
小鼠RNA结合蛋白QKI-5的生物信息学和组织表达分析及其过表达对癌症相关基因表达的影响
2023-05-08 01:32王逢博高登科张海森杨王浩靳亚平陈华涛
江西农业大学学报 2023年2期
王逢博,高登科*,李 超,李 丹,刘 薇,张海森,杨王浩,靳亚平,陈华涛*
(1.西北农林科技大学 动物医学院,陕西 杨凌 712100;2.农业农村部动物生物技术重点实验室,陕西 杨凌 712100)
【研究意义】RNA 结合蛋白(RNA-binding Proteins,RBPs)是一类存在于细胞中与mRNA 有高度亲和性的蛋白质,包含2 000种以上的蛋白质,是RNA 转录后调控过程的关键参与者。其RNA 结合域结构的灵活性以及功能的多样性,使RBPs 能够调控大量转录本的代谢过程[1-2]。Quaking(QKI)是一类重要的RBP,通过QKI反应元件(quaking response element,QRE)与RNA特异性结合,可以调控细胞分化[3-4]、细胞凋亡[5]、细胞周期、胶质细胞命运和发育[6-7]等细胞生物过程。研究发现QKI 能够抑制猪睾丸支持细胞凋亡,且可以促进未成熟的猪睾丸支持细胞中胰岛素样生长因子IGF-1 等分泌因子的分泌以及细胞增殖[8-9]。此外,QKI基因在山羊骨骼肌组织中高表达,且在山羊骨骼肌卫星细胞增殖分化的过程中表现出持续上升的表达规律,提示QKI 在山羊的骨骼肌发育的过程中可能也发挥一定作用[10]。近年来,许多研究证据表明Qki-5 转录本与前列腺癌[11]、肺癌[12-13]、卵巢癌[14]以及乳腺癌[15]等多种癌症的发生密切相关。但是,对于QKI-5的其他功能以及在肝癌发生发展过程中的影响作用鲜有报道。因此,预测QKI-5未知功能并研究其在肝癌发生发展过程中的作用具有重要意义。【前人研究进展】RBPs可通过直接或间接与mRNA 上的作用元件结合,在RNA 的转录后调控中起着关键作用[16-17],参与调控RNA 加工过程的各个方面,在调控RNA 编辑、代谢、剪接、定位和翻译等过程中发挥着重要作用[18-20]。研究证据表明,RBPs 表达改变和功能异常会影响RNA 表达的多个生理过程,进而引起免疫性疾病[21]、神经系统疾病[22]、心脏疾病[23-24]、急性白血病[25]以及结直肠癌[26-27]等多种疾病的发生。RNA 结合蛋白QKI 是STAR(signal transduction and activation of RNA)蛋白家族的一员,Qki基因位于人类第6 号染色体和小鼠第17 号染色体上,在KH 结构域内含有一个RNA 结合基序,其两侧有两个QUA 结构域(QUA1 和QUA2)[28]。QKI 选择性剪切生成5,6,7 kb大小的转录本,分别编码QKI-5、QKI-6、QKI-7蛋白。其中,QKI-5在基因序列上有一段非经典的核定位信号肽,因此其主要位于细胞核中,但在胞浆中也能检测到QKI-5,其可以发挥将胞浆信号输送到胞核的功能[29],而QKI-6、QKI-7主要存在于细胞质内[30]。近年来有许多研究证据表明QKI-5作为一个重要的转录因子,在许多生理、病理过程中发挥作用。Cochrane 等[3]研究发现,在小鼠诱导多能干细胞分化的内皮细胞中,过表达QKI-5能够显著改善实验性后肢缺血后血管生成、新生血管形成及血流恢复。Zhao 等[11]研究发现,Qki-5 基因在体外和体内均能有效抑制前列腺癌细胞增殖。赵易等[15]研究发现,在乳腺癌中QKI-5的高表达可能通过抑制癌细胞的细胞周期致使细胞增殖变缓,从而具有抑制肿瘤生长的作用。段小霞等[14]研究发现,QKI-5在卵巢癌细胞中特异性扩增后能够抑制卵巢癌细胞的增殖以及细胞克隆形成并促进癌细胞发生凋亡。【本研究切入点】QKI-5作为转录因子,其调控的下游靶基因众多,调控方式多种多样。但人们对QKI-5 参与调控哺乳动物多种生理、病理功能认识仍不全面,亟待进一步的深入探究。并且该基因在肝癌发生发展过程的影响仍尚不清楚。【拟解决的关键问题】本研究拟通过克隆小鼠Qki-5基因的蛋白编码序列(coding sequence,CDS),构建其真核表达载体,并对小鼠QKI-5进行了生物信息学分析,为进一步探究Qki-5基因的生物学功能提供关键材料支撑,为后续探究牛、羊等反刍动物Qki-5基因的功能提供前期研究基础;此外,本研究探索Qki-5 基因在不同组织的表达丰度及其在小鼠肝脏中的表达分布,分析在小鼠肝细胞AML12 中过表达Qki-5 基因后检测其对癌症相关基因表达的影响,以期为探究Qki-5 基因在小鼠AML12 细胞中影响癌症相关基因表达的作用机制提供关键材料与前期基础。
1 材料与方法
1.1 试验材料
试验于2022 年02 月至2022 年08 月在西北农林科技大学农业农村部动物生物技术重点实验室完成。试验所用的组织包括小鼠肝脏、心脏、脾脏、肾脏、肺脏、十二指肠、结肠、全脑、肌肉、附睾脂肪、皮下脂肪以及睾丸,共计12个组织,每种组织分别取自3只4月龄C57BL/6野生型雄鼠,小鼠购自北京维通利华公司;HEK293T、AML12细胞购自中国科学院细胞库。
1.2 主要试剂
DH5α感受态细胞、胶回收试剂盒、无内毒素质粒小提中量试剂盒购自北京天根生物科技有限公司;胎牛血清、Opti-MEM 培养基购自美国Gibco 公司,DMEM 高糖培养基购自Hyclone 公司,TurboFect Transfection Reagent 购自美国Thermo Fisher 公司;TRIzol、PrimerSTAR®Max DNA polymera 和Quick Cut™BmaHⅠ购自日本TaKaRa 公司;反转录试剂盒购自日本TOYOBO 公司、SYBR qPCR Master Mix 购自南京诺唯赞生物科技股份有限公司;RIPA 裂解液、PMSF、蛋白酶抑制剂、磷酸酶抑制剂均购自上海碧云天生物技术有限公司;BCA 蛋白浓度检测试剂盒购自江苏凯基生物技术股份有限公司;脱脂奶粉购自BD 公司;兔抗HA 抗体购自Abcam 公司;兔抗GAPDH 多克隆抗体购自武汉三鹰生物技术有限公司;HRP 共轭山羊抗兔抗体购自中国中杉金桥公司;ECL HRP 底物试剂盒购自北京迪宁生物科技有限公司;多聚甲醛固定液购自武汉赛维尔生物科技有限公司;兔抗QKI 单克隆抗体购自博士德生物工程有限公司;非特异性兔抗IgG 抗体购自Abcam 公司;免疫组化超敏UltraSensitiveTMSP 试剂盒购自福州迈新生物技术开发有限公司。
1.3 试验方法
1.3.1 引物设计与合成 根据GenBank 数据库中的小鼠Qki-5 基因CDS 区序列(登录号:NM_001159517.1),使用Primer Premier 6.0 软件设计PCR 扩增引物,引物序列如下:F:5′-AGAGAATTCGATATCGGATCCATGGTCGGGGAAATGGAAAC-3′;R:5′-GGTCTCGAGGGTACCGGATCCTTAGTTGCCGGTGGCGG-3′(下划线为BamHⅠ酶切位点),在上下游引物5′分别添加了pcDNA3.1-Puro-N-3HA载体BamHⅠ酶切位点两侧互补配对的同源序列,预计扩增产物长度为1 026 bp。引物交由西安擎科生物科技有限公司合成。
1.3.2 PCR扩增 称取20 mg小鼠肝脏组织,加入1 mL TRIzol,使用TRIzol法提取总RNA,并使用逆转录试剂盒将提取的RNA 逆转录为cDNA,反应液均在冰上配制,反应过程如下:RNA 5 µL 65 ℃变性5 min;反应后加入4×DN Master Mix 2 µL、Nuclease-free water 1 µL混匀,37 ℃温育5 min;之后加入5×RT Master Mix Ⅱ 2 µL,37 ℃ 15 min、50 ℃ 5 min、98 ℃ 5 min,反应结束后置于-20 ℃保存。以cDNA为模板,PCR扩增带有同源序列的Qki-5基因CDS区片段。PCR 反应体系50 µL:2×PrimerSTAR Max Premix 25 µL、上下游引物(10 µmol/L)各1 µL、cDNA 4 µL(50 ng/µL),加ddH2O 至50 µL。PCR 反应程序:98 ℃预变性3 min,98 ℃变性10 s,53.5 ℃退火15 s,72 ℃延伸75 s,循环35次,72 ℃终延伸5 min。对PCR产物进行琼脂糖凝胶电泳鉴定,符合预期的条带进行胶回收。
1.3.3 pcDNA3.1-mQki-5真核表达载体的构建 使用限制性内切酶BamHⅠ对pcDNA3.1-Puro-N-3HA载体进行线性化,反应体系:pcDNA3.1 2 µL,Quick CutTMBamHⅠ 1.5 µL,10×Quick Cut Green Buffer 2 µL,加ddH2O 至20 µL。将酶切产物进行琼脂糖凝胶电泳鉴定,并将目的条带进行胶回收纯化。之后将胶回收纯化的Qki-5基因CDS区片段与pcDNA3.1-Puro-N-3HA线性化载体进行重组连接,反应体系:线性化载体104.22 ng,插入片段41.04 ng,5×CE Ⅱ Buffer 4 µL,Exnase Ⅱ 2 µL,加ddH2O 至20 µL,37 ℃反应30 min。反应结束后,按照说明书将重组产物转化到DH5α 感受态细胞中,在含有羧苄青霉素抗性的LB 固体培养基中倒置培养37 ℃ 16 h,之后挑取单克隆菌落,置于37 ℃摇床扩增培养12 h,使用无内毒素质粒小提试剂盒提取质粒并测浓度。对重组质粒进行单酶切和PCR 鉴定,将符合预期的重组质粒送至西安擎科生物科技有限公司测序,将测序结果与NCBI 上的序列进行比对,并将测序正确的重组质粒重命名为pcDNA3.1-mQki-5。
1.3.4 生物信息学分析 利用DNASTAR 及MEGA11 软件对小鼠的Qki-5 基因和大鼠、人、山羊、绵羊、牛、牦牛、猪、马、双峰驼、斑马鱼、鲤鱼的Qki基因CDS 区序列进行同源性比对,并构建系统进化树;应用ProtParam(https://web.expasy.org/protparam/)在线软件对小鼠QKI-5蛋白理化性质进行分析,包括蛋白分子式及分子质量、原子总数、氨基酸数、理论等电点、稳定系数、脂肪族系数等理化性质;应用ProtScale(https://web.expasy.org/protscale/)在线软件对小鼠QKI-5 蛋白氨基酸序列的亲/疏水性进行预测;使用NetPhos 3.1 server(https://services.healthtech.dtu.dk/services/NetPhos-3.1/)在线软件对小鼠QKI-5 蛋白进行磷酸化位点预测;使用SingaIP5.0(https://services.healthtech.dtu.dk/services/SignalP-5.0/)在线软件预测小鼠QKI-5 蛋白信号肽;使用TMHMM2.0 Server(https://services.healthtech.dtu.dk/services/TMHMM-2.0/)在线程序进行跨膜区预测;使用PSORTII Prediction(https://psort.hgc.jp/form2.html)在线软件进行亚细胞定位预测;应用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)在线工具网站预测小鼠QKI-5 蛋白的二级结构;应用在线软件SWISS-MODEL(https://swissmodel.expasy.org/interactive)预测山羊、小鼠和人QKI-5 蛋白的三级结构,并使用PyMOL 对预测模型进行比较;最后,使用STRING(https://cn.string-db.org/)在线数据库预测小鼠QKI-5蛋白的互作蛋白。
1.3.5 实时荧光定量PCR(qRT-PCR)检测Qki-5基因在小鼠不同组织的表达谱 在同一时间点收取同一环境中3 只C57BL/6 雄鼠的肝脏、心脏、脾脏、肾脏、肺脏、十二指肠、结肠、全脑、肌肉组织、附睾脂肪组织、皮下脂肪组织以及睾丸组织,共12 个组织样品,分别称取各组织样品20 mg,加入1 mL TRIzol,提取组织总RNA,并反转录为cDNA,稀释至2.5 ng/µL 备用。使用Primer Premier 6.0 软件设计小鼠的Qki-5 基因和36b4基因的引物序列(表1),使用CFX96 荧光定量仪器反应,检测Qki-5 基因和36b4基因的表达。并以36b4基因为内参,使用2-ΔΔCt相对定量法计算不同组织Qki-5 基因的相对表达倍数[31]。
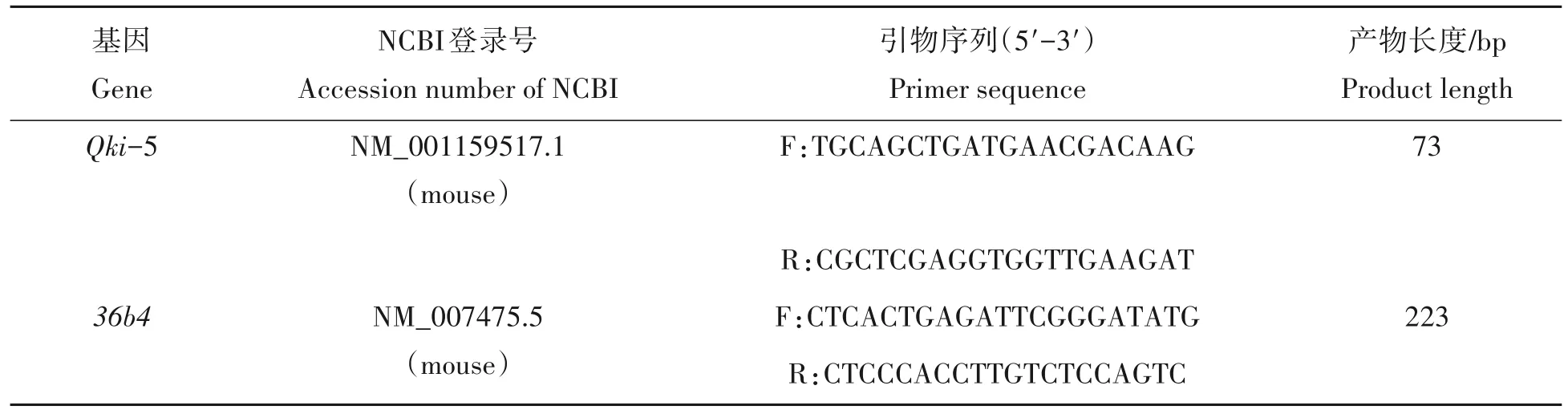
表1 引物序列信息Tab.1 Primer sequence information
1.3.6 免疫组织化学技术(IHC)检测QKI-5 在小鼠肝脏组织中的分布 使用多聚甲醛固定小鼠的肝脏组织,并将已固定的组织制成石蜡切片。使用二甲苯以及梯度酒精将石蜡切片脱蜡脱水;之后使用抗原修复液高火煮沸后调至中火维持20 min 断续沸腾状态,冷却至室温后,PBS 清洗3 次,每次5 min;0.2% TrionX-100室温透膜30 min,PBS 清洗3次,每次5 min;使用1% BSA 室温封闭1 h,之后甩干封闭液;QKI抗体(1:150 稀释)或非特异性IgG 抗体(1:200 稀释)在湿盒中4℃孵育过夜,分别为试验组与阴性对照组,次日室温孵育30 min 后,PBS 清洗5 次,每次5 min;使用免疫组化超敏UltraSensitiveTMSP 试剂盒进行免疫组织化学染色,之后使用光学显微镜观察QKI-5的分布并保存图像[32]。
1.3.7 重组质粒转染AML12 细胞 复苏AML12 细胞,培养24 h 后,将生长状态良好的细胞分别传代至两个6 孔板中,待细胞密度达到60%左右时进行转染。将pcDNA3.1-Puro-N-3HA 空载体和pcDNA3.1-mQki-5 重组质粒按照Turbofect 转染试剂说明书分别转染至AML12 细胞。每个6 孔板中,对照组和试验组各转染3孔,转染后培养24 h收取RNA样品,48 h后收取蛋白样品,分别用于检测小鼠QKI-5在mRNA水平和蛋白水平的表达。
1.3.8 qRT-PCR 检测Qki-5 基因在AML12 细胞的表达 转染24 h 后,加入TRIzol 提取试验组及对照组细胞总RNA,逆转录为cDNA,稀释至2.5 ng/µL 备用。使用qPCR 检测Qki-5 基因和36B4基因在mRNA水平的表达(表1)。反应体系如下:2×ChamQ SYBR qPCR Master Mix 10 µL,上、下游引物各0.4 µL、cDNA 5 µL(2.5 ng/µL),加ddH2O 补足至20 µL。反应程序:95 ℃ 30 s;95 ℃ 10 s,60 ℃ 30 s,共40 个循环;95 ℃ 15 s;60 ℃ 60 s;95 ℃ 15 s。使用荧光定量系统进行反应,并以36b4基因为内参,使用2-ΔΔCt相对定量法计算试验组与对照组Qki-5基因相对表达量及相对倍数。
1.3.9 Western Blotting(WB)检测QKI-5在AML12细胞中的表达 转染48 h后收取细胞样品,按说明书使用RIPA 裂解液提取试验组与对照组的细胞总蛋白,之后使用BCA 试剂盒检测蛋白浓度,并加入裂解液将样品稀释至统一浓度,之后加入上样缓冲液,100 ℃ 水浴10 min,样品放-20 ℃保存或直接进行SDSPAGE 凝胶电泳。电泳结束后,将蛋白转印至PVDF 膜上,之后使用10%脱脂奶粉封闭2 h。封闭完后,使用TBST 洗膜3 次,每次10 min;之后将PVDF 膜置于兔抗HA 标签的一抗工作液(1∶4 000),4 ℃孵育过夜;TBST 洗膜3 次,每次10 min;之后将PVDF 膜置于山羊抗兔二抗工作液(1∶5 000),室温孵育2 h,用TBST洗膜3次,每次10 min。之后将膜在ECL发光液中浸泡2 min,于暗室曝光显影并保存蛋白结果[33]。
1.3.10 在AML12细胞中检测癌症相关基因的表达 分别在mRNA 和蛋白水平验证QKI-5在AML12细胞成功过表达后,使用Primer Premier 6.0 软件设计引物序列(表2),使用荧光定量系统检测Bcl2、P53、Ccnd1、CerbB2、Rasa1、Kras、Cdkn1a、Cdkn1b等8个癌症相关基因的mRNA表达量,并以36b4基因为内参,使用2-ΔΔCt相对定量法计算不同基因的相对表达倍数。

表2 癌症相关基因引物序列信息Tab.2 Primer sequence information for cancer-related gene
1.3.11 数据统计分析 采用2-ΔΔCt法将qPCR中的Ct值转化为基因相对表达量,利用GraphPad Prism 6进行数据分析,并进行非配对t检验,P<0.05表示差异显著,P<0.01表示差异极显著。
2 结果与分析
2.1 小鼠Qki-5基因克隆及真核表达载体的构建
以小鼠肝脏cDNA 为模板,使用带有同源臂的引物进行PCR 扩增,将扩增产物进行琼脂糖凝胶电泳。电泳结果如图1A 所示,扩增片段符合预期目的片段大小(1 026 bp)。将目的条带进行胶回收,与使用BamHⅠ限制性内切酶线性化的载体进行同源重组连接,构建pcDNA3.1-mQki-5 真核表达载体。对重组载体与空载体进行单酶切鉴定,结果如图1B所示,空质粒大小为5 211 bp,目的片段大小为1 026 bp,酶切结果符合预期。并将重组质粒pcDNA3.1-mQki-5进行测序分析,测序结果与NCBI数据库中已公布序列完全一致。以上结果表明,pcDNA3.1-mQki-5真核表达载体构建成功。
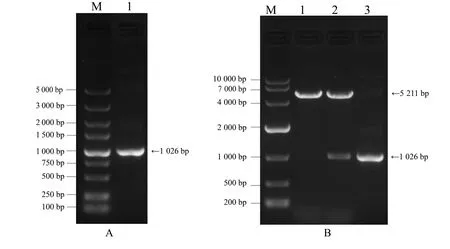
图1 小鼠Qki-5基因CDS扩增与pcDNA3.1-mQki-5真核表达载体鉴定结果Fig.1 Results of mouse Qki-5 gene CDS amplification and identification of pcDNA3.1-mQki-5 eukaryotic expression vector
2.2 生物信息学分析
2.2.1 不同物种间Qki基因CDS 序列相似性比对及进化树构建 利用DNASTAR 及MEGA11 对12 个物种的Qki基因CDS区序列进行同源性比对,并构建系统进化树。相似性比对结果如图2所示,小鼠Qki基因的CDS 区与人、大鼠、山羊、绵羊、牛、牦牛、猪、马、双峰驼、斑马鱼、鲤鱼的相似性分别是96.4%、95.5%、94.5%、94.6%、94.2%、92.7%、95.6%、95.7%、89.8%、68.5%、67.8%,由此可知,小鼠Qki基因的CDS区与人的相似性最高,与鲤鱼的相似性最低。系统进化树的结果如图3所示,这12个物种之间,小鼠Qki基因与大鼠的遗传距离最近,其次是人、马和猪等其他哺乳动物,与鲤鱼的遗传距离最远。
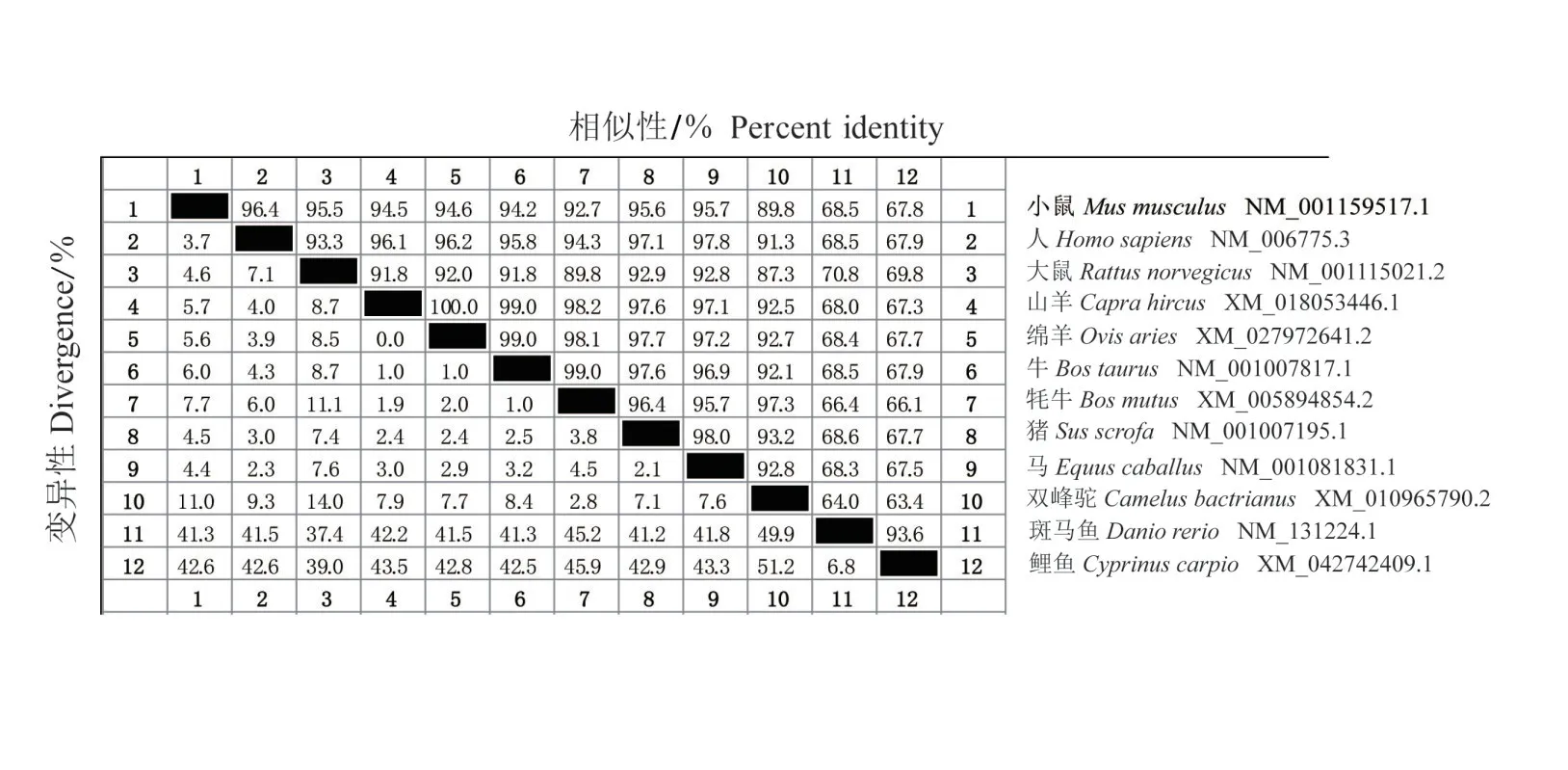
图2 Qki基因CDS区序列相似性比对Fig.2 Homology comparison of Qki gene’s CDS region

图3 小鼠Qki基因的系统进化树Fig.3 Phylogenetic tree of Qki gene in mice
2.2.2 蛋白理化性质预测 应用ProtParam 在线软件对小鼠QKI-5 蛋白理化性质进行分析,结果显示小鼠QKI-5蛋白分子式为C1672H2713N463O493S15,原子总数为5 356,分子质量约为37.67 ku。QKI-5蛋白的氨基酸组成如表3 所示,共包含氨基酸341 个,其中色氨酸(Trp)含量最低,丙氨酸(Ala)、亮氨酸(Leu)和脯氨酸(Pro)含量较高,不含有吡咯赖氨酸(Pyl)和硒代半胱氨酸(Sec)。理论等电点为8.63,为碱性蛋白质;不稳定系数为35.08,为稳定蛋白质;总平均亲水性(GRAVY)为-0.388;脂肪系数为87.01。
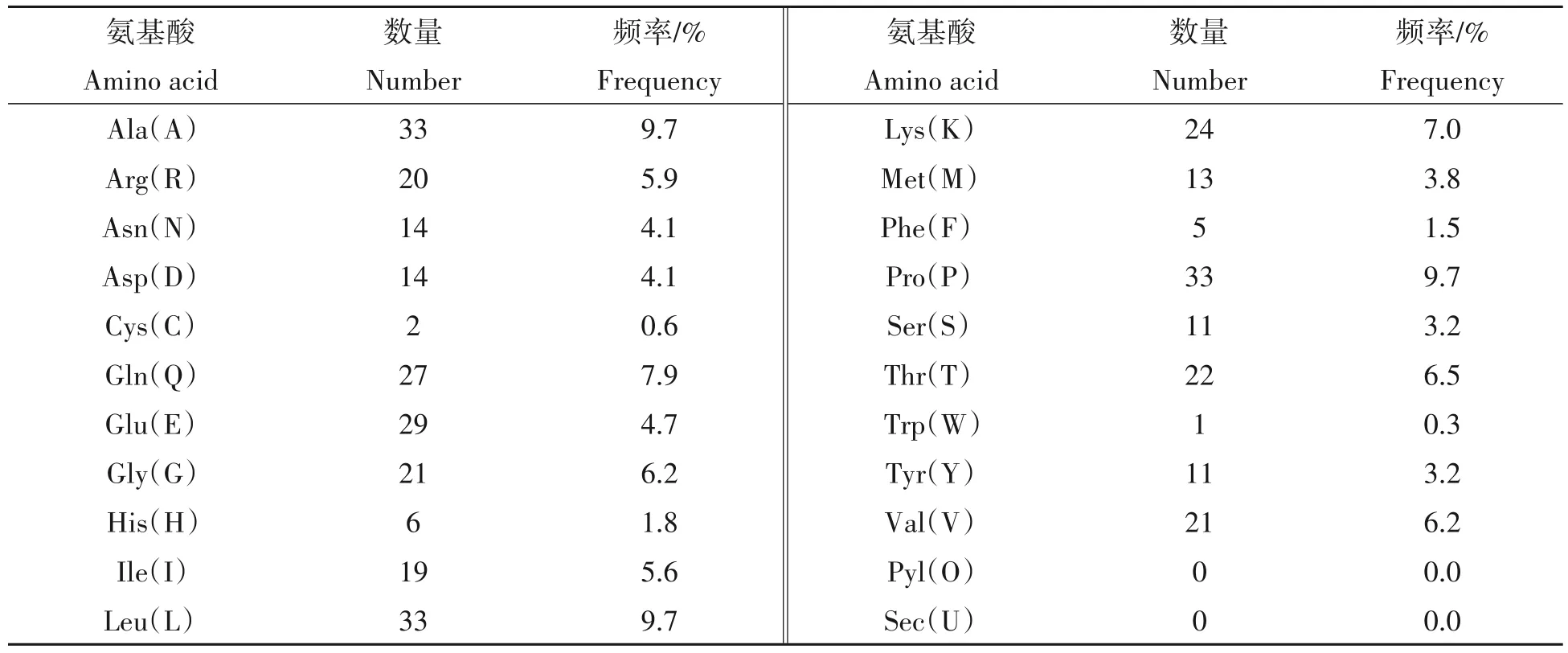
表3 小鼠QKI-5蛋白氨基酸组成Tab.3 Amino acid composition of mouse QKI-5 protein
2.2.3 亲/疏水性预测 应用ProtScale 在线软件对小鼠QKI-5蛋白氨基酸序列的亲/疏水性进行预测,结果显示,最低分值(-3.744)位于第134、135位氨基酸,表明亲水性最强;最高分值(1.789)位于第217、218位氨基酸,表明疏水性最强;且该蛋白大部分位于亲水区域,故小鼠QKI-5蛋白为亲水性蛋白。
2.2.4 磷酸化位点预测 使用NetPhos 3.1 server在线软件对小鼠QKI-5蛋白进行磷酸化位点预测,预测结果表明,QKI-5 蛋白含9 个丝氨酸(Serine)磷酸化位点,12 个苏氨酸(Threonine)磷酸化位点,5 个酪氨酸(Tyrosine)磷酸化位点,共26个磷酸化位点。
2.2.5 信号肽及跨膜结构预测 使用SingaIP5.0 在线软件预测小鼠QKI-5 蛋白信号肽。结果显示,该蛋白不存在信号肽;使用TMHMM2.0 Server 程序进行跨膜区预测,结果表明,QKI-5 蛋白不存在跨膜区域;使用PSORTII Prediction 在线软件进行亚细胞定位预测,结果显示,该蛋白有56.5%定位于细胞核上,17.4%定位于细胞质中,8.7%定位于高尔基体,4.3%定位于分泌小泡、过氧化物酶体、线粒体和细胞骨架。
2.2.6 蛋白质二级结构预测 应用SOPMA 在线工具网站预测小鼠QKI-5蛋白的二级结构,预测结果表示:小鼠QKI-5 主要由α-螺旋、β 转角、延伸链、无规则卷曲构成,其中α-螺旋占32.84%,β 转角占4.40%,延伸链占14.96%,无规则卷曲占47.80%,推测无规则卷曲是QKI-5最大的二级结构原件。
2.2.7 蛋白质三级结构预测 应用在线软件SWISS-MODEL 预测小鼠的QKI-5 蛋白(NCBI 登录号:NP_001152989.1),以及人(NCBI 登录号:NP_006766.1)和大鼠QKI 蛋白(NCBI 登录号:NP_001108493.1)的三级结构,结果如图4 所示。使用PyMOL 对预测模型进行比较,结果显示,三者QKI 蛋白模型间原子位置均方根偏差(Root-Mean-Square Deviation,RMSD)均为0.00,说明小鼠预测QKI蛋白的三级结构与人和大鼠之间的三级结构没有差异;并且将三者的氨基酸序列在NCBI 在线网站上进行比对,结果显示小鼠与人的氨基酸序列完全相同,小鼠与大鼠的氨基酸序列相似率为91%。3 个物种之间的三级结构一致,说明其在功能上可能也具有高度相似性。
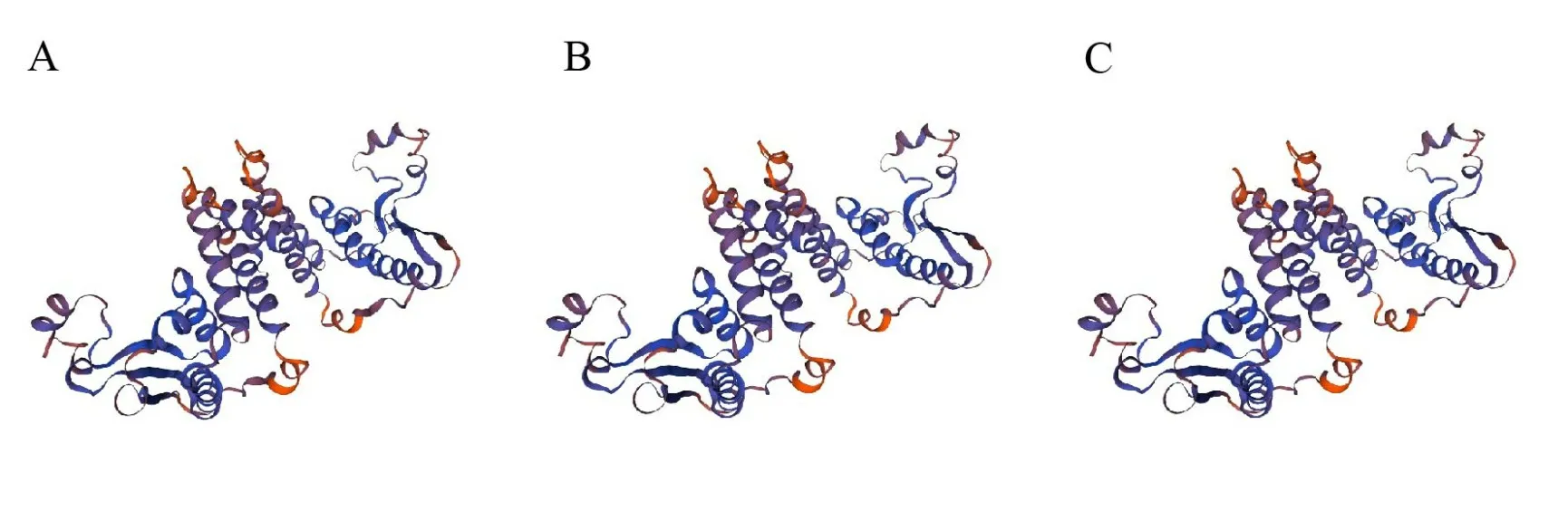
图4 小鼠、人、大鼠QKI蛋白三级结构预测Fig.4 Prediction of the tertiary structure of mouse QKI protein in mouse,human and rat
2.2.8 QKI-5蛋白的互作蛋白预测 使用STRING 在线数据库预测小鼠QKI-5蛋白的互作蛋白,预测结果如图5所示,QKI-5蛋白主要与多聚胞嘧啶结合蛋白4(Poly rC binding proteins,PCBP4)、多重剪接RNA结合蛋白2(RNA binding protein with multiple splicing 2,RBPMS2)、真核翻译起始因子4E 核输入因子1(Eukaryotic translation initiation factor 4E nuclear import factor 1,Eif4enif1)、RNA 结合fox-1 同源物2(RNA binding fox-1 homolog 2,RBFOX2)、RNA 结合蛋白mRNA 加工因子(RNA binding protein mRNA processing factor,RBPMS)、远上游元件结合蛋白(Far upstream element binding protein 3,FUBP3)、U2 小核RNA辅助因子2(U2 small nuclear RNA auxiliary factor 2,U2AF2)、多聚胞嘧啶结合蛋白29(Poly rC binding protein 2,PCBP2)、多聚嘧啶束结合蛋白2(Polypyrimidine tract binding protein 2,PTBP2)、核异质核糖核蛋白L样(Heterogeneous nuclear ribonucleoprotein L like,hnRNPLL)等蛋白之间存在互作。
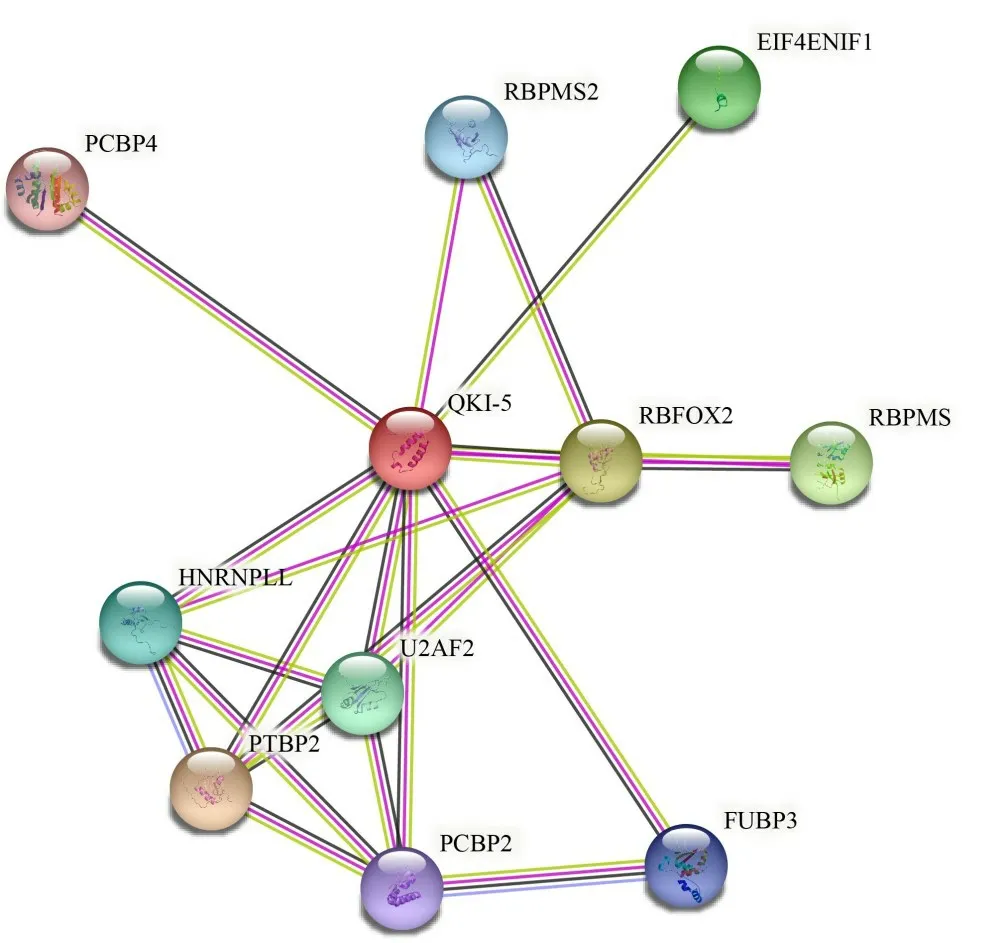
图5 小鼠QKI-5蛋白互作蛋白预测Fig.5 Prediction of proteins interaction with mouse QKI-5 protein
2.3 Qki-5基因在小鼠不同组织中的表达谱
提取3 只C57BL/6 雄性小鼠的12 个组织总RNA,反转录为cDNA 后,检测小鼠Qki-5 基因的表达,以36b4基因为内参,计算Qki-5 基因的相对表达量,并均一化各组织表达量,分析12 个组织的表达相对变化,结果如图6所示。在12个组织中,全脑的表达量最高,其次是肺脏、皮下脂肪组织以及附睾脂肪组织等,肝脏的表达量处于中等水平,十二指肠和结肠的表达量最低。
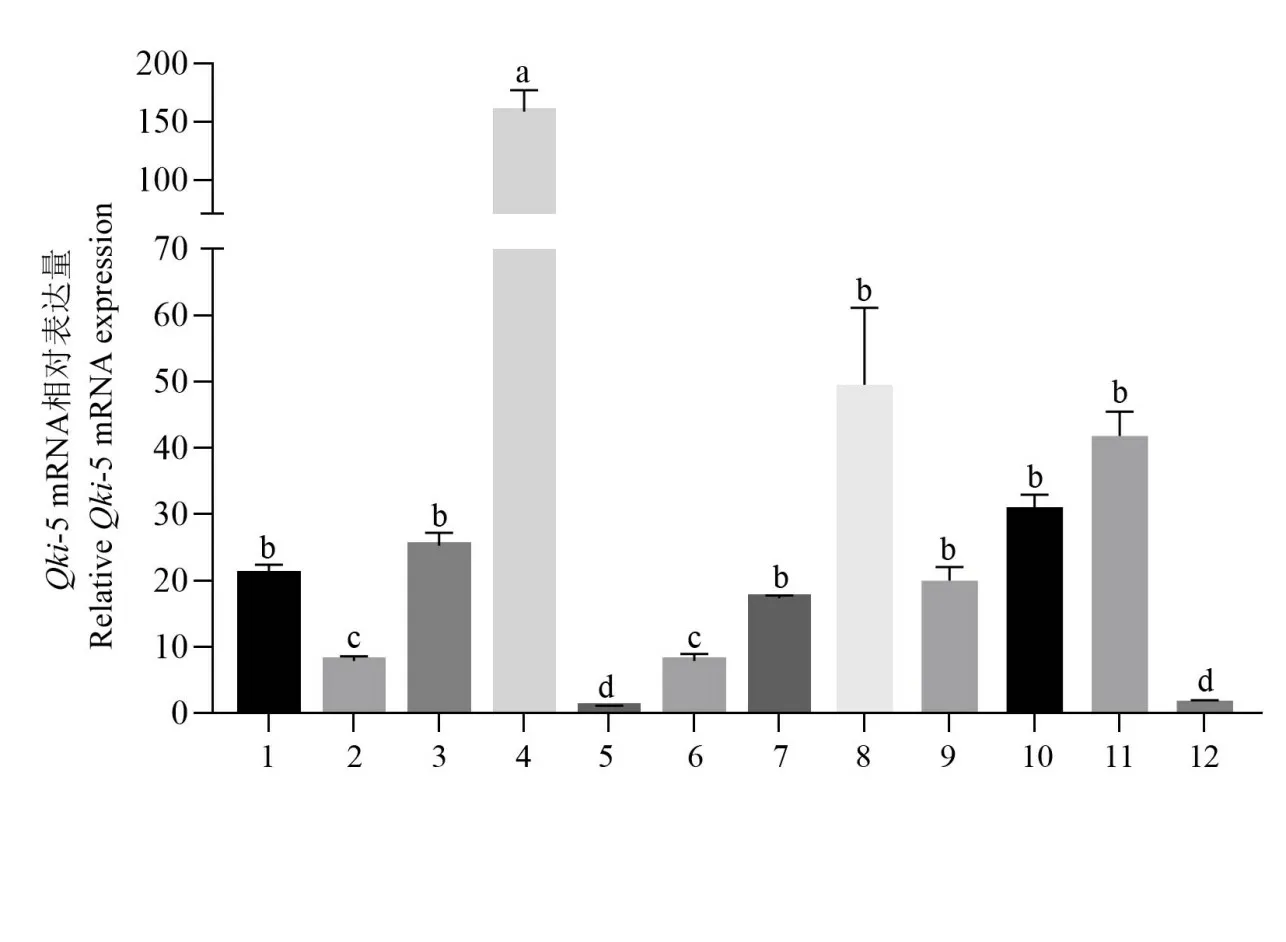
图6 小鼠Qki-5基因mRNA在不同组织的表达丰度Fig.6 Expression abundance of mouse Qki-5 gene’s mRNA in different tissues
2.4 QKI-5蛋白在小鼠肝脏组织中的表达分布
通过免疫组织化学技术检测小鼠肝脏中QKI-5 的表达分布,结果如图7 所示,QKI-5 表达分布于肝细胞中,且主要定位于细胞核内。
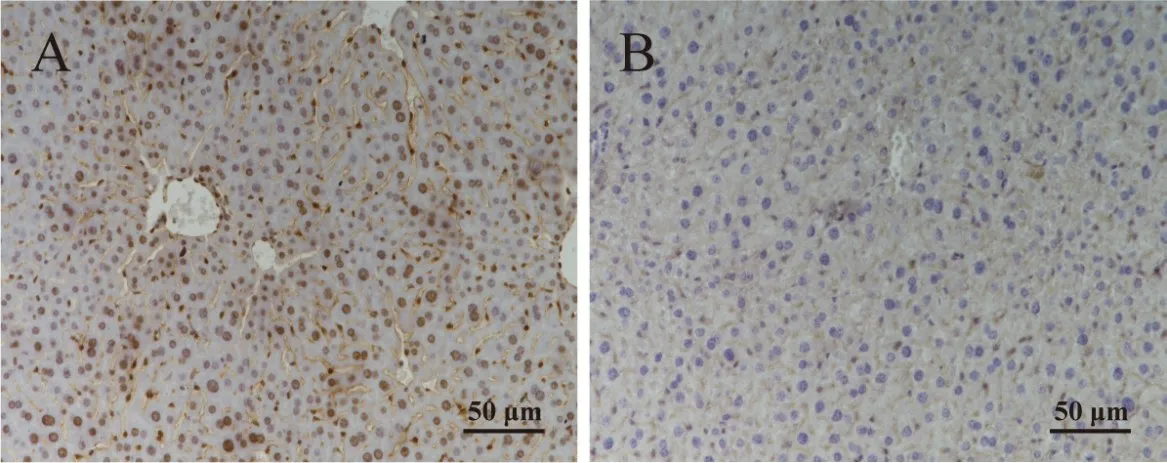
图7 QKI-5在小鼠肝脏组织中的表达分布Fig.7 The distribution of QKI-5 in mouse’s liver
2.5 QKI-5在AML12细胞中成功过表达
将pcDNA3.1-mQki-5 重组质粒和pcDNA3.1-Puro-N-3HA 空质粒分别转染至AML12 细胞后,检测QKI-5的表达水平。结果如图8A 所示,pcDNA3.1-mQki-5试验组Qki-5基因的mRNA 表达量升高,为对照组的5.78 倍(P<0.01)。表明,pcDNA3.1-mQki-5 重组质粒转染AML12 细胞中在mRNA 水平成功过表达。由图8B可知,试验组细胞样品中经HA抗体孵育后,在约38 ku处有明显的条带出现,而在对照组中无条带出现。该结果表明pcDNA3.1-mQki-5 重组质粒转染AML12 细胞中在蛋白水平成功过表达。以上结果表明pcDNA3.1-mQki-5重组质粒转染AML12细胞中在mRNA和蛋白水平均成功过表达。
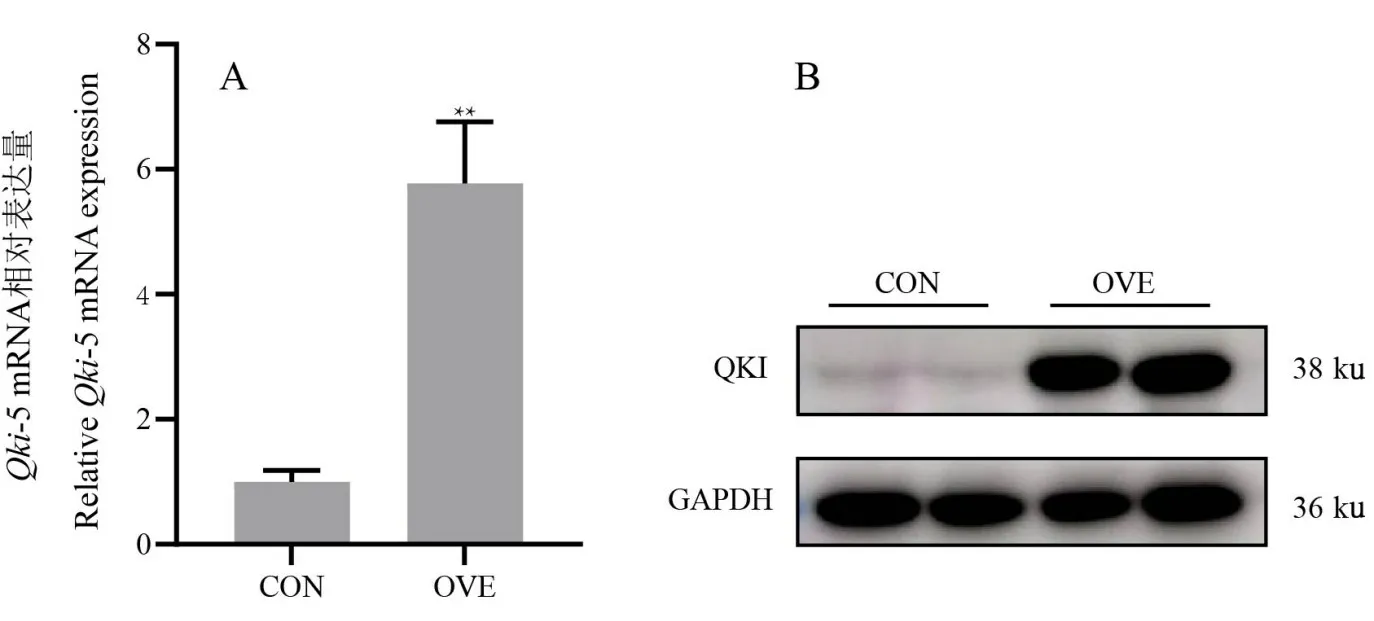
图8 qPCR及Western Blotting检测结果Fig.8 Results of qPCR and WB analysis
2.6 AML12细胞过表达Qki-5对癌症相关基因表达的影响
基于结果2.5,进一步检测对照组和试验组原癌基因Bcl2、Kras、Ccnd1、CerbB2以及抑癌基因P53、Rasa1、Cdkn1a、Cdkn1b8 个癌症相关基因的相对表达量,结果如图9 所示,与对照组相比,试验组的P53mRNA 水平显著增加至3.32 倍(P<0.01),试验组的Ccnd1mRNA 增加至1.69 倍(P<0.05),Bcl2、CerbB2、Rasa1、Cdkn1a、Cdkn1b、Kras基因的mRNA水平均未见变化。
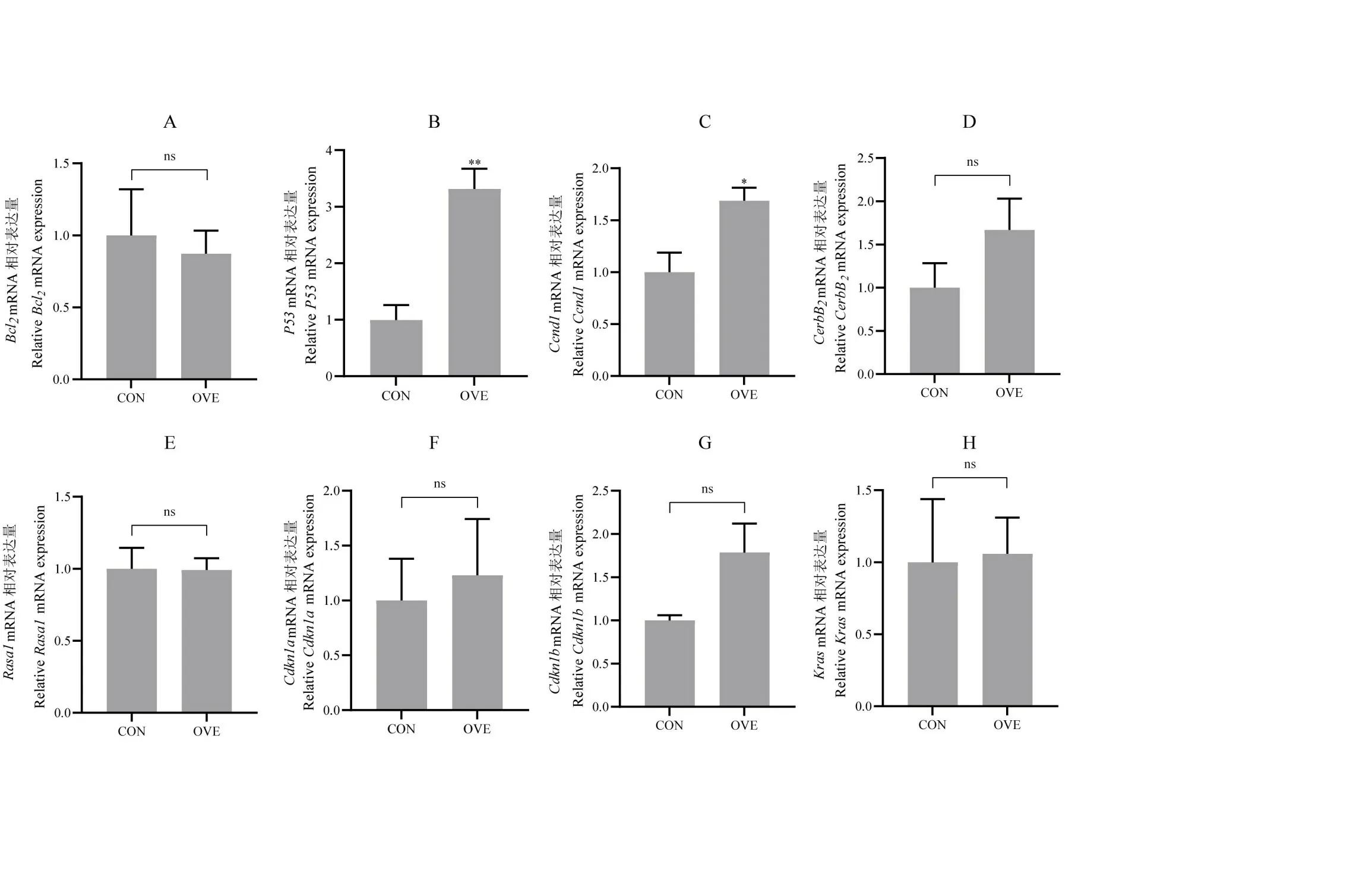
图9 癌症相关基因mRNA相对表达量Fig.9 Relative mRNA expression levels of cancer related genes
3 结论与讨论
近年来,随着科学技术的发展,人们对于RBPs 的研究不断深入,逐渐证明RBPs 在许多生理过程中都发挥重要作用。QKI蛋白是一类重要的RNA 结合蛋白,而QKI-5作为其重要转录本所编码的蛋白,在RNA 转录后调节的过程中也发挥着重要作用。QKI-5作为重要的转录因子,人们对其的生理、病理功能的认识尚不全面。目前,对于RNA 结合蛋白QKI-5 的研究主要集中在其对于乳腺癌、肺癌、卵巢癌等多种恶性肿瘤的影响作用[11-15,34],而其在肝癌发生发展过程中作用的研究未见报道。癌症在动物中的发生率和死亡率很高,严重威胁着动物的生命与健康。因此,本研究构建了小鼠Qki-5基因真核表达载体,通过生物信息学在线软件对QKI-5 蛋白的结构和功能特征进行预测分析;检测QKI-5 在不同组织的表达谱以及在小鼠肝脏中的表达分布;并对于在AML12 细胞中过表达QKI-5 后对癌症相关基因的影响进行研究,以期为后续研究QKI-5在不同生理、病理过程中的功能以及在小鼠AML12细胞中影响癌症相关基因表达的作用机制提供前期材料。
蛋白质互作预测结果表明,与QKI-5蛋白相互作用的前十位蛋白中,主要包括PCBP4、RBPMS2、RBPMS、RBFOX2 以及FUBP3。其中,RNA 结合蛋白PCBP4 是p53 的靶蛋白,在细胞周期中发挥作用,并参与肺肿瘤的抑制作用。Yan 等[35]研究结果表明PCBP4 缺乏的小鼠容易发生肺腺癌、淋巴瘤和肾肿瘤,并且PCBP4缺乏的小鼠胚胎成纤维细胞(MEFs)表现出细胞增殖增加但细胞衰老减少。Notarnicola 等[36]发现,RNA 结合蛋白RBPMS2具有调节雏鸡胃肠道平滑肌发育的作用,其持续表达可通过抑制骨形态发生蛋白的活性,来抑制平滑肌细胞分化标志物的表达,从而刺激平滑肌细胞的增殖。此外,Rabelo-Fernandez 等[37]的研究证据表明,RBPMS 可作为抑癌基因发挥作用,降低RBPMS 水平可以促进卵巢癌细胞增殖,降低细胞对顺铂敏感性,促进卵巢癌细胞对顺铂的耐药。研究证据表明,在糖尿病的发展过程中,RBFOX2 通过与靶基因AS结合,破坏正常的基因在心脏的表达模式,从而引起糖尿病早期心肌病的发生[38]。远上游元件结合蛋白FUBP3具有抵抗猪流行性腹泻感染的功能,研究证据表明,FUBP3可以降解猪流行性腹泻病毒核衣壳蛋白并诱导I型干扰素的产生[39]。上述证据表明,QKI-5可能参与调控癌症、心脏相关疾病以及参与免疫等众多生理、病理过程,但该功能仅为初步预测分析结果,仍需进一步验证。
此外,本研究通过qPCR检测小鼠Qki-5基因在不同组织表达分布,结果表明该基因在所检测的12个组织中均有表达且在全脑中表达量最高,其次是肺脏。由于全脑主要与神经系统相关功能有关,因此推测QKI-5可能在调控神经系统和肺脏的生命活动过程中具有一定作用。研究发现,Qki基因的突变会导致中枢和周围神经系统髓鞘形成障碍而引起频繁震颤,严重可于胚胎早期死亡,也因此得名为Quaking[40]。Hayakawa-Yano 等[41]研究发现QKI-5可以抑制神经元迁移并诱导异位有丝分裂细胞,在神经干细胞中发挥功能。Wang等[12]研究结果表明,QKI-5可以调控肺癌细胞骨架基因ADD3的可变剪接,反映了QKI-5的对肺癌的抑制作用;Zhou等[42]研究发现QKI-5可以通过抑制β-catenin信号通路,从而抑制肺癌细胞的侵袭性。以上结果表明Qki-5基因在神经系统调节以及肺脏相关疾病的发生发展中发挥一定作用。
有许多研究表明QKI-5 对于多种癌症的发生发展具有一定影响作用,因此笔者在小鼠肝细胞中过表达QKI-5 后检测对癌症相关基因的影响。由图9 结果可知,QKI-5 过表达后会引起P53和Ccnd1的相对表达量显著升高,而Bcl2、CerBb2、Rasa1、Cdkn1a、Cdkn1b、Kras未见明显变化。P53基因是重要的肿瘤抑制因子,可以保护细胞免受癌基因激活、辐射、有丝分裂应激、核糖体应激等一系列生理应激反应[43]。在AML12 细胞中过表达QKI-5 后引起P53基因表达水平升高,说明在正常细胞中QKI-5 的过表达可以激活P53的水平升高,从而可以产生一定的抑癌作用。细胞周期蛋白D1(cyclin D1,CCND1)由位于11号染色体第一区3号带(11q13)的Ccnd1基因编码,是一种调控细胞周期的蛋白,在细胞周期G1/S转变中发挥重要作用[44],能够造成细胞增殖甚至恶性增长,具有肿瘤因子的性质。有研究结果表明,在口腔鳞状细胞癌中,QKI-5的低表达可增加Ccnd1的表达,从而促进口腔鳞状细胞增殖。而在AML12细胞中过表达QKI-5 后,Ccnd1的mRNA 水平升高,说明在正常细胞中过表达QKI-5 不能抑制Ccnd1的表达,反而会造成Ccnd1的水平升高,通过该结果推测QKI-5 在正常细胞中不能通过抑制Ccnd1发挥抑癌作用,推测QKI-5 主要通过Ccnd1mRNA 抑制癌细胞的增殖而发挥抑癌作用,对正常细胞没有抑癌作用。通过P53和Ccnd1基因表达结果,可推测对于癌症疾病QKI-5可能在正常细胞中通过激活P53的表达而具有预防作用,而在正常细胞中QKI-5并不通过Ccnd1产生抑癌作用。
本研究成功构建了小鼠Qki-5基因真核表达载体,并通过生物信息学分析预测了其蛋白功能。获得了不同组织的表达谱,Qki-5基因在小鼠体内表达,且不同组织间表达量有明显差异,在全脑、肺脏中表达量较高。在小鼠肝脏中,QKI-5表达分布于肝细胞中,且主要定位于细胞核内。在AML12细胞中,QKI-5的过表达引起P53和Ccnd1的水平升高,提示QKI-5在正常细胞中可通过激活P53而发挥抑癌作用,在正常细胞中QKI-5并不通过Ccnd1产生抑癌作用。本研究结果提示QKI-5可能参与调控神经系统、心脏疾病等生理、病理过程,并证明QKI-5对于小鼠AML12细胞中癌症相关基因的表达具有一定影响。
致谢:国家卫生健康委员会时间生物学重点实验室2022 年度开放基金(NHCC-2022-01)同时对本研究给予了资助,谨致谢意!
猜你喜欢
黄河之声(2022年10期)2022-09-27
中学生数理化(高中版.高二数学)(2022年4期)2022-05-25
中学生数理化(高中版.高二数学)(2022年4期)2022-05-25
疯狂英语·新读写(2021年10期)2021-12-07
学苑创造·A版(2020年9期)2020-10-13
奥秘(2019年8期)2019-08-28
小学生学习指导(低年级)(2017年10期)2017-10-10
商周刊(2017年7期)2017-08-22
中学生数理化·八年级物理人教版(2017年11期)2017-04-18
小猕猴智力画刊(2016年6期)2016-05-14
