
二化螟对双酰胺类杀虫剂抗性的分子机制研究进展
2023-04-18 03:39刘宝生徐德进徐广春张亚楠张月亮黄水金吴顺凡高聪芬
农药学学报 2023年2期
徐 鹿, 孙 杨, 刘宝生, 徐德进, 徐广春, 赵 钧, 张亚楠, 张月亮, 黄水金, 吴顺凡, 高聪芬*,
(1.江苏省农业科学院 植物保护研究所,省部共建国家重点实验室培育基地-江苏省食品质量安全重点实验室,南京210014;2.南京农业大学 植物保护学院,南京 210095;3.江西省农业科学院 植物保护研究所,南昌 330200;4.河南省农业科学院 烟草研究所,河南 许昌 461000;5.淮北师范大学 生命科学学院,安徽 淮北 235000)
双酰胺类杀虫剂具有高杀虫活性和阻止取食速度快的毒理学特征[1],其作用机制主要是与鱼尼丁受体 (ryanodine receptor,RyR) 结合,激活钙离子通道,使钙库中的钙离子快速释放,钙离子与肌钙蛋白结合引起肌肉纤维持续性收缩,导致昆虫瘫痪并停止取食,最终饥饿致死;同时该类杀虫剂对非靶标生物安全且与常用杀虫剂无交互抗性[2]。基于双酰胺类杀虫剂的大面积应用带来了巨大的经济效益,因而合成开发这类化合物成为杀虫剂研发的热点之一。双酰胺类杀虫剂已有3 类11 种产品 (表1):第1 类是邻苯二甲酰胺类,主要有氟苯虫酰胺和氯氟氰虫酰胺;第2 类是邻甲酰胺基苯甲酰胺 (硫代) 类,包括氯虫苯甲酰胺、溴氰虫酰胺、四氯虫酰胺、环丙虫酰胺、环溴虫酰胺、四唑虫酰胺和硫虫酰胺7 种;第3 类为间甲酰胺基苯甲酰胺类,主要有溴虫氟苯双酰胺和环丙氟虫胺[3],与第1、第2 类产品的作用靶标RyR 不同,其作用靶标是γ-氨基丁酸受体 (gammaaminobutyric acid receptor,GABAR)。双酰胺类杀虫剂为防治鳞翅目害虫提供了新型的化学手段,但抗药性问题不可避免,多种鳞翅目害虫已对其产生不同水平的抗性[1],导致其田间防效下降、用量增加。
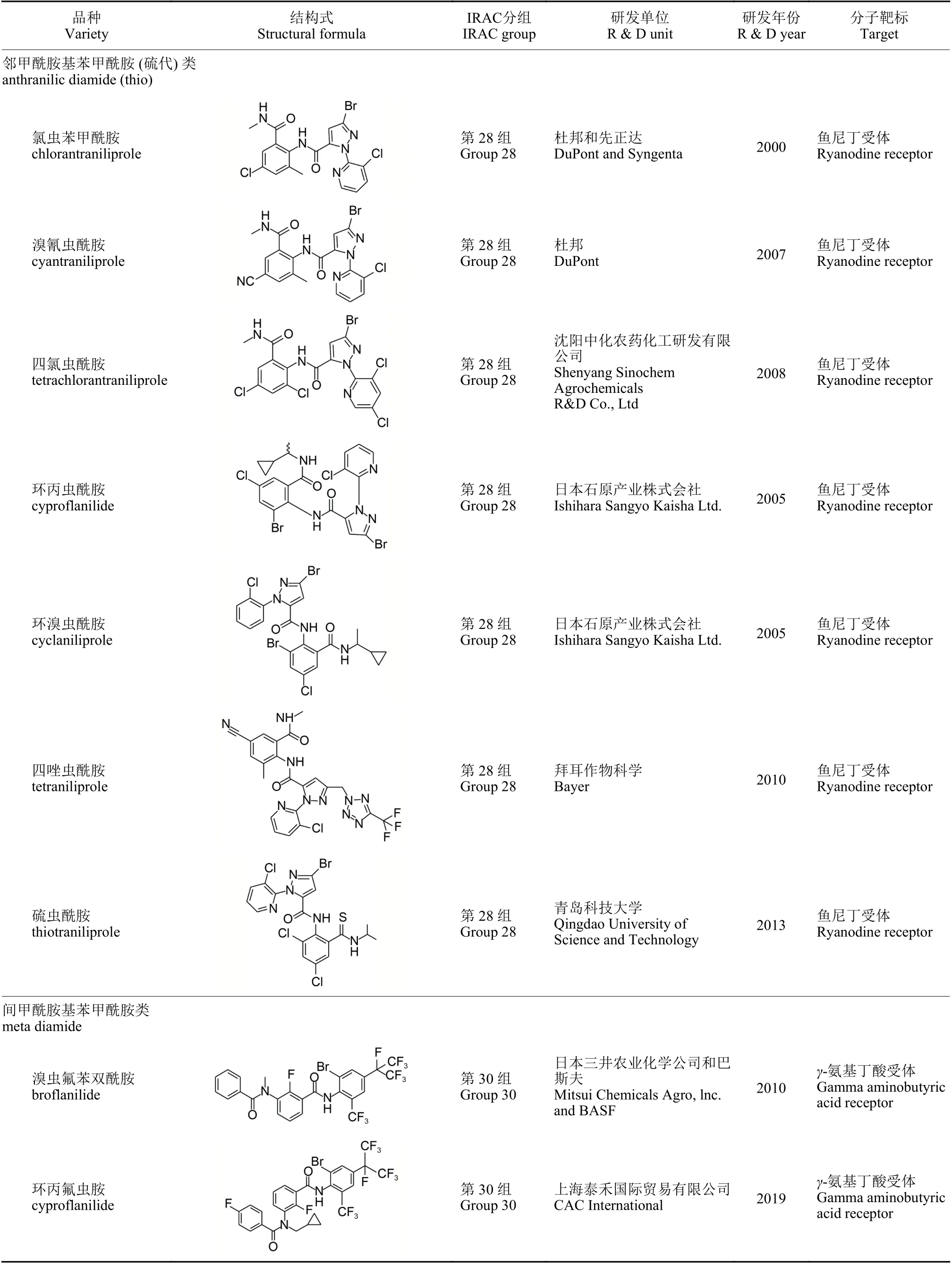
续表1Table 1 (Continued)
二化螟Chilo suppressalis(Walker) 属鳞翅目草螟科,是一种危害水稻生产的重要害虫,分布于亚洲、中东和南欧地区,尤以中国长江流域及以南稻区危害最重,由于杂交稻的推广和耕作制度的变革,二化螟的发生量逐年增加,可造成水稻枯心、枯鞘、枯孕穗和白穗,严重时造成显著减产[4]。据统计,中国二化螟年发生面积为1400 万公顷次,经济损失达115 亿元[5],这与二化螟的抗药性存在直接联系。中国已登记上市的双酰胺类杀虫剂产品中,可用于防治二化螟等水稻螟虫的主要有氯虫苯甲酰胺、溴氰虫酰胺和四唑虫酰胺。双酰胺类杀虫剂经过10 多年的持续使用,二化螟对该类杀虫剂已进化出抗性,其内在基础是抗性基因频率的上升,当高频率的抗性基因在种群中稳定表达时,即可形成高水平抗性种群,导致常规剂量的杀虫剂对二化螟的防治效果下降[6]。因此,研究二化螟对双酰胺类杀虫剂产生抗性的分子机制,可为双酰胺类杀虫剂的合理使用及新型杀虫剂研发提供参考。
害虫对双酰胺类杀虫剂的抗性机制主要涉及代谢抗性和靶标抗性。代谢抗性揭示双酰胺类杀虫剂到达分子靶标之前如何被代谢酶降解和转运,靶标基因位点突变介导的靶标抗性揭示双酰胺类杀虫剂如何发挥致死效应[7]。基于双酰胺类杀虫剂抗性机制的分子检测技术,可以预测二化螟对该类药剂抗性的发生和扩散。本文综述了二化螟对氯虫苯甲酰胺、氟苯虫酰胺和溴氰虫酰胺的抗性现状及氯虫苯甲酰胺抗性分子机制的最新研究成果,分析存在的问题,并展望其未来发展方向,旨在为二化螟的抗药性监测和治理以及双酰胺类杀虫剂科学使用与新品种研发提供依据。
1 2010—2020 年间二化螟对双酰胺类杀虫剂的抗性现状
抗药性监测结果反映了害虫对杀虫剂的抗性水平及地理分布,可为指导田间精准用药、有效治理抗药性害虫提供依据。
据文献报道,有关二化螟对双酰胺类杀虫剂的抗性监测涉及中国江西、湖南、浙江、湖北、安徽、江苏、四川、山东和河南9 省共94 个田间种群[8-21]。氯虫苯甲酰胺于2008 年在中国登记用于防治二化螟,2010 年即监测到浙江瑞安、湖北荆州、安徽宣城和黄山以及江苏邗江种群对其产生低水平抗性[8],而江苏仪征、安徽庐江、浙江龙游和湖北武穴种群对其产生中等水平抗性[9];氟苯虫酰胺与氯虫苯甲酰胺在同一年获得登记,2011年监测到湖南东安、浙江象山、湖北武穴、江西上高、江苏仪征种群对其产生低水平抗性[10]。由此可见,二化螟在2~3 年内即可表现出对双酰胺类杀虫剂的抗性。2013—2015 年间,监测到江西南城、浙江象山、湖北监利和安徽庐江二化螟种群对氯虫苯甲酰胺产生中等水平抗性,浙江苍南和余姚种群对其产生高水平抗性[11-12];江西上高和南城、湖南东安和攸县、浙江金华、余姚和象山、湖北黄冈、荆州、监利和孝感、安徽庐江和潜山种群对氟苯虫酰胺产生中等水平抗性,浙江苍南种群对其产生高水平抗性[13]。与2013—2015年间相比,2016—2018 年间的江西、浙江和湖南田间种群的抗性倍数显著上升,安徽和湖北监测种群的抗性倍数增加[14-18],到2019—2020 年间,江西、浙江、湖南、安徽监测种群的抗性倍数继续显著上升[19-21],但湖北种群的抗性倍数与2016—2018 年间的无显著差别 (图1)。

图1 2010—2020 年间二化螟对双酰胺类杀虫剂的抗性Fig.1 Resistance of Chilo suppressalis to diamide insecticides during 2010-2020
综上所述:目前,二化螟种群对氯虫苯甲酰胺的高水平抗性区域为浙江东部沿海地区、安徽沿江地区、江西环鄱阳湖地区和湖南中南部地区;中等水平抗性区域为湖北部分稻区;敏感区域为江苏、四川和山东地区。二化螟对氟苯虫酰胺的高水平抗性区域为浙江东部沿海地区;中等水平抗性区域为江西环鄱阳湖地区、湖南南部地区和湖北东南地区;低水平抗性区域为安徽沿江地区;敏感区域为江苏、四川、河南和山东地区。二化螟对溴氰虫酰胺的中等水平抗性区域为江西南昌地区和湖北武穴地区;低水平抗性区域为湖南宁乡地区和湖北赤壁地区;敏感区域包括安徽、江苏、四川、河南和山东地区。
2 二化螟对双酰胺类杀虫剂的代谢抗性机制
代谢抗性是解毒代谢增强导致害虫对杀虫剂产生的抗性,细胞色素P450 单加氧酶 (cytochrome P450 monooxygenases,P450s)、酯酶 (esterases,ESTs)、谷胱甘肽S-转移酶 (glutathioneS-transferases,GSTs)、尿苷二磷酸糖基转移酶 (uridine diphosphate glycosyltransferases, UGTs) 和三磷酸腺苷结合盒(adenosine triphosphate binding cassette,ABC) 转运蛋白参与其中。这些酶及ABC 转运蛋白基因的表达上调、复制和突变是介导二化螟对双酰胺类杀虫剂代谢抗性的主要机制。
2.1 P450s 表达上调介导的氯虫苯甲酰胺的抗性
转录水平的上调表达导致二化螟蛋白质含量和酶活性增加是P450s 介导的二化螟代谢抗性的典型特征之一。Xu 等[22]对安徽省芜湖二化螟种群进行了25 代筛选,分别获得了氯虫苯甲酰胺的抗性品系WHR (82.37 倍) 和敏感品系WHS。与WHS 相比,WHR 中P450s 基因CYP6CV5、CYP9A68、CYP321F3和CYP324A12表达上调 (4.48~44.88倍),用致死中量的氯虫苯甲酰胺处理WHR 后,这4 个P450s 基因表现出显著的上调表达 (2.11~3.81 倍)。RNA 干扰 (RNAi) 试验结果表明,无论是分别干扰还是同时干扰,这4 个P450s 基因的表达均可显著增强二化螟对氯虫苯甲酰胺的敏感性。同时,WHR中P450s 酶的活性为WHS 的3.87 倍[22]。Zhao 等进一步分析了上述4 个P450s 基因在采自四川的5 个田间二化螟种群中的表达,发现CYP9A68、CYP321F3和CYP324A12在这5 个田间种群中均为过表达 (1.24~24.96 倍);酶活性和增效剂试验结果也证实P450s 酶参与了二化螟对氯虫苯甲酰胺抗性的形成[23]。上述结果表明,组成型P450s 基因的表达上调介导了二化螟种群对氯虫苯甲酰胺的代谢抗性。
2.2 UGTs 表达上调介导的氯虫苯甲酰胺抗性
UGTs 是一类多功能的解毒代谢酶,参与多种化学物质的代谢,在杀虫剂的代谢抗性中发挥作用。Zhao 等[24]用氯虫苯甲酰胺对采自浙江省余姚市的二化螟种群进行17 代筛选,获得了抗性为44.32倍的品系YYR。在24 个UGTs 基因中,CsUGT40AL1和CsUGT33AG3在YYR 中的表达分别是YYS 的12.36 和5.34 倍;采用RNAi 技术显著抑制这两个基因的表达后,二化螟幼虫对氯虫苯甲酰胺的敏感性显著增加;而UGTs 酶的抑制剂5-硝基脲嘧啶和硫吡唑酮可使氯虫苯甲酰胺对二化螟的毒力分别提高3.38 倍和3.56 倍。Li 等[25]研究发现,UGT2B17上调表达介导了小菜蛾对氯虫苯甲酰胺代谢机制。因此,组成型UGTs 过表达也参与了二化螟对氯虫苯甲酰胺的代谢抗性,同时UGTs 过表达在不同害虫对氯虫苯甲酰胺代谢抗性中具有普遍性。
2.3 ABC 转运蛋白基因表达上调介导的氯虫苯甲酰胺抗性
ABC 转运蛋白家族是后生动物最大的转运体基因家族,包括昆虫,其功能主要是将外源有毒物质排出细胞,与多重抗性相关。Peng 等[26]研究发现,二化螟ABC 转运蛋白基因CsABCA1、CsABCD2和CsABCH2在抗氯虫苯甲酰胺的品系R1、R2 和HR 中共同过表达。Meng 等通过亚致死剂量的氯虫苯甲酰胺处理来自江苏省扬州市的田间二化螟种群,发现CsABCC8、CsABCG1C和CsABCH1显著过表达;增效剂试验显示,维拉帕米显著增加了氯虫苯甲酰胺对二化螟的毒力[27]。上述表明,ABC 转运蛋白很可能也参与了二化螟对氯虫苯甲酰胺的代谢抗性。
3 二化螟对双酰胺类杀虫剂的靶标抗性机制
RyR 是双酰胺类杀虫剂的分子靶标,RyR 氨基酸突变导致靶标敏感性降低,引起双酰胺类杀虫剂对二化螟的药效下降,从而二化螟进化出对双酰胺类杀虫剂的靶标抗性。
3.1 二化螟鱼尼丁受体基因突变位点和突变频率
Sun 等室内经过30 代筛选,获得了二化螟抗氯虫苯甲酰胺R2 品系 (110.4 倍),其中的RyR 存在Y4676D 突变,突变频率为94.7%;2016 年采自江西省南昌市田间HR 品系 (249.6 倍) 的RyR存在Y4676C/D 和I4758M 突变,其突变频率分别为13.2%、13.2%和77.8%[28]。Huang 等[19]发现,2018 年采自江西省南昌市NCha18 品系的RyR 包含Y4667D、I4758M 和Y4891F共3 个突变,抗性倍数达到536.8 倍,其突变频率分别为13.9%、69.4%和5.55%。另外,来自江西省南城县的田间二化螟抗氯虫苯甲酰胺NChe 品系,从2017—2020 年间抗性倍数持续增加 (47.0~1076.8 倍),NChe17 和NChe18 的RyR 包含Y4667D、I4758M和Y4891F共3 个突变 (10.8%~57.2%)[19],而NChe19 和NChe20的RyR 包含Y4667D 和Y4891F 两个突变 (6.9%~93.4%)[20]。同样地,来自浙江省苍南县、浙江省余姚市和湖南省攸县的田间二化螟抗氯虫苯甲酰胺CN、YY 和YX 品系分别在2018—2020 年、2014—2020 年和2018—2020 年间抗性倍数持续升高 (158.7~311.9 倍、77.6~1426.8 倍和488.2~>1000 倍),其中CN 品系的RyR 仅有I4758M 单突变,其突变频率均大于90%;YY 品系的RyR 包含G4910E 和I4758M 单突变及Y4667C 和I4758M双突变,单突变频率为23.8%和95.5%,双突变频率介于33.3%~83.4%;YX 品系的RyR 包含Y4667D 和I4758M 双突变及Y4676C/D 和I4758M三突变,其双突变和三突变频率分别介于25%~67.6%和12.5%~33.4%之间[19-20]。来自浙江省瑞安市和浙江省象山县的田间二化螟抗氯虫苯甲酰胺RA 和XS 品系,除了RA18 品系的RyR 具有Y4667D 和I4758M 双突变(12.5%和100%)外,其余均仅有I4758M 单突变(72.4%~100%),RA 和XS 品系抗性倍数分别达到355.0~1614.4 倍和223.8~2087.5倍,处于高水平抗性[19-20]。对比湖南省桃江县和四川省富顺县的田间二化螟抗氯虫苯甲酰胺TJ17 和FS18 品系,两个品系的RyR 均有I4758M 单突变,但其突变频率具有显著差异 (96.2%和5.6%),TJ17 品系发展出高水平抗性,而FS18 品系处于低水平抗性阶段[19](表2)。
综上所述,二化螟RyR 羧基末端的第2、3 和4 跨膜区的4676D/C、I4758M和Y4891F 以及第4 和第5 跨膜区的胞外连接环上G4910E 和G4915E 的6 个突变已被发现 (图2),且突变频率与二化螟对氯虫苯酰胺的抗性水平相关,尽管有些突变频率较低,但RyR 的突变组合使二化螟获得了对氯虫苯甲酰胺的高水平抗性。
3.2 二化螟鱼尼丁受体基因突变位点功能分析
Huang 等[19]利用反向遗传学工具CRISPR/Cas9成功构建了8 个基因组水平的黑腹果蝇突变系,包括M4758I、Y4667D、Y4891F 和G4915E 共4 个单突变品系以及Y4667D 和M4758I、Y4667C和M4758I、Y4891F 和M4758I、G4915E 和M4758I共4 个双突变系。毒力测定结果显示,果蝇Y4667D突变系 (对应二化螟Y4667D 和I4758M 双突变)对氯虫苯甲酰胺、溴氰虫酰胺、四氯虫酰胺和氟苯虫酰胺的抗性倍数分别为1542.8、487.9、290.1和21.2 倍,果蝇M4758I 和Y4667D 双突变系 (对应二化螟Y4667D 单突变) 对这4 种杀虫剂仅产生了117、90、22 和6 倍抗性,果蝇Y4667C 突变系 (对应二化螟Y4667C 单突变) 仅对这4 种杀虫剂产生低水平抗性或保持敏感 (1.3~8.6 倍),但Huang等[20]构建的果蝇Y4667C 突变系 (对应二化螟I4758M 和Y4667C 双突变) 对以上4 种杀虫剂产生了中等至高水平抗性 (19.5~172.1 倍)。果蝇G4915E和M4758I 双突变系 (对应二化螟G4915E 突变) 对氯虫苯甲酰胺和溴氰虫酰胺产生了153.1 和323.5倍高水平抗性,对氟苯虫酰胺和四氯虫酰胺产生了28.9 和25.2 倍中等水平抗性,而果蝇Y4891F突变系对上述4 种双酰胺类杀虫剂均表现为低水平抗性。由此可见,经CRISPR/Cas9 技术进一步确认了RyR 基因双突变介导了二化螟对双酰胺类杀虫剂的高水平抗性,证实了RyR 突变在双酰胺类杀虫剂靶标抗性中的作用。
4 问题与展望
目前,二化螟对双酰胺类杀虫剂的抗性监测主要采用生物测定法,该方法对试虫、杀虫剂、饲养条件、试验条件和结果测定均有严格要求[29]。Mao 等[16]在2016—2018 年间监测到河南信阳田间二化螟种群对氯虫苯甲酰胺产生了4.3~5.8 倍抗性;而同时间内Lu 等[17]发现,河南信阳田间二化螟对氯虫苯甲酰胺产生了78.7 倍抗性。相同区域二化螟种群对同一药剂的抗性倍数不同,这可能是由于二化螟抗性种群的生测方法没有标准化造成的。因此,不同监测点需要采用标准化生物测定方法,并使用统一提供的杀虫剂样品,以避免相同区域内抗性测定结果间的差异[29]。二化螟RyR基因突变与其对杀虫剂的抗性具有相关性,新一代测序技术 (如扩增子检测) 为获取田间二化螟种群中携带的双酰胺类杀虫剂抗性基因及频率提供了快速手段[30]。因此,可将生物测定法与高通量分子检测方法相结合,预测害虫抗药性的发生和扩散,提前制定药剂使用方案。
二化螟对双酰胺类杀虫剂代谢抗性的形成是一个复杂的过程,有多种代谢酶和转运蛋白参与其中。Sun 等[28]发现,在抗性品系R1、R2 和HR 中P450s 和ESTs 的活性增强,P450s 抑制剂胡椒基丁醚和ESTs 抑制剂磷酸三苯酯显著提高了氯虫苯甲酰胺对二化螟的杀虫活性,CsABC表达上调且其过表达的基因数量随着抗性倍数的增加呈现减少的趋势[26],推断P450s、ESTs 和ABC 转运蛋白基因可能参与了R1、R2 和HR 品系的代谢抗性。在抗性品系WHR 中,P450s 基因CYP6CV5、CYP9A68、CYP321F3和CYP324A12过表达[22],而在抗性品系YYR 中,UGTs 基因CsUGT40AL1和CsUGT33AG3表达上调[24],原因在于来自不同地理种群的抗性品系具有不同的遗传背景,但参与二化螟对双酰胺类杀虫剂代谢抗性形成的解毒酶和转运蛋白功能需要进一步明确,GAL4/UAS表达系统和CRISPR/Cas9 基因编辑技术为验证其功能提供了有利工具,可精准地分析其在抗药性中的作用。另外,利用高通量测序的多组学技术,可以系统地鉴定双酰胺类杀虫剂的代谢抗性基因,例如酰胺酶等[2]。目前,参与二化螟对双酰胺类杀虫剂产生抗性的不同代谢酶的表达调控机制尚不清晰,除了应用经典遗传学鉴定顺式作用元件、转录因子和信号转导通路等外[31],还需关注非编码RNA[2]和昆虫共生菌[32]的调控作用。因此,解析二化螟对双酰胺类杀虫剂代谢抗性的分子机制是持续关注的重点和热点。
RyR 突变介导二化螟对双酰胺类杀虫剂的靶标抗性,其双突变使二化螟获得对双酰胺类杀虫剂的极高水平抗性。不同地理种群的二化螟RyR 突变数量和位点存在差异,这与当地二化螟种群遗传和用药历史相关。随着双酰胺类杀虫剂的推广应用,监测不同地理种群的二化螟中对双酰胺类杀虫剂抗性相关的靶标突变及其频率将是长期性的基础工作。双酰胺类杀虫剂是RyR 的激活剂,RyR 突变导致双酰胺类杀虫剂与RyR 的结合能力下降[2],这是产生靶标抗性的重要原因。因此,除了引入CRISPR/Cas9 技术对二化螟RyR 突变进行精准编辑外,还需要利用放射性配基结合和电生理技术分析突变对RyR 与双酰胺类杀虫剂结合的影响[20],全面证实二化螟RyR 突变在双酰胺类杀虫剂抗性中的功能。另外,Sun 等[28]发现,在R1、R2 和HR 中CsuRyR表达量下降,推测靶标基因表达量下降也可能导致二化螟对杀虫剂的敏感性下降,但二化螟中RyR 表达量改变是否介导对双酰胺类杀虫剂的抗性还需要进行功能分析。
田间二化螟种群中氯虫苯甲酰胺、氟苯虫酰胺、溴氰虫酰胺、四氯虫酰胺和氯氟氰虫酰胺之间存在交互抗性[33],交互抗性发生在二化螟抗氯虫苯甲酰胺的种群中,推测相同的抗性基因介导了对双酰胺类杀虫剂间的交互抗性。由于氟苯虫酰胺对水生生物高毒,因此我国于2016 年取消了其在水稻上的登记;四氯虫酰胺在我国水稻上登记用于稻纵卷叶螟的防治;氯氟氰虫酰胺、环丙虫酰胺、环溴虫酰胺、硫虫酰胺和溴虫氟苯双酰胺还未在水稻上登记。因此,氯虫苯甲酰胺、溴氰虫酰胺和四唑虫酰胺是目前防治水稻螟虫包括二化螟在内的主打双酰胺类药剂。关注二化螟对这3 种双酰胺类药剂的抗性和交互抗性发生程度,可保证其科学合理使用并持续发挥药效。高通量测序结合新型基因功能验证技术可为揭示二化螟多重抗性的机制提供分子手段。
杀虫剂抗性是一个遗传学现象,涉及到代谢和靶标等抗性相关基因及其调控网络。目前通过控制遗传调控治理害虫抗药性的技术日趋成熟,利用害虫的抗药性基因序列设计双链R N A互补识别靶基因mRNA,沉默抗性基因,可降低害虫抗药性水平[34];利用转基因技术表达害虫专一性抗药性基因dsRNA 的转基因植物[35];制备基于dsRNA 的纳米农药制剂对农作物叶面进行喷施或灌根处理[36],可达到杀虫剂减施增效的目的。基于靶标抗性基因的结构信息,设计和合成高活性、高亲和性和高选择性的化合物,是现代杀虫剂研发的重要途径[37]。因此,现阶段需要大力开发与推广遗传防治的技术,控制田间害虫种群数量在经济阀值以下,减少对化学农药的依赖。然而化学农药作为压制害虫种群的暴发或大规模迁移的应急手段,仍将在害虫防治中发挥重要作用,因此,通过监测害虫对杀虫剂抗性、探究其抗药性分子机制以及预测其对杀虫剂的抗性三管齐下的方法,可延缓害虫抗药性的产生和发展[1],也可为遗传防治提供新的候选基因。化学防治与遗传防治有机结合[29],将是治理二化螟抗药性和保持双酰胺类杀虫剂药效及延长其使用寿命的重要方向。
猜你喜欢
江苏农业科学(2022年22期)2022-12-13
特种经济动植物(2022年11期)2022-11-15
湖北植保(2020年5期)2020-10-20
中国果菜(2020年1期)2020-02-23
农药科学与管理(2019年10期)2019-04-20
吉林农业(2017年15期)2017-08-29
现代农业(2016年4期)2016-02-28
作物研究(2015年2期)2015-03-24
应用化工(2014年1期)2014-08-16
郑州大学学报(理学版)(2014年4期)2014-03-01
