照样地高4.82%、9.01%、12.34%。
2.2 增温对土壤脲酶活性的影响
在7月份, 增温后土壤脲酶在0—10 cm处活性降低了3%(>0.05), 在20—30 cm、40—50 cm深度处活性增加了67%、3%(>0.05); 在12月份, 增温后土壤脲酶在0—10 cm、20—30 cm、40—50 cm深度处活性分别增加了19%、49%、44%, 其中40—50 cm处显著增加(<0.05)。7月份增温样地土壤脲酶活性随深度增加而显著降低, 12月份则随土层深度增加活性显著增高(<0.05)。此外, 两种处理下7月份脲酶活性均低于12月份, 并且在40—50 cm深度处差异显著(<0.05)(图3)。重复方差分析结果表明, 不同取样时间和土壤深度交互作用对脲酶活性有显著影响(表1)。7月份土壤脲酶活性与土壤温度和水分的相关关系不显著, 12月份土壤脲酶活性与土壤温度呈显著相关关系(表3)。
2.3 增温对土壤蔗糖酶活性的影响
在7月份, 增温后土壤蔗糖酶在0—10 cm、20—30 cm、40—50 cm深度处活性分别降低了32%、33%、20%(>0.05); 在12月份, 增温后土壤蔗糖酶在0—10 cm、20—30 cm、40—50 cm深度活性分别增加了3%、22%、40%, 其中20—30 cm和40—50 cm处显著增加(<0.05)。7月份增温样地土壤蔗糖酶活性随土层深度增加而显著降低, 12月份则随土层深度增加而显著增高(<0.05)。此外, 两种处理下7月份 0—10 cm深度土壤蔗糖酶活性显著高于12月份(<0.05), 增温处理下其余两层7月份土壤蔗糖酶活性低于12月份, 并且在40—50 cm深度处差异显著(<0.05)(图3)。重复方差分析结果表明, 不同取样时间和土壤深度以及不同取样时间和增温处理交互作用对蔗糖酶活性有显著影响(表1)。7月份土壤蔗糖酶活性与土壤水分和夏季根系生物量呈显著相关关系, 12月份土壤蔗糖酶活性与土壤温度和土壤水分呈显著相关关系(表3)。

图1 增温处理下高寒沼泽草甸近地表空气温度的动态变化(2019/7/1—2020/1/1)
Figure 1 Dynamic changes of near-surface air temperature in alpine swamp meadow under warming treatments from July 1, 2019 to January 1, 2020.

注: (a). 对照样地; (b). 增温样地。
Figure 2 Dynamic changes of monthly average soil temperature and soil water content in different soil depths of alpine swamp meadow under warming treatments from July 1, 2019 to January 1, 2020.
2.4 增温对土壤过氧化氢酶活性的影响
在7月份, 增温后土壤过氧化氢酶在0—10 cm深度处活性增加了14%, 在20—30 cm、40—50 cm深度处活性分别降低了6%、1%, 差异均不显著(>0.05); 在12月份, 增温后土壤过氧化氢酶在0—10 cm、20—30 cm、40—50 cm深度活性分别增加了11%、25%、40%, 其中20—30 cm 、40—50 cm处差异达到显著水平(<0.05)。7月份增温样地土壤过氧化氢酶随土层深度增加活性显著降低, 12月份则随土层深度增加活性显著增加(0.05)。两种处理下7月0—30 cm处土壤过氧化氢酶活性均高于12月(>0.05), 增温处理使40—50 cm深度处7月份土壤过氧化氢酶活性显著低于12月(<0.05)(图3)。重复方差分析结果表明, 不同取样时间和土壤深度交互作用对过氧化氢酶活性有显著影响(表1)。7月份土壤过氧化氢酶活性与土壤温度呈显著相关关系, 12月份土壤过氧化氢酶活性与土壤温度和水分均呈显著相关关系(表3)。
2.5 增温对土壤磷酸酶活性的影响
在7月份, 增温后土壤磷酸酶在0—10 cm、20—30 cm、40—50 cm深度处活性分别降低了14%、45%、31%(>0.05)。在12月份, 增温后土壤磷酸酶在0—10 cm、20—30 cm、40—50 cm深度活性分别增加了17%、11%、28%, 其中40—50 cm处差异达到显著水平(<0.05)。7月份土壤磷酸酶活性随土层深度增加活性降低, 12月份则随土层深度增加活性显著增加(<0.05)(图3)。对照样地7月份土壤磷酸酶活性显著高于12月份(<0.05), 增温样地7月份土壤磷酸酶活性低于12月份, 且在40—50 cm深度处差异显著(<0.05)。重复方差分析结果表明, 不同取样时间和增温处理交互作用对土壤磷酸酶活性有显著影响(表1)。7月份土壤磷酸酶活性与土壤温度和水分的相关关系不显著, 12月份土壤磷酸酶活性与土壤温度和土壤水分呈显著相关关系(表3)。

表1 增温、土壤深度和取样时间对土壤酶活性影响的重复方差分析结果
2.6 增温作用下夏季根系生物量的变化特征
OTC增温处理使高寒沼泽草甸0—30 cm深度范围的根系生物量显著减少(<0.05)(表2)。地下0—10 cm、10—20 cm 、20—30 cm、30—40 cm以及40—50 cm深度的生物量增温后分别减少49.41%、52.12%、48.62%、7.53%和7.72%。夏季根系生物量与土壤水分呈极显著相关关系。
3 讨论
开顶式生长室(OTC)能显著提高浅层土壤温度, 且对内部生态环境扰动小、造价低、安装实施方便, 普遍用于野外模拟增温实验, 由于野外样地往往地处高原腹地, 存在交通和电力供应不便等限制, 使OTC被动增温装置在研究中仍然发挥着重要作用[17]。试验期间, 增温使北麓河高寒沼泽草甸7月份、12月份近地表月平均气温分别增加4.55 ℃和6.64 ℃, 增温效果与高海拔地区温度增幅更大的结论相符[7-8]。从温度动态变化来看, 冬季土壤温度的增幅高于夏季, 与欧阳青、陈友超等人的研究结果一致, 参照陈友超等[18]的研究结果, 大气温度越高、降水越多, 土壤温度的增幅会越小。本研究中, 夏季平均大气温度比冬季高, 降水也主要集中在夏季, 故夏季土壤温度的增幅小。夏季增温样地的土壤水分低于对照样地, 是因为增温室内热量条件改变, 土壤蒸发和植被的蒸腾作用增强, 使增温室内土壤含水量减少。而在冬季, 增温处理提高了近地表空气温度和土壤温度, OTC内表层土体的冻结程度降低, 使OTC内土壤含水量高于对照样地。
温度上升一般会提高土壤酶的活性, 但是本研究中在7月份, OTC增温处理下土壤脲酶、蔗糖酶、过氧化氢酶和磷酸酶活性变化不显著, 总体有下降的趋势。这与Wang等[19]的研究结果一致, 增温对青藏高原那曲高寒草甸的土壤蔗糖酶、过氧化氢酶、磷酸酶活性无显著影响。原因可能是: (1)增温处理会加快有机质的矿化速率, 使土壤表层有机质含量降低, 即酶促反应的底物减少, 这种情况下, 微生物的产酶量变低[20-21]。(2)温度升高引起的土壤水分减少, 抑制了酶的合成与分泌[22]。增温后土壤含水量降低, 植物净初级生产力下降[23], 减少了植物的生物量。由于植物根系是土壤酶的来源之一, 其含量减少会对土壤酶活性产生一定影响[1-2]。本研究中, 夏季增温样地土壤含水量降低, 土壤水分与根系生物量呈极显著相关关系, 0—50 cm的根系生物量与对照样地相比均有减少, 夏季根系生物量与土壤蔗糖酶活性呈显著相关关系, 使增温后土壤蔗糖酶活性降低。另外随着土壤水分的降低, 现存的功能酶库由于扩散的限制不能自由地与基质相互作用, 使活性降低[24-25], 从而抵消了增温对土壤酶活性的正效应。(3)其余三种酶与土壤水分的相关关系不显著, 其活性变化不显著可能是由于不同的酶来源不同。杨林等[26]研究发现增温降低了细菌群落的多样性, 而对真菌群落并无显著影响, 因而没有影响到土壤中多数酶的活性。其次不同酶对温度的敏感性存在差异。有研究表明温度对土壤脲酶活性的影响会随处理时间的延长逐渐减弱[27-28]。孙辉等[29]发现, 土壤过氧化氢酶可能对高寒低温适应, 增温可能降低其活性; 还有研究表明在增温实验过程中, 不稳定碳库的相对丰度将下降, 可能导致水解酶的活性下降[25]。另外, 土壤酶作为一种蛋白质, 其稳定性也受到温度的影响, 增温引起的酶变性率的提高也会抵消温度升高对土壤酶活性的促进作用[1]。综合上述原因, 所以夏季增温对四种土壤酶活性的影响不显著。此外, 本研究中7月份四种土壤酶的活性均随着土壤深度增加而降低, 这与大部分人的研究结果一致[1-3,30], 土壤表层富含各种有机和无机物质, 可为微生物的生长发育提供能量和营养[31]。深层土壤的有机质受到土壤团聚体或有机无机复合物的保护, 土壤微生物较少且空间间隔也更远, 对酶的合成与分泌会采取更为保守的策略[32]。
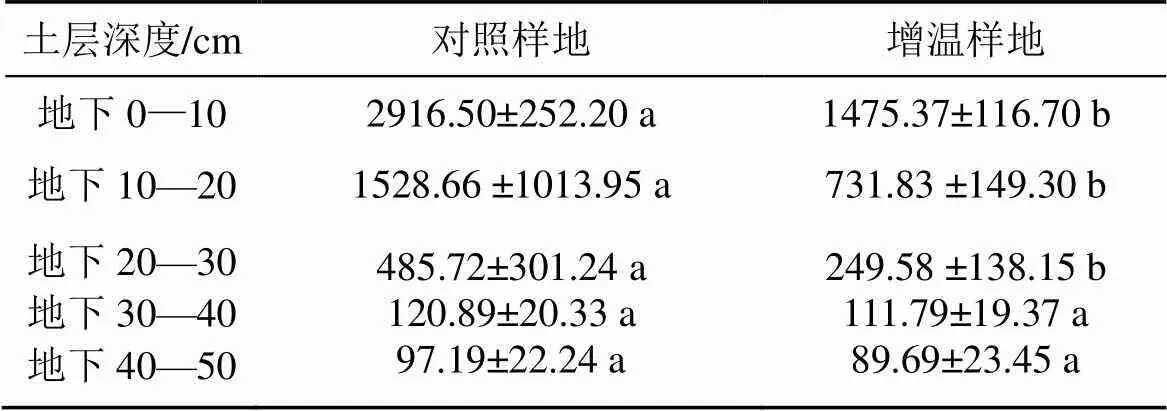
表2 增温作用下夏季根系生物量的分布特征 (g·m-2)
注: 不同小写字母表示同一土层中不同处理间差异显著(<0.05)。
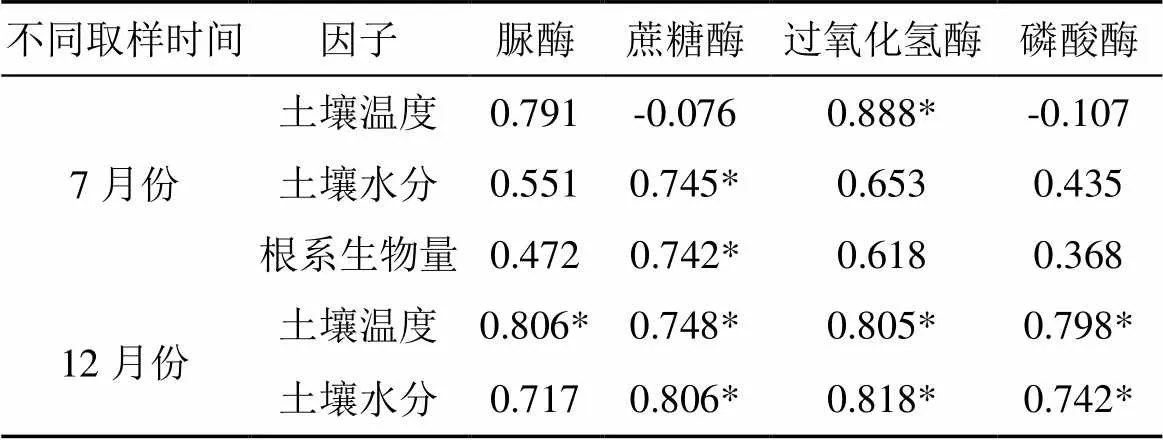
表3 土壤酶活性与土壤温度、土壤水分和根系生物量的相关系数
注: *<0.05。

注: 不同小写字母表示同一取样时间、同一土层深度中不同处理间差异显著(P<0.05); 不同大写字母表示在同一取样时间、同一处理中不同土层间差异显著(P<0.05)。
Figure 3 Changes of soil enzyme activities in different depths of alpine swamp meadows under warming and natural control conditions in July and December
在12月份, 增温使冬季不同土层四种土壤酶的活性均有所提高, 且在20—30 cm、40—50 cm处土壤蔗糖酶和过氧化氢酶活性和40—50 cm处脲酶和磷酸酶活性的增加达到显著水平(<0.05); 增温后四种土壤酶空间分布上均表现为40—50 cm深度活性最高; 且四种土壤酶的活性与土壤温度和水分相关关系显著。冬季对照样地0—10 cm处四种土壤酶的活性均比夏季小, 增温后, 冬季脲酶活性比夏季高, 其余三种酶的活性虽仍比夏季低, 但是差异性减小; 且在40—50 cm深度处四种土壤酶活性冬季显著高于夏季(<0.05)。一方面近地表温度的升高, 使冻结土体中可利用的自由水分增加, 进而提高土壤孔隙之间的连通以便于养分的利用, 为低温下幸存微生物(特别是低温嗜冷微生物)的修复和生长提供了必要条件[33], 而这一过程中存活的微生物可能会加强酶的活化效应。本研究中冬季40—50 cm处增温样地土壤温度相对较高, 相应的未冻水含量也较高, 有利于微生物的生长发育, 促进了酶的活化效能。由此表明土壤温度和土壤水分可能是冬季土壤酶活性的主要限制因素。这与Sardans等[31]的研究结果一致, 增温提高了冬春季土壤脲酶活性, 对夏秋季土壤脲酶活性的影响不显著, 因为冬春季增温样地土壤水分相对充足, 增温与水分的交互作用影响了土壤脲酶的活性, 而夏秋季土壤水分缺乏使土壤脲酶活性无显著变化。另一方面, 这种冬季冻结期间酶活性的升高, 可能是因为冻结作用导致死亡生物细胞破裂释放出营养物质, 供低温下幸存微生物吸收利用, 从而提高了酶活性, 以及雪被的保温作用为适应低温的土壤微生物提供了较为稳定的微环境, 有利于增加土壤酶的生产和活性[34]。
低的养分有效性往往是高寒草地生态系统的主要限制因子[35], 增温后温度、水分以及与养分密切相关的土壤酶活性的变化必将对返青期青藏高原植物生长产生影响, 而植物的生长变化又将改变地面反照率、净辐射量、物质的能量交换等一系列生物物理参数, 最终导致高原生态过程发生变化, 从而对青藏高原生态安全屏障功能及当地畜牧业生产产生影响[36]。研究发现, 如果土壤变暖作为全球变暖的一部分继续下去, 碳、氮循环很可能通过与碳、氮相关的土壤酶受到显著的影响[24]。所以研究增温作用下, 与碳、氮、磷等元素相关的土壤酶活性在不同季节、不同土壤深度处的响应, 能为分析全球气候变暖背景下, 高寒沼泽草甸生态过程的变化提供一定的理论依据。
4 结论
增温后青藏高原高寒沼泽草甸土壤脲酶、蔗糖酶、过氧化氢酶和磷酸酶活性受到土壤深度与季节的综合调控。在季节上, 增温对土壤酶活性的影响冬季大于夏季。夏季土壤酶受到土壤水分和根系生物量降低等因素的影响, 抵消了增温的正效应, 对增温的响应不显著。冬季土壤酶活性在增温作用下有不同程度的提高, 同时受不同深度土壤水分、温度的制约, 在土壤温度、水分含量均较高的40—50 cm处变化显著。在空间分布上, 夏季增温样地四种土壤酶活性总体随土层深度增加而降低, 冬季随土层深度增加而显著升高。未来气候变暖背景下, 为准确评估青藏高原高寒沼泽草甸的土壤-生态变化过程, 应系统研究不同季节、不同土壤深度下土壤酶活性—微生物种群结构—土壤养分变化—植被生长间相互关系。
[1] GIRISH S, AJIT V. Soil Enzymology [M]. Berlin Heidelberg: Springer-Verlag, 2013.
[2] 关松荫, 张德生, 张志明. 土壤酶及其研究法[M]. 北京: 农业出版社, 1986.
[3] 郑蔚, 周嘉聪, 林伟盛, 等. 土壤增温对亚热带杉木幼树不同深度土壤微生物胞外酶活性的影响[J]. 应用生态学报, 2019, 30(3): 832–840.
[4] JING Xin, WANG Yonghui, CHUNG H, et al. No temperature acclimation of soil extracellular enzymes to experimental warming in an alpine grassland ecosystem on the Tibetan Plateau[J]. Biogeochemistry, 2014, 117(1): 39– 54.
[5] 李红琴, 徐海燕, 马小亮, 等. 马衔山多年冻土与季节冻土区土壤微生物量及酶活性的季节动态[J]. 冰川冻土, 2017, 39(2): 421–428.
[6] IPCC. Climate Change 2013: The Physical Science Basis[M]. UK: Cambridge University Press, 2013.
[7] XIA Jiayang, CHEN Jiquan, PIAO Shilong, et al. Terrestrial carbon cycle affected by non-uniform climate warming[J]. Nature Geoscience, 2014, 7(3): 173–180.
[8] 钞然, 张东, 陈雅丽, 等. 模拟增温增雨对典型草原土壤酶活性的影响[J]. 干旱区研究, 2018, 35(5): 1068–1074.
[9] 李娜, 王根绪, 高永恒, 等. 模拟增温对长江源区高寒草甸土壤养分状况和生物学特性的影响研究[J]. 土壤学报, 2010, 47(6): 1214–1224.
[10] 蒋观利, 吴青柏, 张中琼. 青藏高原不同高寒生态系统类型下多年冻土活动层水热过程差异研究[J]. 冰川冻土, 2018, 40(1): 7–17.
[11] 李成阳, 赖炽敏, 彭飞, 等. 青藏高原北麓河流域不同退化程度高寒草甸生产力和群落结构特征[J]. 草业科学, 2019, 36(4): 1044–1052.
[12] 范思睿, 范广洲, 董一平, 等. 青藏高原四季划分方法探讨[J]. 高原山地气象研究, 2011, 31(2): 1–11.
[13] WALKER T W D, JANSSENS I A, WEEDON J T, et al. A systemic overreaction to years versus decades of warming in a subarctic grassland ecosystem[J]. Nature ecology & evolution, 2020, 4(1): 101–108.
[14] 欧阳青, 任健, 尹俊, 等. 短期增温对亚高山草甸土壤养分和脲酶的影响[J]. 草业科学, 2018, 35(12): 2794–2800.
[15] 岳广阳, 赵林, 王志伟, 等. 多年冻土区高寒草甸根系分布与活动层温度变化特征的关系[J]. 冰川冻土, 2015, 37(5): 1381–1387.
[16] 陈慷惋. 转基因棉花的毒素在土壤中的分布及对土壤酶活性的影响[D]. 武汉: 华中农业大学, 2008.
[17] 牛书丽, 韩兴国, 马克平, 等. 全球变暖与陆地生态系统研究中的野外增温装置[J]. 植物生态学报, 2007, 31(2): 262–271.
[18] 陈有超, 鲁旭阳, 李卫朋, 等. 藏北典型高寒草原土壤微气候对增温的响应[J]. 山地学报, 2014, 32(4): 401–406.
[19] WANG Xuexia, DONG Shikui, GAO Qingzhu, et al. Effects of short-term and long-term warming on soil nutrients, microbial biomass and enzyme activities in an alpine meadow on the Qinghai-Tibet Plateau of China[J]. Soil Biology & Biochemistry, 2014, 76: 140–142.
[20] ALLISON S D, VITOUSEK P M. Responses of extracellular enzymes to simple and complex nutrient inputs [J]. Soil Biology Biochemistry, 2005, 37(5): 937–944.
[21] 赵艳艳, 周华坤, 姚步青, 等.长期增温对高寒草甸植物群落和土壤养分的影响[J]. 草地学报, 2015, 23(4): 655– 671.
[22] CARLYLE C N, FRASER L H, TURKINGTON R. Tracking soil temperature and moisture in a multi-factor climate experiment in temperate grassland: Do climate manipulation methods produce their intended effects? [J]. Ecosystems, 2011, 14: 489–502.
[23] 石福孙, 吴宁, 罗鹏.川西亚高山草甸植物群落结构及生物量对温度升高的响应[J]. 生态学报, 2008, 28(11): 5286–5293.
[24] LIORENS L, PENUELAS J, ESTIARTE M, et al. Contrasting growth changes in two dominant species of a Mediterranean shrubland submitted to experimental drought and warming[J]. Annals of Botany, 2004, 94(6): 843–853.
[25] JING Xin, WANG Yonghui, CHUNG H, et al. No temperature acclimation of soil extracellular enzymes to experimental warming in an alpine grassland ecosystem on the Tibetan Plateau[J]. Biogeochemistry, 2014, 117 (1): 39–54.
[26] 杨林, 陈亚梅, 和润莲, 等. 高山森林土壤微生物群落结构和功能对模拟增温的响应[J]. 应用生态学报, 2016, 27(9): 2855–2863.
[27] 白春华, 红梅, 韩国栋, 等. 土壤三种酶活性对温度升高和氮肥添加的响应[J]. 内蒙古大学学报(自然科学版), 2012, 43(05): 509–513.
[28] BAI Yinping, LI Feng, YANG Gang, et al. Meta-analysis of experimental warming on soil invertase and urease activities[J]. Acta Agriculturae Scandinavica, Section B- Soil & Plant Science, 2018, 68(2): 104–109.
[29] 孙辉, 吴秀臣, 秦纪洪, 等. 川西亚高山森林土壤过氧化氢酶活性对升高温度和CO2浓度的响应[J]. 土壤通报, 2007, 38(5): 891–895.
[30] 黄文华, 王树彦, 韩冰, 等. 草地生态系统对模拟大气增温的响应[J]. 草业科学, 2014, 31 (11) : 2069–2076.
[31] SARDANS J, PENUELAS J, ESTIARTE M. Changes in soil enzymes related to C and N cycle and in soil C and N content under prolonged warming and drought in a Mediterranean shrubland[J]. Applied Soil Ecology, 2007, 39 (2): 223–235.
[32] SALOME C, NUNAN N, POUTEAU V, et al. Carbon dynamics in topsoil and in subsoil may be controlled by different regulatory mechanisms[J]. Global Change Biology, 2009, 16(1): 416–426.
[33] 王奥. 季节性冻融对高山森林土壤微生物与生化特性的影响[D]. 雅安: 四川农业大学, 2012.
[34] 尹鹏, 胡霞, 廖金花, 等.高山地区土壤微生物生物量和土壤酶活性对雪被变化的响应[J]. 生态科学, 2015, 34(1): 91–96.
[35] 徐振锋, 唐正, 万川, 等. 模拟增温对川西亚高山两类针叶林土壤酶活性的影响[J]. 应用生态学报, 2010, 21(11): 2727–2733.
[36] 丁明军, 张镱锂, 刘林山, 等. 青藏高原植物返青期变化及其对气候变化的响应[J]. 气候变化研究进展, 2011, 7(5): 317–323.
Effects of experimental warming on soil enzyme activities in different seasons of the alpine swamp meadow ecosystem on the Qinghai-Tibet Plateau
GAO Yu1,2, YONG Shaoning3, WANG Junfeng1,*
1. State Key laboratory of Frozen Soil Engineering, Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China 2.University of Chinese Academy of Sciences, Beijing 100049, China 3.Gansu Provincial Department of Transportation, Lanzhou 730000, China
In order to study the response characteristics of soil enzyme activities of the alpine swamp meadow ecosystem distributed in permafrost region of the Qinghai-Tibet Plateau to climate warming, an experimental site with alpine swamp meadow ecosystem was selected in the Beiluhe area and an Open Top Chamber (OTC) method was used to conduct a warming experiment. In July and December of 2019, soil samples at the warming and the control plots were collected in soil depths of 0-10 cm, 20-30 cm, and 40-50 cm, respectively, and activities of soil urease, invertase, catalase and phosphatase at the different soil layers were analyzed. The results showed that in July, when the average monthly near-surface air temperature at the warming plots was 4.55 ℃ higher than that at the control plots, the soil urease, invertase, catalase and phosphatase activities of different soil depths had a decreasing tendency, but did not reach a significant level(>0.05). Soil invertase activity was significantly correlated with soil moisture and root biomass(<0.05). In December, at the condition that the average monthly near-surface air temperature at the warmed plots was 6.64 ℃ higher than that at the control plots, the activities of soil urease, invertase, catalase and phosphatase at different depths all increased, but only the activities of soil invertase and phosphatase at the depth of 20-50 cm, and the activities of soil urease and catalase at the depth of 40-50 cm reached significant levels (<0.05).The activities of the four soil enzymes were significantly correlated with soil temperature and moisture(<0.05). For the changes of enzyme activities in depth, the four soil enzyme activities in summer generally decreased with the increase of soil depth (<0.05. In winter, the four soil enzyme activities all rose significantly with the increase of soil depth (<0.05). At the warming plots, the activities of the four soil enzymes in different soil depths in December were generally greater than those in July, and the difference was significant at 40-50 cm depth (<0.05). The effect of warming on soil enzyme activity in winter was greater than that in summer. In summer, soil enzyme activity was mainly affected by soil moisture and root biomass, and its response to warming was not significant. While in winter, the soil enzyme activities were usually restricted by soil moisture and temperature at different depths, and the soil enzyme activities were higher at the depths with high soil temperature and water content.
soil enzyme activity; experimental warming; alpine swamp meadow;Qinghai-Tibet Plateau
高钰, 雍少宁, 王俊峰. 增温对青藏高原高寒沼泽草甸不同时期土壤酶活性的影响[J]. 生态科学, 2023, 42(1):1–10.
GAO Yu, YONG Shaoning, WANG Junfeng. Effects of experimental warming on soil enzyme activities in different seasons of the alpine swamp meadow ecosystem on the Qinghai-Tibet Plateau[J]. Ecological Science, 2023, 42(1): 1–10.
10.14108/j.cnki.1008-8873.2023.01.001
S154.1
A
1008-8873(2023)01-001-10
2020-10-26;
2020-11-23
国家自然科学基金项目(41771080); 国家自然科学基金青年基金项目(41003032); 甘肃省交通运输厅项目(E1390602)
高钰(1996—), 女, 山西吕梁人, 硕士研究生, 主要从事寒区生态环境与气候变化研究, E-mail: gaoyu18@mails.ucas.ac.cn
王俊峰, 男, 博士, 研究员, 主要从事寒区生态与环境变化研究, E-mail: wangjf2008@lzb.ac.cn
猜你喜欢
生物技术通报(2020年12期)2020-12-21
腐植酸(2020年1期)2020-11-29
水土保持研究(2019年6期)2019-10-19
浙江理工大学学报(自然科学版)(2018年6期)2018-11-07
辽宁林业科技(2017年4期)2017-06-22
中国组织化学与细胞化学杂志(2017年1期)2017-06-15
浙江大学学报(工学版)(2016年2期)2016-06-05
广西林业科学(2016年1期)2016-03-20
医学研究杂志(2015年8期)2015-06-22
中国医学科学院学报(2013年6期)2013-03-11
