
NT-3对耳蜗螺旋神经节发育的调控机制研究进展
2023-05-22 06:47王雪艳侯书乐金永德孙莲花贾彦均权海燕金玉莲审校杨军
中华耳科学杂志 2023年2期
王雪艳 侯书乐 金永德 孙莲花 贾彦均 权海燕 金玉莲,*审校:杨军
1 延边大学附属医院耳鼻咽喉头颈外科(延吉 133000)
2 上海交通大学医学院附属新华医院耳鼻咽喉-头颈外科;上海交通大学医学院耳科学研究所;上海耳鼻疾病转化医学重点实验室(上海 200092)
3 青岛市中心医院耳鼻咽喉头颈外科(青岛 266000)
4 延边大学附属医院中心实验室(延吉 133000)
耳蜗螺旋神经元(spiral ganglion neurons,SGN)的发育需要各类神经营养因子的调控,其中神经营养素3(neurotrophin-3,NT-3)有着举足轻重的作用。NT-3 通过原肌球蛋白相关激酶C(tropomyosin receptor kinase C,TrkC)和p75 神经营养因子受体(neurotrophin receptor,p75NTR)相关的信号通路发挥促进细胞存活、突触可塑性、改变神经支配模式和再生髓鞘的作用,并且修复和逆转SGN发育过程中的损伤[1-3]。本文就NT-3 在SGN 发育中作用机制的研究进展综述如下。
1 NT-3在SGN发育中的作用
SGN 的发育过程依赖于多种神经营养因子的协同或拮抗作用,包括神经生长因子(nerve growth factor,NGF)、脑源性神经生长因子(brain derived neurotrophic factor,BDNF)、NT-3 和神经营养素4/5(neurotrophin-4/5,NT-4/5)等。研究发现敲除NT-3 基因的小鼠中SGN 的数量约减少了85%,因此认为NT-3 是SGN 发育中重要的神经营养因子,它主要来源于健康耳蜗内的毛细胞(hair cell,HC)和支持细胞(sertoli cells,SCs)[4]。其中SCs 中的NT-3 是维持SGN 存活和正常生理功能的关键因子,而HC中的NT-3 可以增加突触的密度。无论是SCs 源性还是HC 源性的NT-3,二者主要在耳蜗中发挥作用[5]。但NT-3 在耳蜗中如何分布,有待进一步研究。研究显示:当敲除SCs 源性NT-3 基因时,耳蜗基底部传入神经纤维的数量减少,因此,NT-3分布于耳蜗基底部高频区域的可能性较大;但是用β-半乳糖苷酶标记法探讨NT-3 分布的研究却发现,NT-3的梯度由顶回到低回逐渐降低[6]。
NT-3 参与SGN 发育的多个过程。SGN 来源于听基板(otic placode)和神经管,但以听基板为主要来源[7]。听基板为外胚层区域增厚的部分。在胚胎发育早期,该区域发生内陷形成听杯(otic cup),而听杯继续凹陷并从上皮细胞分离逐渐形成由假复层柱状上皮包裹的囊泡状结构,即耳囊(otocyst/otic vesicle)。耳囊前腹侧部分细胞发生迁移形成耳蜗-前庭神经节(cochleo-vestibular ganglion,CVG),CVG 神经母细胞迁移到蜗管的底部,为SGN 的发育做准备。在这个过程中NT-3参与了SGN的迁移和定位[8]。出生后12日(P12),Ⅰ型SGN 神经纤维发生突触修剪并与内毛细胞(inner hair cells,IHC)建立成熟的突触联系,标志着听力开始形成。此时主要是高水平的NT-3 诱导早期听力的获得[9]。出生后3 周(3W),Ⅱ型SGN 与外毛细胞(outer hair cell,OHC)建立成熟稳定的突触联系,听功能发育基本成熟。到成年时期,NT-3对噪音、耳毒性药物和年龄相关性听力损失,均有SGN损伤后的保护作用,因此NT-3 在成熟的突触结构中也发挥其生物学效应[10]。综上所述,自胚胎期到成年期,NT-3 在耳蜗中持续表达,不同时期参与不同的生物学过程,从而维持着SGN的正常发育与损伤后的修复。Jin 等[9]对P0、P5 和P20 三个时期SGN 进行体外细胞培养,培养基中分别添加NT-3、BDNF、白血病抑制因子(leukemia inhibitory factor,LIF),NT-3 组、BDNF 组、NT-3+BDNF 组和NT-3+BDNF+LIF 组作为实验组,与未添加任何神经营养因子作为对照组。结果发现在P0 和P5 期,三种神经营养因子联合使用组(NT-3+BDNF+LIF组)的SGN的存活率明显高于单独使用神经营养因子(NT-3 组和BDNF 组)或联合使用两种神经营养因子组(NT-3+BDNF组);而在P20 期,单独使用神经营养因子组(NT-3 组和BDNF 组),SGN 的存活率高于两种营养因子联合(NT-3+BDNF 组)或三种神经营养因子联合使用组(NT-3+BDNF+LIF 组),从而证实了SGN 在耳蜗的发育早期高度依赖外源性神经营养因子,且在发育后期对NT-3 的依赖性更加突出。在此基础上,又进行体内实验[11],利用Ad-GFP 作为NT-3的载体注入到P5期大鼠耳蜗鼓阶中,结果发现P10、P15、P20期大鼠耳蜗底回、中回和顶回的NT-3mRNA表达水平明显高于正常组,且范围逐渐扩大到神经纤维和血管纹,进而在SGN 内发挥广泛作用;与同期未转染NT-3的大鼠相比,转染后的大鼠ABR阈值降低。因此,在SGN 发育早期及时补充外源性NT-3 可以有效避免因其不足导致的听力损失。
目前体内实验[12,13]和体外实验[11,14,15]的研究已证实NT-3 加速SGN 轴突的延伸速度,并促进SGN 数量和密度的增加[16],但其作用机制尚不清楚,下文中将从受体和信号通路阐述其在SGN 发育中的作用机制。
2 NT-3促进SGN发育的机制
HC 和SCs 产生的NT-3 经轴突运送到SGN 后,与酪氨酸激酶受体TrkC 和p75NTR 结合发挥生物学作用。TrkC 与p75NTR,与SGN 的结合方式不同,TrkC 与SGN 特异性结合,p75NTR 与SGN 非特异性结合。TrkC 是NT-3 浓度依赖性受体,当缺乏NT-3时,TrkC被半胱氨酸酶切割发挥促凋亡作用;但当NT-3 充足时,TrkC 受体促进SGN 分化发育的作用掩盖了p75NTR的促凋亡作用[17]。TrkC受体通过RAS-丝裂原激活细胞外信号调节激酶(MEK1/2)-细胞外信号调节激酶(ERK1/2)信号通路和磷脂酰肌醇3 激酶(PI3K)信号通路,将信息传递到细胞内,从而调控SGN的发育。
p75NTR 的存在增强了NT-3 与TrkC 结合的特异性,当缺乏TrkC 受体时,NT-3 通过p75NTR 诱导细胞凋亡;除此之外,p75NTR也具有促进神经细胞增殖和突触生长的作用[18]。免疫荧光染色结果发现,p75NTR在耳蜗神经胶质细胞中表达,主要是在雪旺细胞,而在SGN的胞体和突触中未见表达,因此p75NTR在SGN中,可能是通过形成髓鞘从而起到保护作用,其参与的信号通路可能是为NT-3/p75NTR/应激活化蛋白激酶(JNK)通路和NT-3/p75NTR/p38丝裂原活化蛋白激酶(p38 MAPK)通路。
2.1 TrkC介导的信号通路
2.1.1 RAS-MEK1/2-ERK1/2信号通路
RAS-MEK1/2-ERK1/2信号通路是丝裂原活化蛋白激酶(MAPK)家族的亚家族之一,其作用是将细胞外的信号逐级放大并传递到细胞核,再通过转录因子完成各种生命活动。当NT-3 与TrkC 受体结合后,TrkC 受体发生二聚化导致自身激活发生自身的磷酸化,为下游信号蛋白提供锚定位点,GRB2-SOS 复合物与激活后的受体结合后导致Ras蛋白由结合二磷酸鸟苷(GDP)转为结合三磷酸鸟苷(GTP)而自身活化,将胞浆中的Raf 蛋白募集到质膜上,并诱导其构象的改变。激活后的Raf 并逐级磷酸化MEK1/2 和ERK1/2。ERK1/2 平时位于胞质内,当被激活后进入核内与转录因子结合,使转录因子活化,活化后的转录因子结合到DNA 上启动下游靶基因的表达,实现生理功能;除此之外,活化后的ERK1/2 也可以磷酸化胞质和核内的激酶[19]。SGN 体外细胞培养发现,NT-3 通过RASMEK1/2-ERK1/2 信号通路促进SGN 发育。下面将从细胞迁移、细胞分化发育和细胞凋亡三个阶段,阐述NT-3 通过TrkC 受体经RAS-MEK1/2-ERK1/2信号通路对SGN发育的作用机制(图1)。
在神经元发育过程中,NT-3 可能与TrkC 受体结合,通过RAS-MEK1/2-ERK1/2 信号通路,参与了神经元的迁移。Yamauchi等[20]认为NT-3 增强了TrkC 受体磷酸化Dbs的能力,诱导了周围神经系统中雪旺细胞的迁移。沿SGN 纤维分布的雪旺细胞表达NT-3,因此雪旺细胞为SGN的再生带来可能。Hossain 等[8]认为起源于神经嵴的SGN 从听泡迁出后紧贴着耳蜗上皮自蜗底沿着蜗管向顶端延伸,其轴突穿透上皮并发生修剪,与HC建立连接,在这个过程中NT-3参与了SGN的迁移和定位。因肾小球足细胞表达神经元标志性蛋白,所以被用作神经元研究的替代物,研究发现NT-3 通过激活TrkC 经ERK 通路增加了足细胞的迁移,因此推测NT-3 可能与TrkC 受体结合通过RAS-MEK1/2-ERK1/2 信号通路参与SGN的迁移。
除此之外,NT-3 与TrkC 受体结合通过RASMEK1/2-ERK1/2 信号通路促进神经元分化发育。Kevin 等[21]研究发现MIAMI(marrow isoled adult multilineage inducible)细胞在体外具有分化为多种神经元细胞的潜能,分化后的神经元细胞表达βIII-微管蛋白神经元标志物和具有成熟神经元细胞的电生理特性,并且细胞表面唯一表达TrkC受体,这种细胞的分化潜能完全依赖于NT-3诱导ERK1/2短暂激活(5至15分钟)的级联反应。最新研究认为在大鼠海马神经元中,NT-3通过TrkC受体的二聚化启动细胞内的信号通路促进了TrkC-PTPσ复合物的相互作用,诱导了SGN海马神经元突触的分化过程[22,23]。研究还发现RAS-MEK1/2-ERK1/2信号通路具有诱导小鼠胚胎干细胞分化的潜能,因此,在SGN分化过程中,NT-3 可能通过TrkC 受体经RAS-MEK1/2-ERK1/2信号通路发挥作用。
NT-3 与TrkC 受体结合通过RAS-MEK1/2-ERK1/2 信号通路,还可以诱导了神经元细胞凋亡。Saini 等[24]认为在少突胶质细胞增殖中,NT-3 浓度和抗凋亡蛋白Bcl-2 表达水平呈现正相关,协同起促进作用。在无神经营养因子维持时,抗凋亡蛋白Bcl-2 足以维持SGN 的存活[25];虽然TrkC 和Bcl-2均表现为NT-3 浓度依赖性,但NT-3 存在时,MEK1/2和Bcl-2过表达导致SGN死亡,可能是NT-3过度激活TrkC受体通过RAS-MEK1/2-ERK1/2信号通路诱导细胞凋亡的能力大于Bcl-2 维持SGN存活的能力[26]。
2.1.2 PI3K信号通路
在SGN 中,PI3K 直接或者间接接受TrkC 受体信号转导,起到促存活和促突触作用。由于细胞间存在着极其复杂的信号通路间的交流,Grb2与SOS形成的信号转导复合物,在RAS-MEK1/2-ERK1/2信号转导与PI3K/丝氨酸-苏氨酸激酶(Akt)信号转导中起到桥梁作用,所以RAS 也可以直接激活PI3K[27]。Aletsee 等[28]在SGN 细胞培养中通过添加NT-3 诱导神经突起的数量和长度显著增加,而使用PI3K 抑制剂明显减弱这种促进作用,因此PI3K也是NT-3/TrkC 调节SGN 发育过程中的关键酶(图1)。PI3K相关的信号通路众多,如PI3K/AKT/哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)通路和PI3K/AKT/环磷腺苷效应元件结合蛋白(CREB)通路等,抑制mTOR 或者过表达CREB 均可以提高SGN 的存活率和突触生长,但NT-3/TrkC具体是通过哪种信号通路仍有待验证。
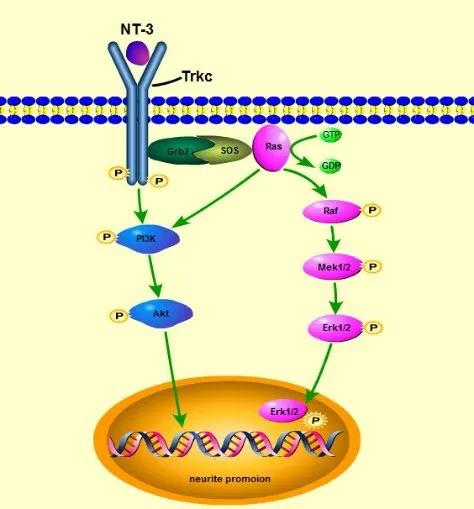
图1 NT-3/TrkC信号通路示意图Fig.1 Schematic diagram of the NT-3/TrkC signal pathway
2.2 p75NTR介导的信号通路
2.2.1 JNK通路
JNK 也是MAPK 信号通路中的一个亚型,在细胞因子和细胞应激等多种生命活动中发挥作用。JNK 对SGN 的保护作用包括促进细胞凋亡和突触生长,这是由JNK 在细胞中不同定位决定的-通常细胞核中的JNK 促进细胞凋亡,而细胞质中的JNK促进突触生长。JNK 分为JNK1、JNK2和JNK3三个亚型,而JNK3 在诱导SGN 凋亡中起主导作用[29],敲除JNK3 后抑制了HC 丢失后SGN 的凋亡,因此使用JNK3 抑制剂成为SGN 损伤后的潜在治疗方式[30]。然而无外界刺激或NT-3 充足时,JNK1、JNK2 和JNK3 诱导了SGN 突触的形成并维持其生长[2]。抑制JNK 信号通路,SGN 外植体神经突触的长度和数量均减少,但通过添加NT-3 可以逆转[31,32]。因此,NT-3促进神经元存活的机制可能是通过抑制细胞核JNK 的磷酸化,从而抑制SGN 细胞的凋亡;或者是促进胞质内JNK 的磷酸化而促进SGN 的轴突生长和存活[33],此外还有观点认为NT-3 与p75NTR 的结合降低后,JNK 的蛋白表达明显降低,即NT-3 对SGN发育的调节作用可能是通过p75NTR/JNK通路发挥作用[34]。
2.2.2 P38MAPK通路
P38MAPK 信号通路参与中枢神经系统和外周神经系统中髓鞘形成和修复,它通过在成熟少突胶质细胞[35]和雪旺细胞[36]中发挥作用,保持了神经元周围髓鞘的完整性,促进了神经元的存活[37]。除此之外,此通路还与突触可塑性密切相关[38]。Mullen等[39]认为在SGN中,BDNF通过P38信号通路对其突触数量起促进作用,然而Aletsee等[40]认为P38不能通过NT-3影响SGN,根据NT-3与TrkC结合特异性强于p75NTR的特点,推测可能原因是NT-3优先与TrkC结合,导致NT-3/TrkC 介导的信号转导途径抑制NT-3/p75NTR/P38MAPK信号转导途径所致。
3 小结与展望
SGN 在HC 或SCs 损伤后仍可以存活数周到数月,而且TrkC 受体和p75NTR 在成年后的SGN 中仍持续表达,这为SGN 损伤后,或干细胞治疗和人工耳蜗植入后[41-42],补充外源性NT-3 以维持SGN 存活并发挥生物学功能提供理论依据。虽然BDNF在一定程度上可以代替NT-3,但是这些神经营养因子激活的信号通路却大不相同,毋庸置疑NT-3是耳蜗发育不可或缺以及无法替代的神经营养因子,深入了解NT-3 在耳蜗发育过程中的作用及机制,可为以后将其应用于临床提供理论依据。
猜你喜欢
自然杂志(2021年6期)2021-12-23
中国听力语言康复科学杂志(2021年6期)2021-12-21
中华家教(2018年7期)2018-08-01
现代装饰(2018年5期)2018-05-26
海南医学(2016年8期)2016-06-08
文学少年(原创儿童文学)(2016年16期)2016-02-28
听力学及言语疾病杂志(2015年5期)2015-12-24
电源技术(2015年5期)2015-08-22
弹箭与制导学报(2015年1期)2015-03-11
中国当代医药(2015年7期)2015-03-01
