
基于LC/MS的伊犁马3 600 m速度赛赛前、赛后血浆代谢组学差异变化
2023-04-14 08:27张仕琦李晓斌张文杰王世昌郑文祥欧阳文祁居中杨开伦
新疆农业科学 2023年2期
张仕琦,李晓斌,张文杰,韩 明,王世昌,郑文祥,欧阳文,祁居中,杨开伦
(1.新疆农业大学动物科学学院/新疆肉乳用草食动物营养重点实验室,乌鲁木齐 830052;2.新疆伊犁哈萨克自治州昭苏县种马场,新疆昭苏 835000)
0 引 言
【研究意义】运动马在不同生理活动(训练、竞赛)中,肌肉细胞收缩、能量消耗以及体内代谢产物的积累,导致力的下降,产生运动性疲劳[1],掌握机体运动过程中代谢产物变化情况,对于预防、恢复运动疲劳和提高运动性能有重要作用,能够为运动马的营养调控提供思路。代谢组学可以通过血液或尿液代谢产物的整体模式测定或指纹分析,进一步揭示机体代谢变化[2],对于马匹运动性能的调控可能发挥积极作用。【前人研究进展】Enea等[3]利用代谢组学技术揭示长时间运动(75%的VO2max,直至耗竭)和短时间的运动(短期、密集性运动厌氧试验)机体尿液代谢产物具有差异。Lehmann等[4]利用代谢组学方法鉴定出人在中等强度运动中,中等链酰基肉碱是具有促进脂肪氧化能力的主要生物标记物。Lewis等[5]利用质谱技术对运动员运动前后200多个代谢物进行了测定,得到了综合的血浆代谢物特征。【本研究切入点】国内运动马产业快速发展,国产运动马运动性能上的差距以及运动性疲劳现象等问题日益显现。随着科技设备的发展,马匹运动过程中的生理参数如心率、呼吸等能够即时测定,但血液中的代谢物目前仍未能实现运动过程中的即时监测,马匹在运动过程中生化状态变化的数据较难获取,也未见报道。【拟解决的关键问题】试验采用赛后即刻状态趋近于运动过程中状态的方法,应用非靶向代谢组学检测分析技术,研究伊犁马3 600 m速度赛前和赛后即刻血液中主要代谢物的差异变化,为运动马运动过程中的代谢调控提供新的参考和思路。
1 材料与方法
1.1 材 料
1.1.1 饲粮组成
根据马场提供的遗传系谱,挑选5匹(3♂+2♀)无亲缘关系、体重(384.40±34.18)kg、年龄为2岁的正常速度赛用伊犁马,试验马匹每日等强度训练25 d后,于晨饲4 h进行3 600 m速度赛,试验马以(11.75±0.87)m/s的平均速度完成比赛。赛前安稳状态下和赛后即刻测定心率和呼吸频率,采集血液、制备血浆样品,采集血浆的质谱,采用非靶向性代谢组学方法进行代谢物的测定。表1

表1 饲粮组成及营养水平表(干物质基础,%)Table 1 Composition and nutrient levels of the diet (DM basis,%)
试验马匹单厩饲养,每天按时打扫马厩。每天每匹试验马饲喂相同饲粮(8 kg干牧草和4 kg精料补充料)。每天分3次饲喂(07:00,15:00和22:00),先粗后精,自由饮水。
1.2 方 法
1.2.1 试验设计
分别在试验正试期第25 d进行的3 600 m速度赛的赛前安稳状态下、赛后即刻(即马匹完成赛程时立即采集血液),测定呼吸频率和心率,采集颈静脉血液(肝素钠抗凝管)。血液以3 500 r/min离心10 min后收集上清血浆,分装,液氮冻存。
1.2.2 样品预处理
试验样本预处理:取100 μL血浆加入80%的甲醇水溶液400 μL,涡旋振荡后在-20℃条件下静置60 min,于14 000×g、低温离心20 min,取上清取进行真空冷冻干燥,加入100 μL的复溶剂进行溶解残留物,混匀后,于14 000×g、低温离心15 min,正离子模式取上清液8 μL、负离子模式取上清液12 μL进行LC-MS分析。
质控(QC)样本预处理:从每个已处理好的样本中取等量上清液混匀作为QC样本。blank样本为试验样本的空白基质,样本前处理过程与试验样本相同。
1.2.3 色谱和质谱分析
采用超高效液相色谱系统(Vanquish UHPLC)对10个样品进行分离。柱温40℃;流速:0.3 mL/min;正模式流动相组成:流动相A:0.1%甲酸,95%乙腈,10 mmol/L醋酸铵。流动相B:0.1%甲酸,50%乙腈,10 mmol/L醋酸铵。负模式流动相组成:流动相A:95%乙腈,10 mmol/L醋酸铵,pH9.0。流动相B:50%乙腈,10 mmol/L醋酸铵,pH9.0。分离后用QE HF-X质谱仪进行质谱分析。扫描范围选择m/z 100-1 500;ESI源的设置如下:Spray Voltage:3.2 kV;Sheath gas flow rate:35 arb;Aux Gas flow rate:10 arb;Capillary Temp:320 °C。Polarity:positive;negative;MS/MS二级扫描为data-dependent scans。表2

表2 色谱梯度洗脱程序Table 2 Chromatographic gradient elution procedure
1.2.4 游离氨基酸定量测定
取50 μL血浆加50 μL蛋白沉淀剂(含NVL),涡旋震荡,13 200 r/min、4℃离心4 min去蛋白,取8 μL上清加入42 μL标记缓冲液和20 μL衍生液混匀,于55℃恒温条件下衍生15 min后冷却、离心,取50 μL上清进行LC-MS检测。
1.3 数据处理
采用SPSS 22.0统计软件的配对样本t检验对心率、呼吸频率和游离氨基酸定量数据进行差异分析,结果以“平均值±标准差”(mean±SD)表示,以P<0.05为差异显著水平,P<0.01为差异极显著水平,0.05≤P≤0.1具有差异趋势。
非靶向代谢组学数据:将下机数据文件导入Compound discoverer(CD)搜库软件中,设置参数质量偏差0.005‰、信号强度偏差30%、信噪比3、最小信号强度100 000、加和离子等信息进行峰提取,用QC样本对定量结果进行归一化,对数据进行质控,得到质控合格的数据。分析PCA和PLS-DA。PLS-DA模型以变量投影重要度VIP>1.0,差异倍数FC>1.2或FC<0.833为差异显著水平。
2 结果与分析
2.1 3 600 m速度赛前、后伊犁马的心率和呼吸频率变化
研究表明,试验马匹在3 600 m速度赛过程中心率、呼吸频率显著升高(P<0.05)。表3
2.2 代谢物质总离子色谱图
研究表明,伊犁马血浆经过正、负离子检测,得到的两个二维数据阵。数据阵中质荷比、保留时间及其峰面积等数据,在mzCloud数据库中进一步对化合物精确定性和相对定量。图1

表3 3 600 m速度赛前、后伊犁马的心率 和呼吸频率Table 3 Heart rate and respiratory rate of the Yili horse before and after 3 600 m speed race(bpm)
2.3 代谢差异产物
2.3.1 偏最小二乘法判别(PLS-DA)
研究表明,样本都处于楠圆置信区间内,并且5个质控样品能够较好的聚类。赛前、赛后血浆样品有明显分开区域。试验伊犁马血浆正离子和负离子模式数据建立的PLS-DA模型的解释率和预测能力强,且排序检验R2>Q2,解释率大于该模型的预测率,模型未“过拟合”,模型可靠,能很好的描述样本。图2,图3
2.3.2 代谢差异产物的筛选
研究表明,与赛前相比,伊犁马赛后血浆甘油、柠檬酸、琥珀酸、精氨基琥珀酸、乳酸、丙酮酸、α-酮戊二酸、尿酸、犬尿氨酸、胱氨酸、N-乙酰天冬氨酸、苯丙氨酸、谷氨酸、乙酰左旋肉碱含量显著升高(VIP>1.0)。血浆葡萄糖、D-葡萄糖-6-磷酸、亮氨酸、色氨酸、组氨酸、甜菜碱、谷氨酰胺、脯氨酸、苏氨酸、乙酰精氨酸、混旋肉碱、辣椒素、多巴胺含量显著降低(VIP>1.0)。表4
2.4 游离氨基酸定量
研究表明,伊犁马3 600 m速度赛赛后血浆氨基酸含量的变化较大,为进一步验证,又进行血浆氨基酸定量测定。伊犁马3 600 m速度赛赛后即刻与赛前相比血浆多种氨基酸含量产生显著变化,其中:血浆Asp、Lys含量极显著升高(P<0.01);Trp、His、Asn含量以及色氨酸/支链氨基酸比值极显著降低(P<0.01);Ala含量显著升高(P<0.05)。表5
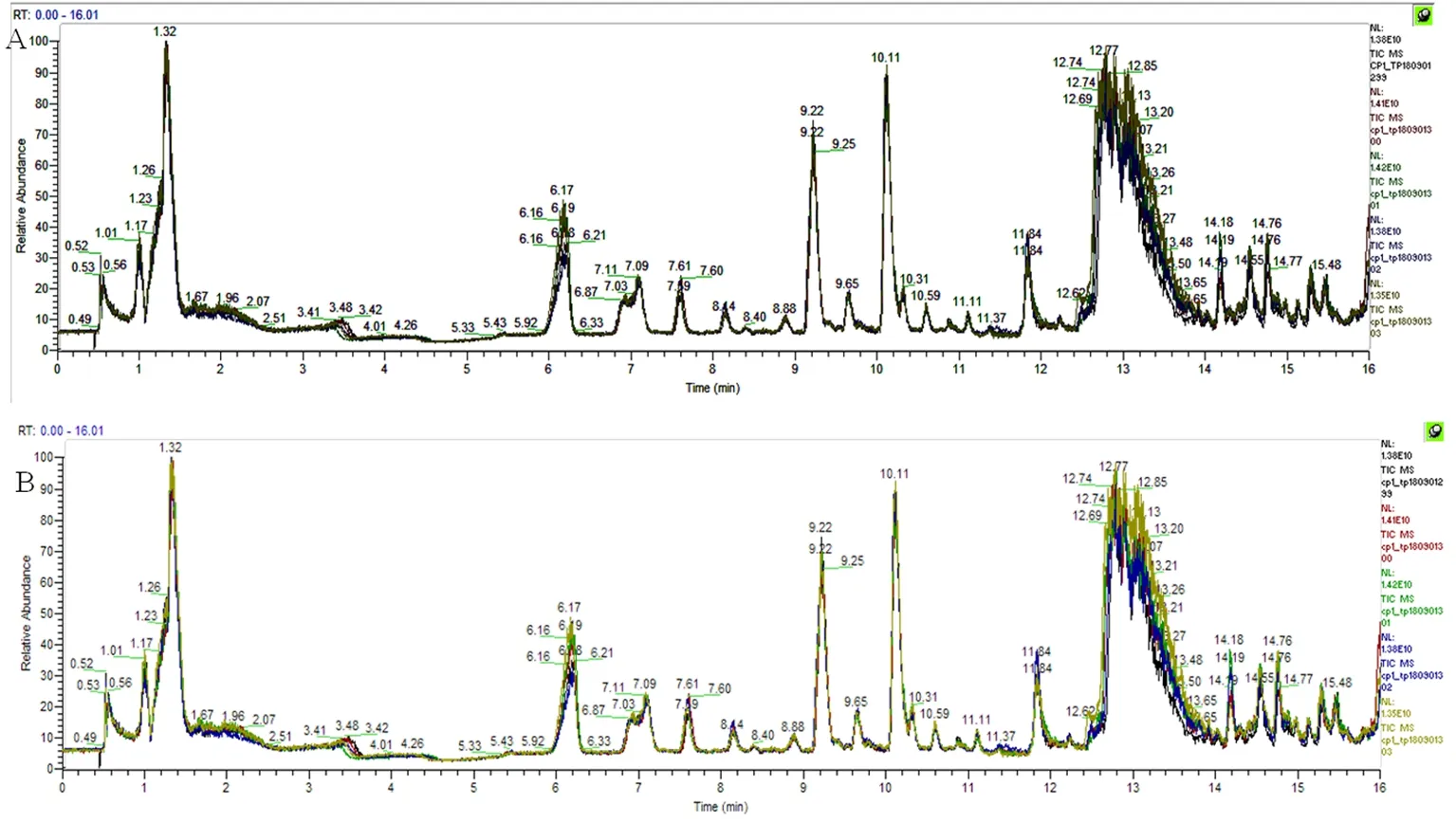
注:A为正离子模式,B为负离子模式
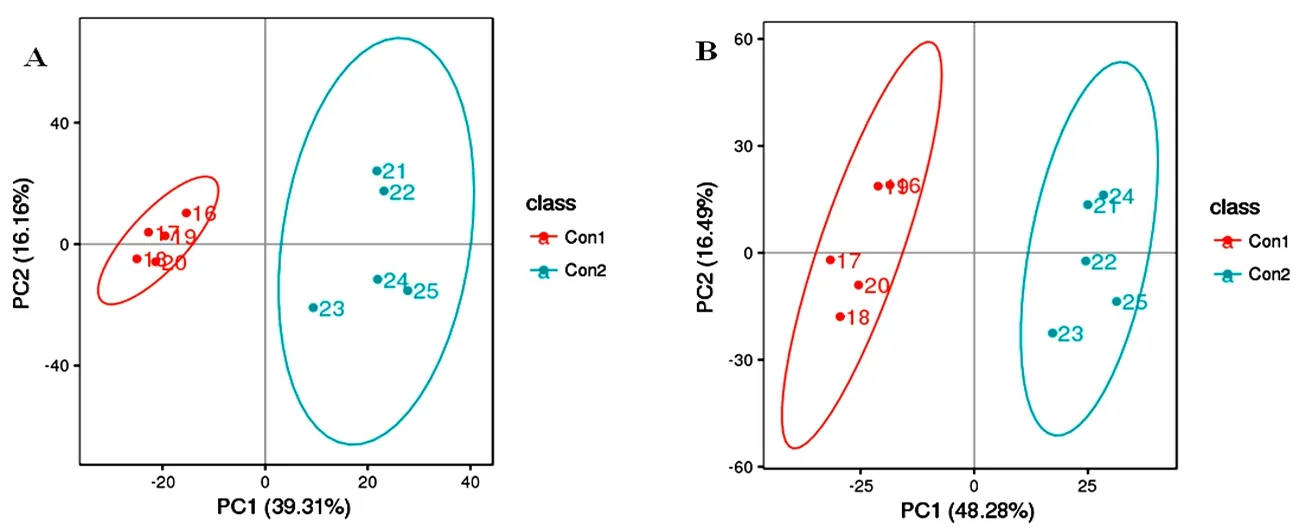
注:图中置信椭圆为95%。A为正离子模式,B为负离子模式,下同
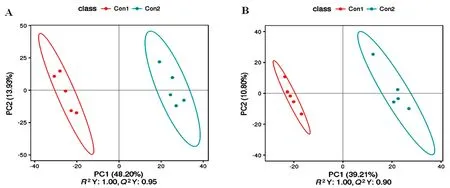
注:R2:表示模型解释率;Q2:表示模型预测能力

表4 伊犁马3 600 m速度赛赛前赛后的血浆代谢差异物(n=5)Table 4 Plasma differential metabolites before and after 3 600 m Speed Race in Yili horse(n=5)
2.5 KEGG富集分析
研究表明,经过KEGG富集后,伊犁马在3 600 m速度赛中柠檬酸循环、丙氨酸、天冬氨酸和谷氨酸代谢、五糖和葡萄糖醛酸间转化、半胱氨酸与蛋氨酸代谢、氧化磷酸化、脂肪酸降解、鞘脂代谢等途径呈现显著性变化(P<0.05)。表6
3 讨 论
3.1心率30~42次/min,呼吸数8~16次/min是成年马的正常阈值[6]。试验马匹在赛前心率和呼吸频率在正常范围内,马匹的生理状态稳定。赛后即刻试验马匹心率和呼吸频率达到较高水平,表明运动期间马匹心肺功能快速运转,增加血氧供应,促进机体氧化供能,使得血液中营养物质快速代谢转换。利用非靶向代谢组学的方法对伊犁马3 600 m速度赛赛前、赛后即刻两个时间点的血浆代谢小分子进行测定,发现能量代谢、氨基酸代谢、脂质代谢等途径在伊犁马运动过程中呈现显著性变化。
3.2伊犁马3 600 m速度赛后即刻血浆柠檬酸、琥珀酸含量上升。柠檬酸、琥珀酸是需氧生物体内能源物质的最终代谢通路—柠檬酸循环(三羧酸循环,TCA)的重要物质,是有氧代谢供能的中枢环节。在三羧酸循环中乙酰胆碱和草酰乙酸生成的柠檬酸,柠檬酸的含量可反应机体内能源物质有氧氧化代谢的状况。表明伊犁马在3 600 m速度赛过程中有氧氧化参与代谢供能,加速了三羧酸循环的物质代谢。
3.3伊犁马3 600 m速度赛后即刻血浆葡萄糖含量下降,丙酮酸、乳酸含量上升。说明马匹在剧烈运动时ATP快速分解供能且不断消耗氧气,当磷酸肌酸耗尽后,肌细胞逐渐呈现厌氧酵解状态,能量代谢转为糖酵解途径,分解糖原,产生大量乳酸[7-8]和丙酮酸。丙酮酸加氢还原成乳酸后继续分解释放ATP[9]。在运动员比赛过程中糖无氧供能加强,赛前、赛后乳酸和葡萄糖浓度显著变化[10]。
伊犁马3 600 m速度赛后血浆葡萄糖浓度降低。葡萄糖是机体内主要的供能物质,血浆葡萄糖浓度降低可能是因为肌肉运动刺激APMK通路,加速葡萄糖向肌肉转运速度,以供肌肉氧化供能[11]。马拉松运动员赛后[12]以及大鼠一次性力竭运动后[13]血清葡萄糖浓度也呈现显著降低态势。

表5 伊犁马3 600 m速度赛赛前赛后的血浆游离氨基酸含量变化(n=5, μmol/L)Table 5 Changes of free amino acids in pasma before and after 3 600 m Speed Race in Yili horse(n=5, μmol/L)
研究中赛后即刻伊犁马血浆乳酸浓度升高。乳酸是糖酵解的最终产物,机体运动导致无氧代谢增强,乳酸在体内不断积累[14],使得血浆乳酸浓度升高。研究表明,运动员训练后乳酸酵解能系统参与机体供能,血清中乳酸含量升高[10]。乳酸根离子(La-)与钠(Na+)或钾(K+)离子形成盐,对肌肉细胞不构成威胁,但肌肉中H+的积累,降低pH能够抑制肌动蛋白和肌球蛋白纤维的相互作用,以及抑制肌浆网中钙离子(Ca2+)再摄取,可能延长肌肉收缩时间并导致疲劳[15-16]。乳酸是判断运动性疲劳的标志物之一。
伊犁马在3 600 m速度赛过程中供能方式不仅有葡萄糖和柠檬酸参与的有氧氧化,还有在耗氧状态下,糖酵解系统逐渐参与机体供能,也逐渐积累了能够引起降低力的乳酸。
3.4伊犁马血浆各氨基酸含量赛前、赛后即刻出现显著差异,伊犁马在运动过程中血浆氨基酸含量有显著影响。血液中氨基酸的浓度不高,但更新迅速,一些组织器官可以不断地从血液中摄取/释放氨基酸,尤其是骨骼肌[17]。机体运动时,血液多集中在肢体骨骼肌中,且随着骨骼肌运动收缩,能量代谢,血液循环也加快,使得血液中氨基酸含量也发生变化。
伊犁马3 600 m速度赛后即刻血浆谷氨酸含量显著升高,血浆谷氨酰胺含量显著降低。可能说明在伊犁马运动过程中机体谷氨酰胺快速分解用于供能产生的谷氨酸。谷氨酸和谷氨酰胺可在体内相互转化,运动时谷氨酰胺可分解产生谷氨酸间接参与能量供应,运动强度越大谷氨酰胺浓度下降越快[18]。骨骼肌中谷氨酰胺持续下降被认为是过度训练的指标之一[19]。Glu和Gln不仅能促进氨代谢,还可促进机体氧化供能。且运动时Glu+丙酮酸=Ala+α-酮戊二酸反应增强,可减少乳酸生成[20-21]。
伊犁马3 600 m速度赛后血浆亮氨酸、色氨酸含量显著降低,犬尿氨酸含量升高。亮氨酸属于支链氨基酸,可参与供能。在运动过程中肌糖原减少,BCAA氧化参与供能,使得运动后血浆中BCAA浓度下降[22]。色氨酸是5-羟色胺(5-HT)的前体,脑内5-HT合成增加就是由于大脑吸收血液运输传递的游离Trp而引起的,可调节神经兴奋系统。在运动过程中BCAA浓度不仅与运动供能有关,也关系到脑中游离Trp的浓度,因为BCAA能竞争结合Trp在血脑屏障上的载体-白蛋白通道[23-24],BCAA被氧化为骨骼肌收缩过程提供的能量,导致BCAA减少,减少竞争抑制使得更多游离Trp进入到大脑中,利于5-HT的合成,造成中枢疲劳[25-26]。亮氨酸及其代谢通路对外周疲劳和中枢疲劳均可产生影响。亮氨酸、色氨酸及其涉及的代谢通路可作为营养调控的参考方向之一。
伊犁马赛后血浆甜菜碱含量显著降低。可能是在伊犁马运动过程中甜菜碱参与了能量代谢。甜菜碱以及胆碱主要是机体甲基供给物,其中一条甲基供给路线是机体肌酸合成代谢,甜菜碱可反应生成蛋氨酸,然后在腺苷酶催化生成腺苷蛋氨酸(SAM)又经过S-腺苷蛋氨酸-胍基乙酸N-甲基转移酶(GAMT)的催化下形成肌酸[27-28],参与磷酸原供能系统。因此甜菜碱-蛋氨酸循环也可作为营养调控的参考方向之一。
3.5脂质中包括甘油三酯和类脂,而甘油三脂由脂肪酸和甘油合成,作为骨骼肌的主要能量来源,与耐力运动性能的发挥有重要关系。可能由于机体中脂肪细胞缺少甘油激酶,运动中脂质分解产生的甘油,必须通过血液送至肝脏进行代谢[29],再参与三羧酸循环彻底氧化供能(氧充足)或糖异生产生乳酸(供氧不足)。伊犁马在运动过程中机体脂质也参与了分解供能,使得血浆甘油含量升高。
运动时糖酵解系统供能比例增加,并且当生成CoA的速率大于柠檬酸的利用率时,肉碱则充当乙酰基的受体生成乙酰肉碱[30],伊犁马3 600m速度赛后血浆混旋肉碱含量降低,乙酰左旋肉碱含量显著升高。左旋肉碱是可运输脂肪酸进入线粒体进行TCA和β-氧化循环[31]。研究证明,适当补充左旋肉碱可改善脂肪代谢,进而改善机体有氧能力[32]。肉毒碱也可作为营养调控的参考方向之一。
伊犁马在3 600 m速度赛中参与能量供应的代谢物变化最为显著。运动前期磷酸原供能系统快速供能[33],甜菜碱、蛋氨酸参与肌酸合成,运动前后变化显著。磷酸原供能短暂时间后转变为有氧氧化供能;线粒体三羧酸循环供能、葡萄糖氧化、氨基酸(支链氨基酸)氧化、脂肪氧化加强,促进肌肉供能。随后氧气供应不足,供能方式转变为糖酵解供能。运动后期主要依赖脂肪(甘油)经过甘油激酶等催化作用生成磷酸二羟丙酮作用下进行糖酵解或者参与β-氧化供能。伊犁马在比赛期间呈现无氧、有氧以及混氧的代谢特点。因此,在考虑调控运动马运动性能方面,提高马匹运动过程中供能物质的供给具有重要意义。其次,运动过程中糖酵解供能时间较长,乳酸、氨等代谢物的堆积导致力的衰减,减少代谢废物的堆积以及加速代谢物的排出对于运动性能的提高以及运动后疲劳恢复也很重要。谷氨酰胺、精氨酸、谷氨酸等功能性氨基酸可能对于代谢废物的排出或减少产生有积极作用。结合试验结果,供能物质亮氨酸、甘油、蛋氨酸、甜菜碱以及功能物质色氨酸、谷氨酰胺、肉毒碱这几类物质涉及的调节通路可能为运动马营养调控提供参考。
4 结 论
基于LC/MS伊犁马血浆代谢组学的分析,3 600m速度赛伊犁马机体呈现出无氧、有氧以及混氧代谢的特点。伊犁马3 600 m速度赛前、赛后血浆样本的LC/MS代谢谱存在差异,涉及柠檬酸循环、糖酵解系统以及脂肪代谢、氨基酸代谢等过程。其中,亮氨酸、色氨酸、谷氨酰胺、蛋氨酸、甜菜碱、甘油、肉毒碱这几类物质涉及的调节通路可能为运动马营养补剂的研发提供参考。
猜你喜欢
中国饲料(2022年5期)2022-04-26
西部(2021年3期)2021-07-11
西北农业学报(2020年9期)2020-09-10
中国调味品(2017年2期)2017-03-20
草食家畜(2016年1期)2016-03-18
中外医疗(2015年11期)2016-01-04
医学研究杂志(2015年12期)2015-06-10
中国洗涤用品工业(2015年8期)2015-02-28
食品工业科技(2014年9期)2014-03-11
无机化学学报(2014年9期)2014-02-28
