
天山北坡中部天山云杉1 a生天然更新幼苗存活数量与功能性状的微生境分析
2023-04-14 08:27胡文聪潘存德赵善超宋梦真童海麦田晨阳
新疆农业科学 2023年2期
胡文聪,潘存德,赵善超,宋梦真,童海麦,田晨阳
(1.新疆农业大学林学与风景园林学院/新疆教育厅干旱区林业生态与产业技术重点实验室,乌鲁木齐 830052;2.新疆林业和草原局,乌鲁木齐 830000)
0 引 言
【研究意义】天山云杉(Piceaschrenkianavar.tianschanica)是天山森林群落最主要的建群种,对天山森林生态系统的形成与维持有着至关重要的作用。天山云杉良好的天然更新是天山森林生态系统可持续发展的基础,但地表的微生境异质性对其更新幼苗的发生、存活和生长具有强大的筛选作用,在很大程度上决定着其天然更新的成败。研究1 a生天然更新幼苗存活数量和功能性状与微生境因子之间的关系,对环境筛(environmental filter)在天山云杉幼苗定居阶段筛选有重要意义。【前人研究进展】森林更新过程中幼苗定居阶段是最敏感、脆弱的阶段[1],幼苗的更新受到控制其发生和存活的众多生物和非生物微生境因子的影响,海拔高度、枯落物种类和厚度、苔藓厚度、草本盖度[2-6]以及相邻的成年个体[7]等,这些微生境因子在森林中的异质性普遍而又显著,且在时间和空间上都有体现。幼苗发生后在不同微生境因子的作用下,功能性状的表达出现变异,该种变异不仅潜在地促进了幼苗与其生境的适应,更重要的是通过功能性状变异驱动了其与周围生物生态位的分化[8]。土壤和光照条件局部尺度的异质性,导致幼苗功能性状上的变异,尽管幼苗的功能性状变异对生境异质的优化能力较弱,但仍能对幼苗的存活和生长起到正向的作用[9]。在不同的土壤、光照条件下,幼苗的根干物质比例(root dry mass fraction,RMF)、比叶面积(specific leaf area,SLA)都有不同程度的差异,且这些差异与幼苗的存活有着紧密的关联[10]。【本研究切入点】对于天山云杉的天然更新问题,研究往往着眼于化感物质的自毒作用[11-12]和土壤、海拔[13]等宏观环境因子对其天然更新成败的影响,有关天然更新幼苗存活数量和功能性状与微生境因子之间关系的研究却鲜有报道。需分析天山云杉1 a生天然更新幼苗存活数量和功能性状与微生境因子的关系,并结合微生境和功能性状分析1 a生天然更新幼苗存活数量与微生境的关系。【拟解决的关键问题】解析影响天山云杉1 a生幼苗存活数量和功能性状的关键微生境因子,并分析不同微生境条件下1 a生天然更新幼苗存活数量和功能性状的变化。
1 材料与方法
1.1 材 料
研究数据采集于新疆农业大学实习林场1林班(N 43°20′~43°22′,E 86°46′~86°49′;海拔1 800~2 500 m;面积485.0 hm2)。新疆农业大学实习林场(N 43°16′~43°26′,E 86°46′~86°57′)地处天山北麓乌鲁木齐市西南方向110 km的头屯河上游,隶属昌吉市辖区,东西长约15.0 km,南北长约17.5 km,面积10 043.5 hm2(其中有林地面积5 455.7 hm2),森林覆盖率56.45%;地势南高北低,海拔高度在1 700~3 200 m,坡度多在10~40°。林区地处欧亚大陆腹地,属温带大陆性冷凉山区气候,在天山中部林区具有典型性和代表性,年均降水量约为600 mm,60%集中在5~8月的雨季,7月空气相对湿度约为-65%;年均气温约为3℃,1月平均温度约为-10℃,7月平均温度约为14℃,年日照时长大于1 300 h,无霜期约为140 d。森林以天山云杉纯林为主,主要分布在海拔2 750 m以下的北、东北和西北坡,土壤为山地灰褐色森林土。林下的维管束植物高度多在3~20 cm,代表性物种有森林草莓(Fragariaveaca)、珠芽蓼(Polygonumviviparum)、圆叶鹿蹄草(Pyrolarotundifolia)、林地早熟禾(Poanemoralis)和天蓝岩苣(Cicerbitaazurea)等;林下常见的苔藓类植物有拟垂枝藓(Rhytidiadelphustriquetrus)、羽藓(Thuidiumtamariscinum)、塔藓(Hylocomiumsplendens)、牛毛藓(Ditrichumheteromallum)和扭口藓(Barbulaunguiculata)等。
1.2 方 法
1.2.1 试验设计
采用典型选样的方法,于2020年9月上旬,在新疆农业大学实习林场1班全域范围内,依据不同微生境条件下天山云杉的天然更新状况,共设置大小为1 m×1 m典型样方100个,样方中心距离林缘至少50 m。
于2020年9月下旬幼苗封顶停止生长后,逐一查数典型样方内1 a生幼苗存活数量,并在每个样方中随机挖取根、叶完整幼苗10株作为样株,标记后放入纸制采样袋带回实验室分析。
1.2.2 微生境因子获取
微生境地形因子包括海拔(elevation,Ele)、坡向(aspect,As)、坡度(slope,Slo)和坡位(slope position,Sp)。用GPS记录样方经纬度和海拔高度,用罗盘仪测量坡度和坡向,用地形图确定坡位。微生境非地形因子包括草本盖度(herbaceous coverage,Hc)、草本高度(herbaceous height,Hh)、苔藓盖度(moss coverage,Mc)、苔藓厚度(moss thickness,Mt)、枯落物盖度(litter coverage,Lc)、枯落物厚度(litter thickness,Lt)、腐殖质层厚度(humus thickness,Ht)和根系盘结层厚度(root system twine layer thickness,Rt),以及大树邻体效应(adult neighbor effect,Ane),即:以样方为中心一定半径范围(5 m)内的同种和异种大树胸高断面积之和[14]。其中:Hc、Mc和Lc采用网格(10 cm×10 cm)比例法测量;其余微生境非地形因子采用测量工具直接测定。表1
1.2.3 幼苗基础形态特征和干重获取
对带回实验室的天山云杉1 a生天然更新幼苗样株,分别样方以株为单位首先对其叶、茎、根进行分离,使用V750 PRO扫描仪(日本 EPSON)对分离的根、茎、叶扫描得到图像,采用根系形态与结构系统(WinRHIZO 2009a)分析扫描得到的图像,获得根、茎、叶基础形态特征指标值,包括:长度(length,Len;cm)、表面积(superficial area,Sa;cm2)、投影面积(projected area,Pa;cm2)、体积(volume,Vol;cm3)和平均直径(average diameter,Ad;cm)。将扫描后的根、茎放在烘箱(80℃)内烘至恒质量,用电子天平(0.001 g)称量根、茎干重(g)。每一样方单株幼苗基础形态特征指标值和干重的算术平均值作为样方幼苗基础形态特征指标值和干重的指标值。表1
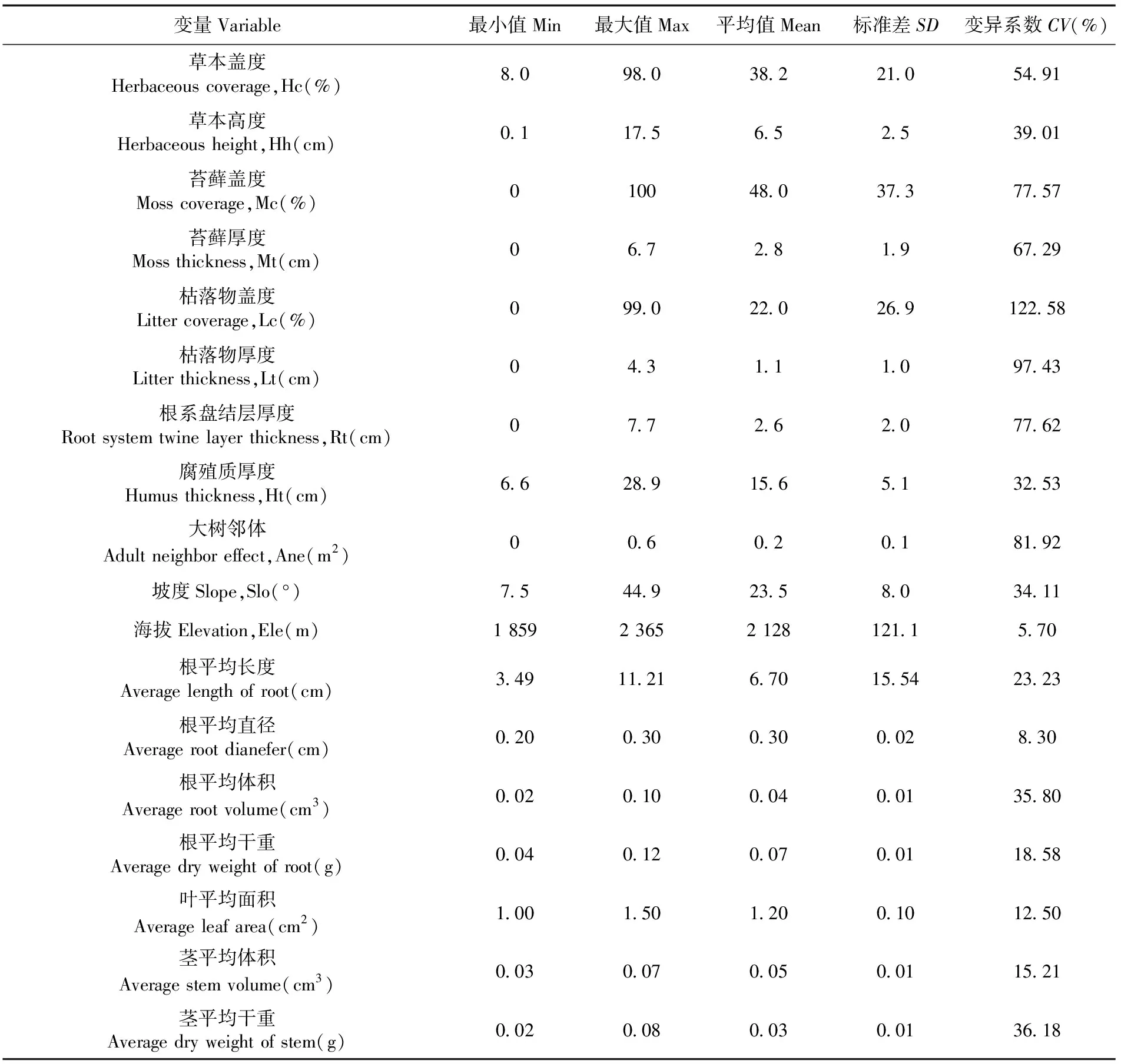
表1 样方数据基本Table 1 Basic situation of sample data
1.3 数据处理
1.3.1 幼苗功能性状指标
选取7种功能性状指标作为天山云杉1 a生天然更新幼苗的功能性状指标,即:叶面积(leaf area,LA;cm)、根直径(root diameter,RD;cm),比茎密度(specific stem density,SSD;g/cm3)、根茎比(ratio of root and stem,RRS)、比根长度(specific root length,SRL;cm/g)、根长密度(root Length Density,RLD;cm/cm3)和根组织密度(root tissue density,RTD;g/cm3)。其中,LA和RD直接由根系形态与结构分析系统获取的指标值得出;其余5个基于幼苗基础形态特征的功能性状指标计算公式如下:
1.3.2 冗余分析
冗余分析(redundancy analysis,RDA)的响应变量由100个样方的1 a生天然更新幼苗存活数量和7种功能性状指标(LA,RD,SSD,RRS,SRL,RLD,RTD)组成,解释变量数据由100个样方的4个微生境地形因子(Ele,As,Alo,Sp)和9个微生境非地形因子(Hc,Hh,Mc,Mt,Lc,Lt,Ht,Rt,Ane)组成,共包括13个因子。Sp为定性变量,分为下坡(bottom)、中坡(middle)和上坡(upper)。在数据进行标准化处理前,As需转换,即将0~360°的罗盘测量方位角(As)转换为0~1的坡向指标值(transformation of aspect,TRASP)[15],转换公式如下:
式中,TRASP为坡向指标值;As为坡向方位角角度。
通过转换,TRASP的值变化在0~1,0代表北北东方向,1代表南南西方向,即值越大表示微生境越干热。
在进行冗余分析前,对解释变量、响应变量数据进行标准化处理,消除不同变量量纲之间的差异带来的影响。对响应变量数据先进行除趋势对应分析(detrended correspondence analysis,DCA),结果得DCA第一轴梯度长度=1.123<3SD(standard deviation,SD)。先对微生境因子构成的解释变量进行前向选择,进行冗余分析(RDA),并对全部典范轴、每个单一典范轴和解释变量进行置换检验。采用层次分割方法(hierarchical partitioning)[16]对冗余分析的结果进行变差分解,以获取每个解释变量的校正解释率。
数据整理、计算采用Microsoft Excel软件;统计分析计算采用R软件;DCA、RDA采用vegan程序包,变差分解采用rdacca.hp程序包。
2 结果与分析
2.1 微生境因子与RDA排序轴的相关性
研究表明,筛选得到7个微生境因子(Hc、Ane、Lt、Mt、Rt、Ele、Sp),7个微生境因子的校正解释率(47.43%)能接近全模型的校正解释率(48.51%)。冗余分析得到前4个典范轴(RDA1、RDA2、RDA3和RDA4)的特征根。RDA1和RDA2的特征值分别为0.0837和0.0442,方差解释率分别为24.09%和12.28%。RDA1、RDA2和全部7个典范轴合计所代表的解释变量均与1 a生天然更新幼苗存活数量、幼苗功能性状指标之间均呈极显著相关(P<0.01)(RDA1:F=45.36,P=0.001;RDA2:F=18.51,P=0.001;7轴合计:F=12.1650,P=0.001),前向选择筛选出的7个微生境因子均对1 a生天然更新幼苗存活数量和功能性状存在极显著影响,其中前4个典范轴的累计方差解释率为52.24%。表2
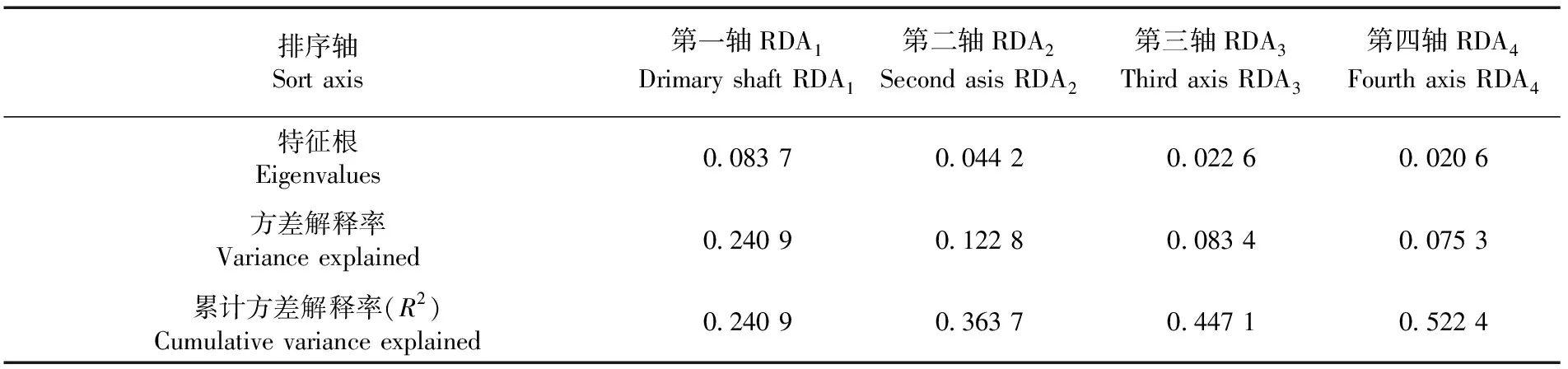
表2 RDA排序轴的特征根及其对1 a生天然更新幼苗 存活数量和功能性状的方差解释率Table 2 The characteristic roots of the RDA ordination axis and their variance explanation rates for the survival number and functional traits of 1 year old natural regeneration seedlings
Mt与RDA1呈极显著正相关(P<0.001),相关系数为0.923 0,Lt、Ane与RDA1呈极显著负相关(P<0.001),相关系数分别为-0.987 4和-0.957 6,沿RDA1从左到右,Lt、Ane逐渐降低,Mt逐渐增加。Ele、Hc、Rt与RDA2呈极显著负相关(P<0.01),相关系数分别为-0.832 2、-0.950 4和-0.770 7,沿RDA2自下而上,Hc、Ele和Rt逐渐降低。微生环境因子中的Mt、Rt、Ele、Hc对天山云杉1 a生天然更新幼苗的存活数量和功能性状的变异起着较大的作用。表3
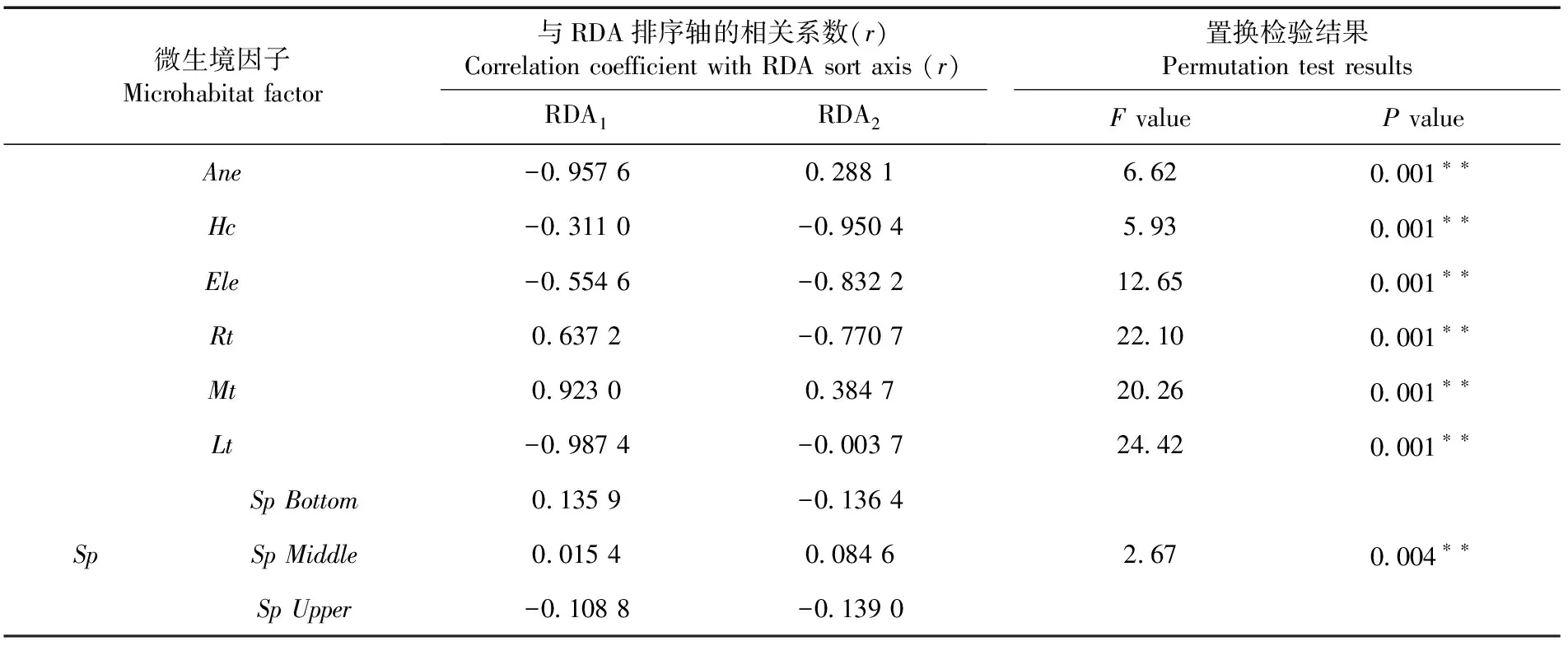
表3 1 a生天然更新幼苗微生境因子与RDA微环境排序轴的相关性及置换检验Table 3 Correlation between microenvironment factors and RDA micro-environment ordination axis and permutation test results of 1 a natural regeneration seedlings
Notes:*significant correlation (P<0.05);**highly significant correlation (P<0.01)
2.2 1 a生天然更新幼苗存活数量和功能性状与微生境因子的RDA排序
研究表明,LA、RLD箭头连线较短,与排序轴相关(拟合)程度较低。根功能性状中,RRS与Mt、Rt呈正相关,与Lt有着较强的负相关。Rd与Lt、Ele、Hc呈正相关,与Mt呈正相关。SRL、RTD与Rt、Hc呈正相关,与Lt呈负相关。SSD与Mt呈正相关,与Ele呈负相关。1 a生天然更新幼苗存活数量则与Mt有着较强的正相关,与Lt呈负相关(1 a生天然更新幼苗与幼苗功能性状用红色箭头表示,微生境因子用蓝色箭头表示。箭头连线的程长代表相关程度,箭头之间的夹角代表相关性,大于90°呈负相关,小于90°呈正相关;定性变量的形心垂直投影到响应变量的箭头的延长线上,投影点近似于该响应变量的数值沿着变量的位置)。图1
2.3 1 a生天然更新幼苗样方RDA排序
研究表明,可以把第一象限概括为“苔藓较厚、草本盖度较低、枯落物厚度较薄、海拔较低、根系盘结层较薄和大树邻体效应较弱”的微生境;第二象限概括为“枯落物厚度较厚、苔藓层较薄、草本盖度低、根系盘结层较薄和大树邻体效应较强”的微生境;第三象限概括为“枯落物厚度较厚、草本盖度较高、苔藓层较薄、根系盘结层较厚、海拔较高和大树邻体效应较强”的微生境;第四象限概括为“苔藓层较厚、草本盖度较高、枯落物厚度较低、根系盘结层较厚、海拔较高和大树邻体效应较弱”的微生境。第一象限(高SSD和低RD)的幼苗存活数量最多;第二象限(低RTD、低SRL和低RRS)的幼苗存活数量较低;第三象限(高RD和低SSD)的幼苗存活数量最低;第四象限(高RTD、高SRL和高RRS)的幼苗存活数量居中。图2
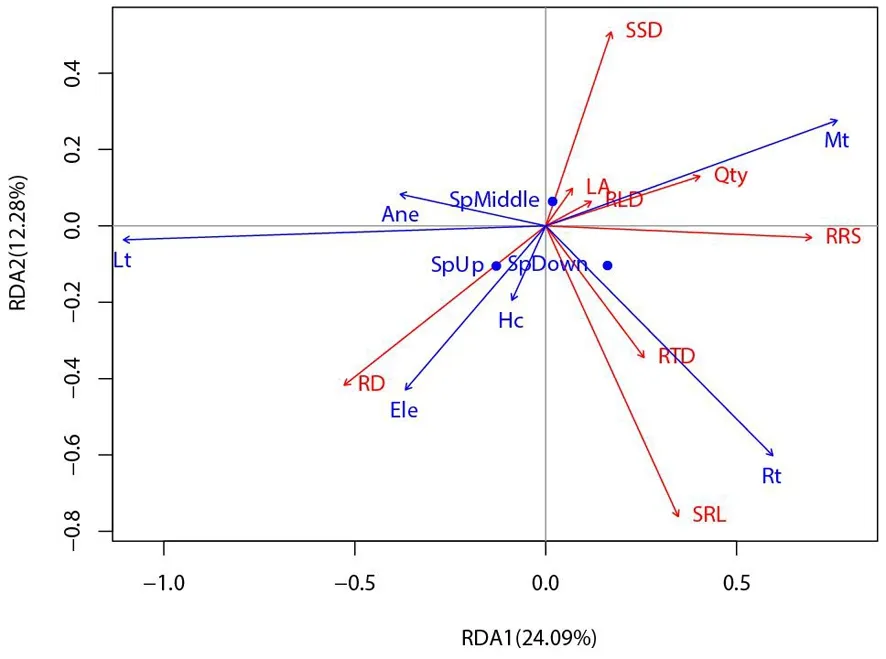
注(Note):草本盖度:Hc; 苔藓厚度:Mt; 枯落物厚度:Lt; 根系盘结层厚度:Rt; 海拔:Ele;大树邻体效应 :Ane; 坡下:Sp Bottom; 坡中:Sp Middle; 坡上:Sp Upper; 比茎密度:SSD; 叶面积:LA; 根直径:RD; 比根长:SRL; 根茎比:RRS; 根长密度:RLD; 根组织密度:RTD; 1 a幼苗存活数量:Qty,下同,the same as below
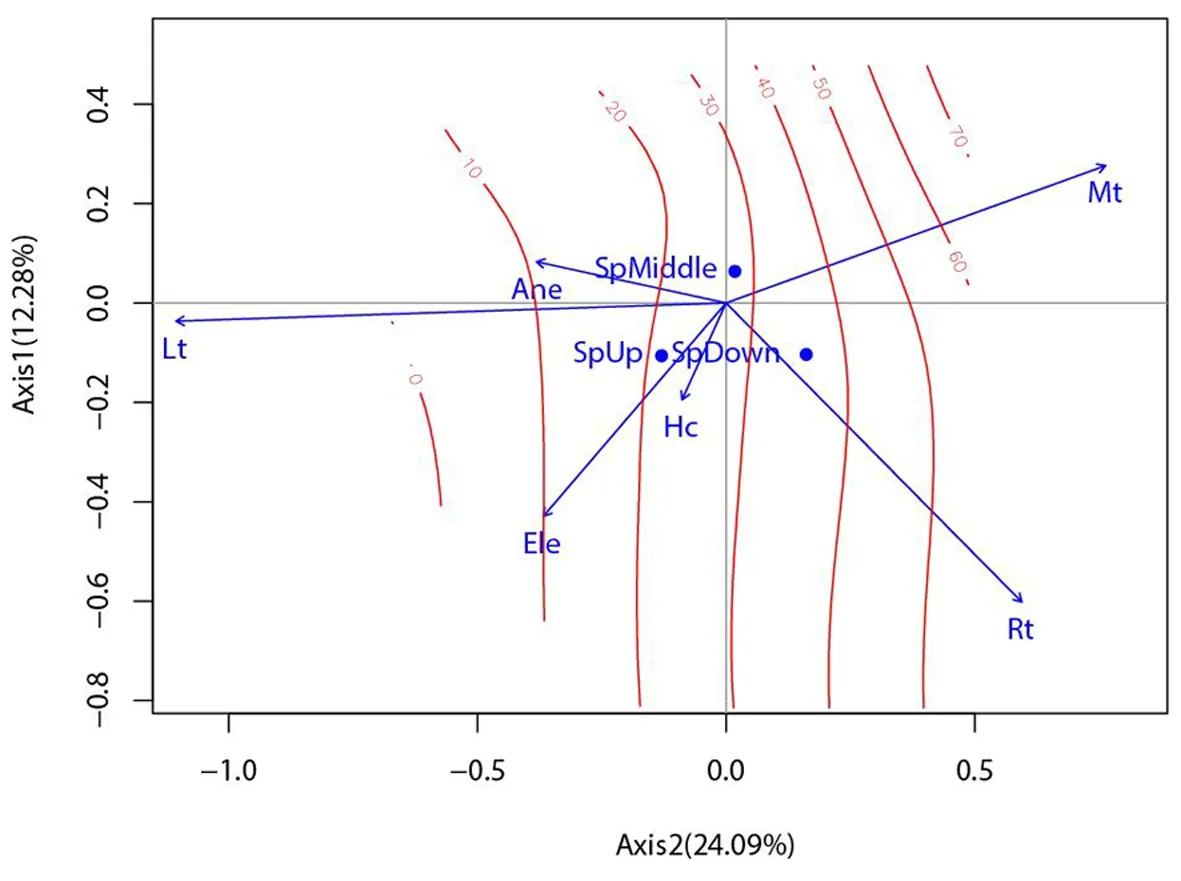
图2 1 a生天然更新幼苗存活数量等值密度拟合曲线-微生境因子的双序轴
2.4 微生境因子的变差分解
研究表明,微生境因子能够解释1 a生天然更新幼苗存活数量和功能性状变异的47.43%,其中林下地被物Lt、Mt和Hc的解释率可占到总解释率的58.09%,Rh和Ele的解释率的占比分别为20.64%和12.54%,Sp和Ane解释率的占比分别为5.92%和2.80%。表4
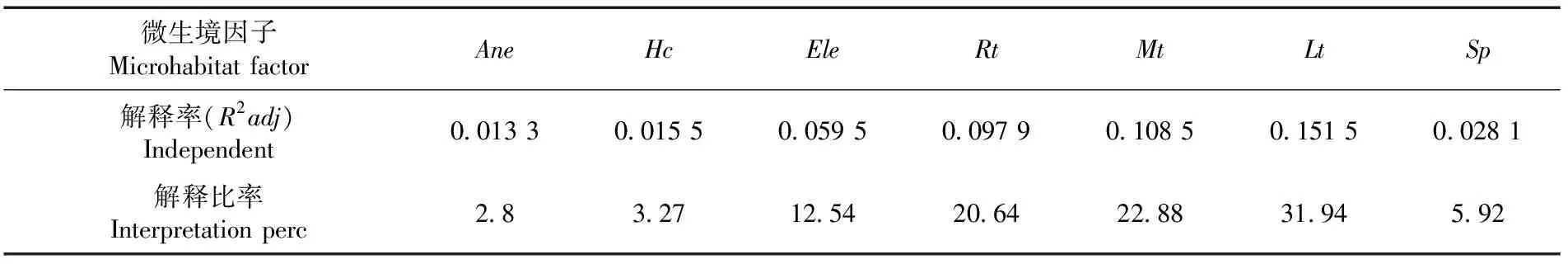
表4 RDA生境因子的变差分解Table 4 Variability decomposition RDA of redundant analysis habitat factors
3 讨 论
3.1 天山云杉1 a生天然更新幼苗功能性状变异的微生境解释
1 a生天然更新幼苗的RRS、RD和SRL与根系盘结层、枯落物、苔藓密切相关。其中苔藓、根系盘结层厚度对RRS和SRL有着正向的影响。在相关的研究中也发现,在苔藓上更新的幼苗有着更细更长的根和较高的根茎比[17-18]。幼苗根茎比的变化体现幼苗资源对地上地下部分的分配策略。苔藓作为耐荫型植物,幼苗在苔藓上发生时不必为光资源进行竞争,其对资源的竞争更多地表现在地下部分。虽然苔藓能够为种子提供良好的萌发条件,但是种子萌发后更重要的是扎根到土壤殖质中持续地获取营养,适应出细长根的性状,从而保证幼苗的存活。SRL可以看作植物对土壤营养变化的反应指标,在土壤养分匮乏、地下资源竞争剧烈的条件下,幼苗的SRL会增大[19]。研究发现,往往在根系盘结层较厚的微生境,幼苗会拥有较高的SRL和RRS,在地下竞争剧烈的微生境中,幼苗存活策略的重心在地下部分,做出扎深根增强养分吸收能力的对策,来对抗地下的竞争。1 a生天然更新幼苗的RD则与枯落物厚度表现出正相关,在枯落物为主的地被物微生境中,没有像苔藓一样较好的保水性,总体呈现较为干燥的状态。较高的RD体现了较强的水分运输能力[20],是幼苗根系在干燥生境中的适应表现。幼苗的SSD与植物种的形态、机械和生理生态特性相关[21],较高的SSD能更好地维持幼苗机械结构。研究中,在1 850~2 350 m海拔范围内,1 a生天然更新幼苗的SDD随着海拔的上升而下降,幼苗的生长速率加快。天山云杉幼苗比茎密度随海拔高度的上升而下降[22],与研究结果基本一致。天山云杉1 a生天然更新幼苗对其自身生物量的分配及功能性状之间的差异,反映了其在地理和微生境尺度上对自身所需资源有效性的调整。
3.2 天山云杉1 a生天然更新幼苗存活数量变异的微生境解释
通过天山云杉1 a生天然更新幼苗存活数量和功能性状与微生境因子的RDA排序,以及1 a生天然更新幼苗样方的RDA排序与幼苗存活数量的等密度拟合曲线得到, 1 a生天然更新幼苗的存活数量主要沿着Axis1在变化,与Axis2基本平行,故在此仅讨论Axis1上幼苗存活数量与微生境因子和功能性状的相互影响。RDA1的基本情况为:自左向右,地被物由枯落物逐渐转变为苔藓,大树邻体效应逐渐降低,天山云杉1 a生天然更新幼苗存活数量由少变多。
Axis1右半轴的第一和第四象限代表了以苔藓为主的地被物微生境,幼苗功能性状表现为高RRS、高SRL、高RTD和低RD。从地被物类型来看,苔藓具有良好的保水、保温性,不仅能够改善周围的微气候[23-24],更重要的是其粗糙的表面还能够减少附近的空气流动,有助于种子的滞留。天山云杉种子一方面在苔藓层上容易得到积累和保留,另一方面苔藓层也能够长时间为种子萌发提供稳定有利的温度和湿度,使得天山云杉天然更新幼苗在融雪后的5月下旬至6月中旬的雨季初期大量发生,这种有利的微生境条件还为幼苗后续的存活提供了条件,在以往的研究中也发现苔藓对树木幼苗的天然更新与存活有促进作用[25]。在以苔藓为主的地被物微生境中,天然更新幼苗的主根易于伸长,其具有更长和相对更细的根系,故SRL和RTD较高,生长季根系吸收水分和养分的能力也较强。在Axis1右半轴的微生境中,一方面苔藓层能使天山云杉天然更新幼苗大量发生,且后续还能为其提供有利的存活条件;另一方面,幼苗适应苔藓层生境,功能性状上发育为较为发达的根系系统,具有较强的水分和养分吸收能力。
Axis1左半轴的第二和第三象限代表了以枯落物为主的地被物微生境,天山云杉1 a生天然更新幼苗的功能性状表现为低RRS、低SRL、低RTD和高RD。枯落物能够通过多个方面影响幼苗发生和存活。地表较厚的枯落物(以天山云杉凋落针叶为主)是天山云杉林土壤种子库中天山云杉种子分布的主要垂直空间[26-27],但其也阻挡种子萌发后幼苗根系与土壤的接触。枯落物是类似天山云杉(千粒重不足10.0 g)小种子物种幼苗发生和建成的主要障碍[28]。天山云杉的针叶及其凋落的针叶中含有水溶性化感自毒物质3,4-二羟基苯乙酮(3,4-dihydroxyacetophenone),其通过春季融雪和夏季降雨淋溶大量释放后,不仅能够显著抑制天山云杉种子萌发,使种子在吸水后的枯落物生境中霉变失活,而且还能够显著抑制天山云杉幼苗胚芽、胚根的生长和鲜重的增长[29-30],成为阻碍天山云杉天然更新幼苗发生和建成不易察觉的隐形障碍。在较干燥的生境中,根系往往具有较强的耐脱水性和吸水性,在功能性状上具体表现为有较低的SRL和较粗的根系[31]。在Axis1左半轴上,大树邻体效应(Ane)也在增强,很明显这种效应也抑制了天山云杉幼苗的更新和存活。在地被物为枯落物的微生境中,枯落物通过融雪和降雨淋溶释放的化感自毒物质产生的自毒作用,一方面抑制了天山云杉种子的萌发及其幼苗根系的生长,另一方面较为干燥和缺乏养分的枯落物微生境也不利于天山云杉幼苗根系的生长和发育。
幼苗时期是天山云杉极其脆弱的阶段,在不同微生境中1 a生天然更新幼苗存活数量差异较大,受到环境的筛选作用强烈,尤其是林下地被物的类型对天山云杉1 a生天然更新幼苗有较强的筛选作用,同时环境筛的作用也对幼苗的功能性状表达施加了压力。幼苗在不同微生境中存活数量的变化和功能性状的变异,体现了幼苗面对环境筛筛选所带来的各种资源不平衡或短缺时,对其自身资源的分配策略和微生境中幼苗发生策略以及幼苗发生后微生境所影响的功能性状对幼苗存活的影响。研究中,天山云杉1 a生天然更新幼苗存活数量和功能性状变异微生境因子未能解释的部分占52.57%,可能来源于一些调查中没有考虑到的微生境因子影响,如土壤养分、微生物和光照条件等。此外,在此需要特别指出的是,天山云杉1 a生天然更新幼苗在冬季和次年春季死亡率极高[32],哪些微生境因子和功能性状有助于幼苗渡过寒冷严酷的冬季和冷热多变的初春,也有待在接下来的研究中做进一步深入分析。
4 结 论
天山云杉林下微生境的高度异质化影响了天山云杉1 a生天然更新幼苗的存活数量和功能性状。筛选出了草本盖度、苔藓厚度、枯落物厚度、根系盘结层厚度、大树邻体效应、海拔和坡位7个能显著影响天山云杉1 a生天然更新幼苗存活数量和功能性状变异的微生境因子。微生境因子能解释天山云杉1 a生天然更新幼苗存活数量和功能性状变异的47.43%。其中,由苔藓厚度、枯落物厚度和草本盖度构成的林下地被物的解释率可占到总解释率的58.09%;根系盘结层厚度和海拔的解释率占比分别为20.64%和12.54%。
MicrohabitatInterpretationofSurvivalQuantityandFunctionalTraitsof
One-year-oldNaturalRegenerationSeedlingsofPiceaschrenkianavar.
猜你喜欢
纺织学报(2022年8期)2022-08-26
军事文摘(2021年18期)2021-12-02
润·文摘(2021年11期)2021-11-20
现代农村科技(2020年7期)2020-12-19
意林·全彩Color(2019年11期)2019-12-30
儿童故事画报·自然探秘(2017年7期)2018-03-14
启迪与智慧·教育版(2018年12期)2018-02-26
山西林业科技(2017年4期)2018-01-23
民族音乐(2016年4期)2016-12-06
甘肃林业(2016年2期)2016-11-07
