
MITF致病性突变与Waardenburg综合征临床表型相关性分析
2023-05-22 06:49倪晓琛李佳楠杨仕明陈伟
中华耳科学杂志 2023年2期
倪晓琛 李佳楠 杨仕明 陈伟
解放军总医院第六医学中心耳鼻咽喉头颈外科医学部(北京 100853)
国家耳鼻咽喉疾病临床医学研究中心(北京 100853)
聋病教育部重点实验室(北京 100853)
聋病防治北京市重点实验室(北京 100853)
MITF(microphthalmia-associated transcription factor)作为一种基础螺旋-环-螺旋-亮氨酸拉链结构(basic helix-loop-helix leucine zipper,bHLH-Zip)家族中的转录因子,其主要功能是调控基因的表达水平[1]。不仅可以参与调节神经棘细胞的分化以及发育的过程,同时可以调节黑色素的合成。MITF突变可能导致多种不同的临床综合征[2,3],其中以Waardenburg 综合征(Waardenburg Syndrome,WS)最为多见。
WS是一种以听力色素异常为特征的显性遗传的综合征性疾病,呈现高度外显性,临床特征具有明显异质性。其中以虹膜异色、前额白发、感音神经性聋、以及内眦异位为主要临床表型特征。WS患者可以出现不同程度的感音神经性聋,约占WS患者的90.2%[4]。其中以MITF为致病基因的WS 患者感音神经性聋的发生率为89.6%[4]。WS 还可以表现出不同程度不同部位的色素异常,包括虹膜颜色,头发,面部以及全身皮肤。
目前WS 临床表型以及突变位点的相关性的研究局限于PAX3以及SOX10[5]。研究表明无义介导的mRNA 降解途径(nonsense-mediated mRNA decay,NMD)对截短蛋白的调控可以部分解释临床表型的异质性。MITF突变导致WS 的原因同样为单倍体剂量不足[6,7]。而目前有关MITF的相关研究尚未有明确报道。MITF是否也受到NMD 机制的调控,从而影响疾病的表型,仍需更多的研究探索。本研究拟采用meta 分析的方法,检索目前已知的WS的MITF突变位点,并收集突变位点相关临床表型,通过广义估计方程(generalized estimating equations,GEE)对突变位点以及表型的相关性进行评估,探寻突变位点的突变规律,突变位点与临床表型之间的相关性。
1 方法
1.1 建立MITF突变数据库
1.1.1 文献纳入标准
由两名具有资质相同的人员进行文献检索。查找所有MITF为致病基因的WS 文献,文献纳入标准:(1)研究中纳入的人群为明确诊断为WS 的患者。WS的诊断标准如下:(2)已行基因检测明确致病基因为MITF突变;(3)具有明确MITF的突变位点。排除标准:(1)合并其他有关遗传性耳聋的致病基因;(2)报道中MITF并非新发致病性突变。
1.1.2 文献检索策略以及筛选过程
文献检索范围从1995年至2021年11月10日,于PubMed,CNKI数据库进行相关文献检索,使用检索词”Waardenburg syndrome”;“MITF”,“Waardenburg综合征”;共检索到247 篇文章,并由两名研究人员依据文献摘要进行初步筛选,将可能符合纳入标准的文献全文下载,阅读全文并依据纳入标准对文献进行筛选。在初步选定的36篇文献中,8篇文献不符合纳入标准。1 篇文献主要内容为MITF突变致病机制的相关研究,其余7篇文献中,有2篇文献中患者携带除MITF外的其他遗传性聋相关基因,5篇文献中报道的致病性突变位点并未新发突变。故最终纳入文献共计28篇[8-35]。文献检索流程图见图1。

图1 文献检索过程Fig.1 Flow Diagram
1.2 数据收集
收集纳入文献中患者的突变位点信息,纳入数据库中致病性突变位点共计34 例。并同时于ClinicalVariant数据库进行MITF致病性突变位点检索,检索到致病性突变位点共计53 例并纳入数据库。将纳入数据库中所有研究中的突变位点标准化,利用MutationTaster[36]对突变位点编号标准化,参考MITF-205(NM_000248.4)编码区碱基序列进行编号。突变位点相同时则进行合并。基于Ensembl 平台[37](https://www.ensembl.org/index.html)明确突变位点的功能区位置,以及突变基因编码蛋白质的氨基酸序列、可能影响的蛋白质结构域。收集整理研究中WS患者的临床表型以及听力学特征。
1.3 数据整理
突变类型:依据以下突变类型进行分类:错义突变、无义突变、移码突变、剪切突变、插入突变、缺失突变。
表型描述:依据研究中描述的临床表型进行描述以及分类。根据WS 表现出的主要临床特征,大致分为三类,虹膜颜色改变(Heterochromia irides;Normal),皮肤色素异常改变,头发颜色改变(Abnormality of the hair;Normal)。其中,进一步将皮肤色素异常分类,分为面部雀斑(Freckles),皮肤脱色素白斑(hypopigmentation),二者均有(Both),以及二者均无(None),共计四种类型。由于研究中多数患者均为重度感音神经性耳聋,未进一步描述听力学信息。
1.4 数据分析
若以突变位点进行分组,各组中患者数量较少,易产生偏倚,故不适宜针对突变位点进行分组,故而采用突变基因编码蛋白质结构以及功能的改变分组,分析不同组间与临床表型之间的相关性。依据MITF-M 蛋白的功能以及结构特点[38],将突变蛋白氨基酸改变,分为以下八种,DNA 结合阈的氨基酸替代(AA substitution in the DNA binding region,AA of DNA),HLH 结构阈的AA 替代(AA substitution in the HLH region,AA of HLH),核定位信号的AA 替代(AA substitution in the Nuclear location signal(NLS)region,AA of NLS),缺失DNA结合阈(deletion of DNA binding region,deletion of HLH region,deletion of DNA),缺失HLH 结构阈(deletion of HLH region,deletion of HLH),缺失核定位信号(deletion of the NLS region,deletion of NLS),缺失全部bHLH-Zip 结构阈(deletion of the subtotal region,deletion of subtotal),以及未缺失bHLH-Zip 结构阈(deletion of none of the above region,deletion of none)。家系中不同成员之间携带相同的致病基因突变位点,就不同致病位点而言,属于重复多次测量资料,但是由于家系中各成员之间具有相关性,不具备Logistic 回归的数据独立性条件,故采用广义估计方程(generalized estimating equations,GEE)进行数据分析。以SPSS17.0 软件进行GEE 数据分析。将不同突变蛋白中的氨基酸改变的类型作为自变量,临床表型作为因变量进行分析,比较分析不同的氨基酸改变与表型之间的关系。首先进行数据整理,将氨基酸改变这一自变量进行数据处理,生成虚拟变量。导入所有数据后,进行GEE 分析。设定患者编码作为主变量。基于准似然独立准则(quasi-likelihood under the independence model criterion,QIC)选定最佳矩阵模型,在本研究中采用独立矩阵模型进行分析。由于因变量属于二分类数据,在回归分析时采用二元回归分析(Binary logistic)。选择不同自变量进行数据分析,研究不同氨基酸改变与表型之间的相关性关系。
2 结果
2.1 文献筛选结果
在进行文献检索以及筛选纳入过程后,共计纳入文献28 篇,收集MITF致病性突变位点共计34个。进一步于ClinicalVariant 数据库再次行MITF致病性突变位点检索,并经突变位点编号标准化后,最终建立包含58 个致病性突变位点的MITF突变数据库。并进一步收集到32个家系共计96名患者的临床表型,包括虹膜颜色、皮肤色素改变、头发颜色以及听力下降情况。
在58 例致病性突变位点中,以无义突变最多(21/58,36.2%),其次为错义突变(13/58,22.4%),移码突变(13/58,22.4%),剪切位点改变(4/58,6.90%),内含子替代(3/58,5.20%),缺失突变(2/58,3.40%),插入突变(2/58,3.40%)无义突变的突变位点分布较为均匀在各个外显子中均可见无义突变,而错义突变多位于7,8 号外显子(图2)。以上外显子均部分参与了MITF特征性结构域bHLH的编码。在突变位点的改变中则以单碱基改变为主,编码的蛋白以截短蛋白最为多见。而错义突变的编码氨基酸替代均在这一特征性结构域中。这一结构域中包含了DNA 结合域、MITF-M 的核定位信号(Nuclear location signal)以及二聚化的结合位点,乙酰化的修饰位点。错义突变导致的氨基酸替代均发生在bHLH结构中。
2.2 突变种类与表型的关系
依据突变基因编码蛋白的结构以及功能改变,将32个家系分为两类,共计7组。
听力下降作为WS 的主要临床特征,在96 名患者中,有75 名患者呈现出这一临床特征,约占78%,与既往文献报道中的89.6%的结果相近。其次为虹膜异色,约占66%。在这其中,以缺失核定位信号组出现比例最高。进一步将GEE 分析结果汇总于表2。就虹膜异色而言,有四种突变类型与虹膜异色的出现具有统计学差异;面部雀斑以及头发颜色异常均只与一种突变类型的相关性具有统计学差异;听力下降与氨基酸替换中的两种类型的相关性具有统计学差异,包括HLH结构阈以及NLS的氨基酸改变。
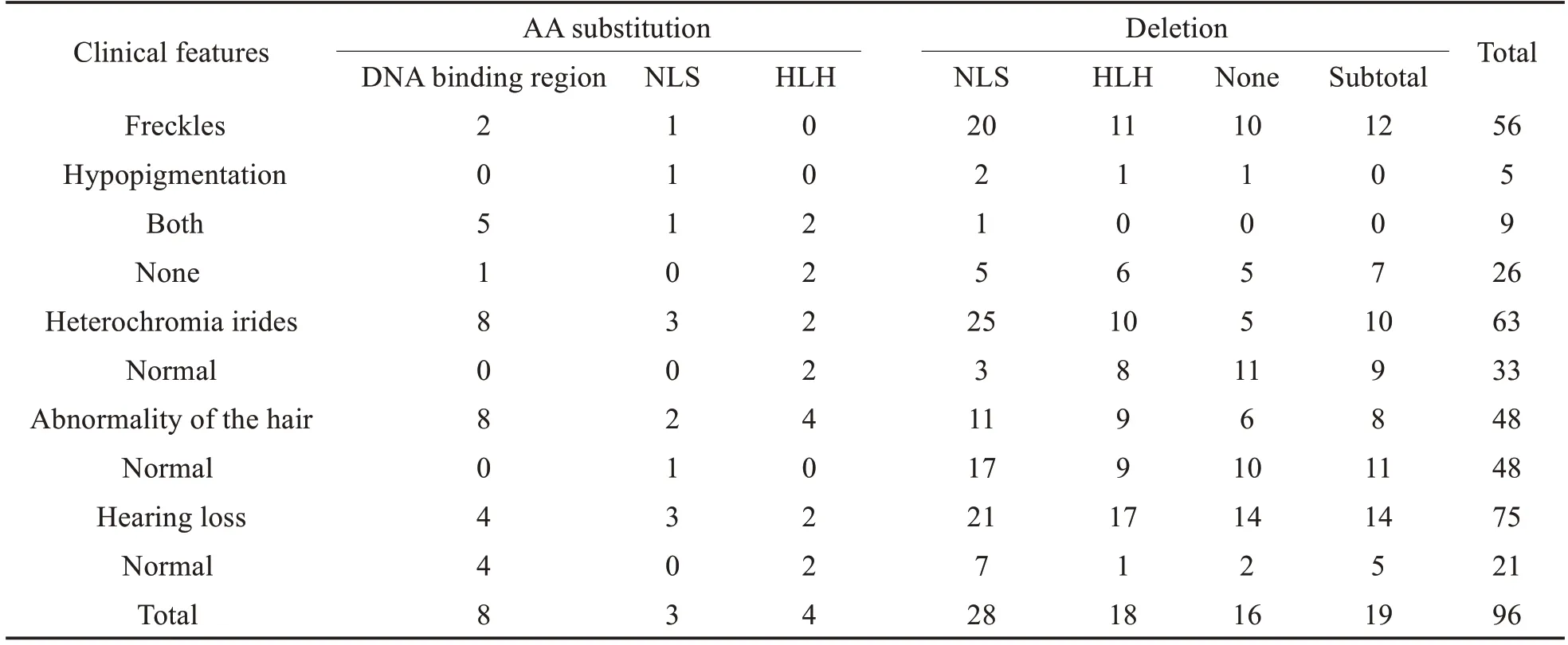
表1 临床表型汇总Table 1 Summary of clinical phenotypes
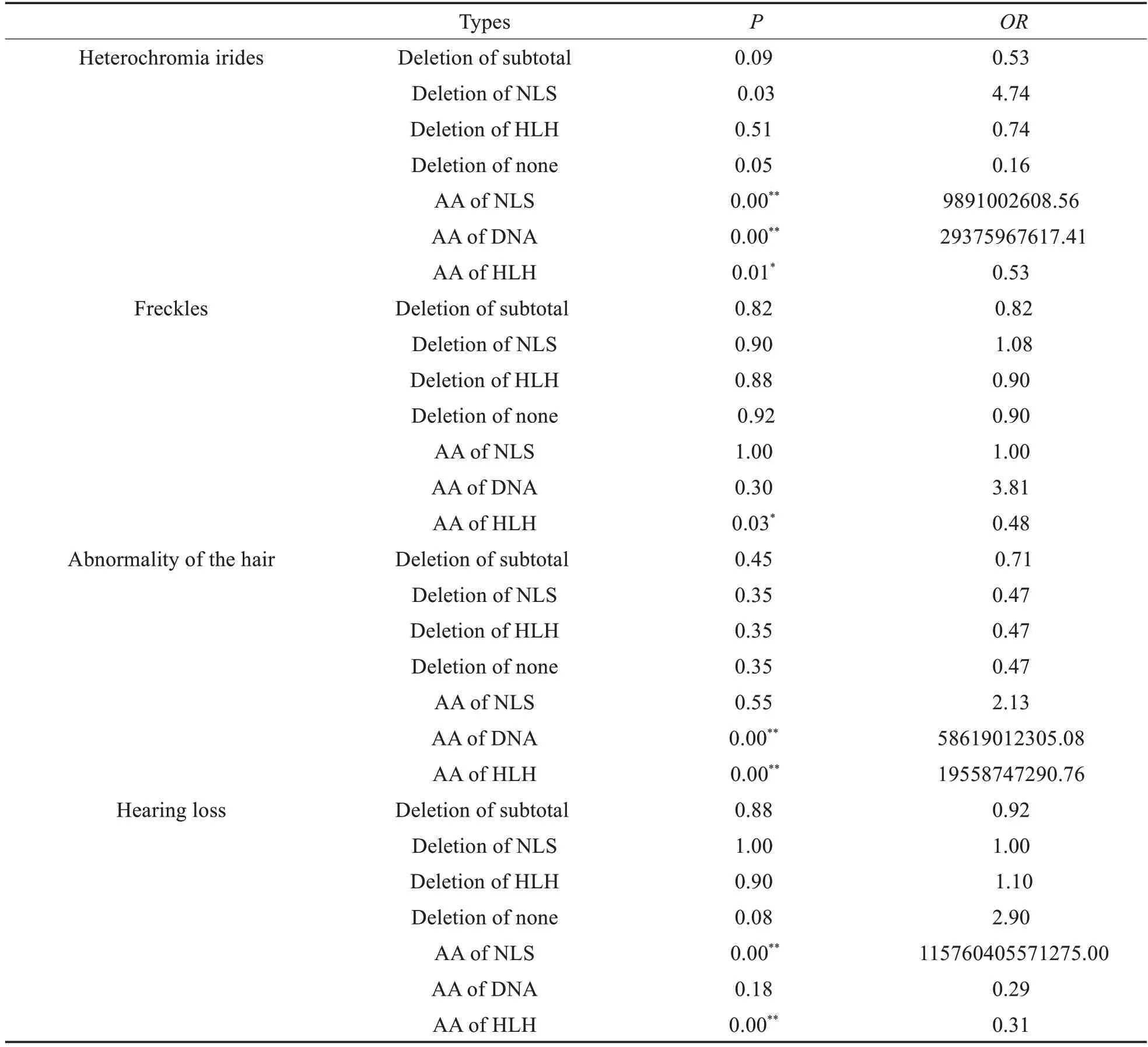
表2 统计结果汇总Table 2 Summary of statistical results
对于不同的突变类型来说,当突变类型为HLH 结构阈的氨基酸替代时,与头发颜色改变的出现呈现正相关,并具有统计学差异,而与虹膜异色、面部雀斑、听力下降呈现负相关,并具有统计学意义。
3 讨论
本研究通过建立MITF致病性突变数据库,以GEE的统计学方法进一步探索并分析MITF致病性突变与临床表型出现之间的关系。
研究结果显示,突变基因编码的截短蛋白与听力下降等均未见相关性。这可能与人体中的截短蛋白监控机制相关,即无义介导的mRNA 降解(nonsense-mediated decay,NMD)。鉴于NMD 在人体转录调节中的重要作用以及对mRNA 质量的监控机制[39],NMD 机制可以清除截短蛋白的mRNA,以减少截短蛋白的编码,降低截短蛋白的负性效应,并进一步调节遗传性疾病的相关表型[39]。
本研究中的错义突变则与临床表型的出现具有相关性,这一结果可能与MITF的作用过程以及作用特点相关。MITF致病性突变中错义突变位点大多位于MITF基因的外显子7,8,均影响MITF蛋白的bHLH-Zip结构阈。bHLH-Zip在MITF发挥作用的过程中具有十分重要的作用,主要参与蛋白质的二聚化,包括同源以及异源二聚化。MITF只有在形成二聚体后才可以与DNA 结合,发挥功能[7]。就MITF-M 而言,其主要在细胞核中发挥调节功能。因而核定位信号(nuclear localization signal,NLS)在MITF-M 的作用过程中具有重要作用。MITF-M 在NLS 介导下,经由核孔小体进入细胞核中,与DNA结合,进而发挥转录因子的调节作用[38]。鉴于HLH、NLS 的重要作用,故当错义突变位点位于以上结构的编码区时时,尽管未产生氨基酸数目的改变,但是影响了蛋白质的结构,编码蛋白质出现功能异常,导致疾病相关表型的出现。
本研究还进一步发现,听力下降与位于NLS以及HLH 的错义突变具有相关性。这与MITF-M 在听力形成的重要作用密切相关。目前有关WS 发病机制的研究多认为与神经嵴细胞的异常迁移以及分化相关,尤其黑色素细胞的分化[40],在黑色素细胞的相关调节通路中,大多通过MITF 将细胞内外的信号进一步转化,进而调节黑色素细胞的功能[41,42]。因而MITF在WS 致病机制中有着不可替代的作用。其中MITF-M 则是主要表达在黑色素细胞中的一种亚型[43]。MITF-M 不仅是黑色素细胞分化过程中的重要调节因子,还影响黑色素合成过程中的多种基因的表达水平[44,45],导致相关色素异常表型出现。MITF突变导致听力下降则与主要与中间细胞有关[46]。黑色素细胞在分化过程中迁移至内耳血管纹中,形成中间细胞,与边缘细胞以及基底细胞等其他种类细胞,共同组成血管纹,维持耳蜗内电位的稳定。研究表明影响血管纹中这三种主要细胞的突变基因可能通过影响耳蜗内电位,使得耳蜗内电位失衡,进而导致毛细胞凋亡,最终出现听力损失。尤其中间细胞中的内向回收钾离子通道Kir4.1 在耳蜗内电位的维持中有着必要的作用[47]。鉴于MITF在听力形成以及黑色素合成中的重要作用,当发生错义突变后,MITF的功能以及结构则会受到影响,导致临床表型的出现。结合以上研究,我们有理由推测HLH 以及NLS 在MITF-M作用过程中的重要作用。但仍需进一步实验进行验证。
MITF突变后由于单倍体剂量不足导致WS 相关临床表型的出现。但MITF在表达过程中受到多种正向以及负向调控,这些调控途径能否通过调节MITF表达水平而影响疾病表型仍需研究[48]。故而对于同一家系而言,患者出现不同的表型可能与多种调节途径相关。
本研究也具有一定的局限性。首先MITF数据库的建立过程中,未将患者的人群以及种族纳入考虑范围。因为不同的人群以及种族之间,相关临床表型的出现具有一定的差异。如面部雀斑则更易在亚洲人群当中出现。而在本研究纳入的人群中,亚洲人群较多,故可能存在一定的偏倚;其次突变类型之间纳入的患者数量具有一定的差异,会对结果产生一定的影响。尽管在数据分析时,采用的GEE方程进行分析,对各类型包含的患者数量不具有要求,但是错义突变的患者数量较少,可能具有一定程度的局限性;最后在进行表型分析时,由于一些明确突变位点的患者未获取到表型信息,所以未纳入进一步的研究中。而在这些人群中,较多突变属于错义突变,可能会对数据分析造成一定影响,进而产生偏倚。尽管我们研究中表明特定位置的突变类型与一些临床表型相关,但是这些表型出现的过程中不仅仅受到MITF的调控,还受到其他多种基因的调控。
本研究中,我们对MITF的突变位点与临床表型之间的相关性进行了初步分析,得出了表型与突变位点之间的初步分析结果,即致病性突变中错义突变影响MITF的特征性结构阈bHLH-Zip 编码区从而影响MITF蛋白的功能,导致疾病的出现。bHLH-Zip 结构阈中包含的DNA 结合域,NLS 等重要结构,在蛋白的作用过程中有着重要意义。尤其NLS 有着重要作用。位于NLS 以及HLH 编码区的错义突变,与WS中听力下降的出现具有相关性。
猜你喜欢
广西医科大学学报(2022年5期)2022-06-07
昆明医科大学学报(2022年3期)2022-04-19
中南医学科学杂志(2019年6期)2019-12-05
现代园艺(2017年21期)2018-01-03
湖南畜牧兽医(2016年3期)2016-06-05
兽医导刊(2016年12期)2016-05-17
中国康复理论与实践(2015年10期)2015-12-24
医学研究杂志(2015年5期)2015-06-10
现代检验医学杂志(2015年5期)2015-02-06
浙江医学(2014年17期)2014-04-13
