
芳香烃受体调控人脂肪间充质干细胞的免疫调节功能
2023-06-05 06:43武文婧高竞溪赵晓妍赵春华
基础医学与临床 2023年6期
武文婧,高竞溪,赵晓妍,孙 昭,韩 钦*,赵春华*
1.中国医学科学院基础医学研究所 北京协和医学院基础学院 中国医学科学院组织工程研究中心,北京 100005;2.中国医学科学院 北京协和医学院 北京协和医院 肿瘤内科,北京 100730
间充质细胞(mesenchymal stem cells, MSCs)具有自我更新、多向分化和免疫调节能力,MSCs可从多种组织中分离获得,如脂肪、骨髓、胎盘、脐带、和牙髓等。国际细胞治疗学会(ISCT)定义经典培养条件下的MSCs为贴壁增殖,高表达CD105、CD73和CD90,低表达CD45、CD34、CD14、CD11b、CD19和HLA-DR,能够在体外分化为成骨细胞、脂肪细胞和软骨细胞的细胞[1]。MSCs的免疫调节能力是其临床应用于免疫系统相关疾病治疗的主要药效机制[2-3],因此更加深入地研究MSCs发挥免疫调节功能的机理至关重要。目前的研究认为,间充质干细胞发挥免疫调节作用主要通过分泌可溶性因子,如研究较多的吲哚胺2,3双加氧酶(indoleamine2,3-dioxygenase1, IDO1)、肿瘤坏死因子刺激基因-6(tumor necrosis factor-α stimulated gene 6, TSG6)和前列腺素2(prostaglandin E2, PGE2)等[4-5]。IDO1被认为是打开人MSC免疫调节功能的关键分子[6]。IDO1是一种重要的色氨酸分解酶,在肿瘤高表达,色氨酸代谢产物通过芳香烃受体(aryl hydrocarbon receptor,AHR)通路抑制肿瘤免疫。2020年发表在Cell上的一项研究报道IL4I1是除IDO1以外在肿瘤中的重要色氨酸分解酶[7],也通过AHR通路抑制肿瘤免疫。AHR是一种配体激活的转录因子,它整合环境、饮食、微生物和代谢相关因素[8],由许多免疫细胞表达,AHR信号提供了一个整合环境和代谢对免疫反应影响的分子途径[9]。在色氨酸代谢中,IDO1和IL4I1将色氨酸(tryptophan,Trp)转化为犬尿氨酸(kynurenine, Kyn),并通过激活Kyn-AHR代谢通路发挥免疫调节作用。因此,本研究拟深入探讨AHR参与间充质干细胞免疫调节作用的分子机制。
1 材料与方法
1.1 材料
1.1.1 主要试剂:IFN-γ、poly(I:C)、佛波酯(PMA)和氯唑沙宗(CZ)(MCE公司);TGF-β(北京科昕生物科技有限公司);山羊抗兔、山羊抗小鼠二抗和脂多糖(LPS)(均上海碧云天生物技术有限公司);Lipofectamine 3000(Invitrogen公司);DMEM/F12、RPMI-1640和胎牛血清(Gibco公司);siRNA(苏州吉玛基因股份有限公司);引物(上海生物工程有限公司);反转录试剂盒(Takara公司);ELISA试剂盒(江苏酶免实业有限公司)。
1.1.2 细胞:脂肪来源间充质干细胞(hAD-MSCs),取自中国医学科学院整形外科医院吸脂术患者废弃的脂肪组织,经体外分离培养获得。本研究经中国医学科学院基础医学研究所伦理委员会批准,供者均签订知情同意书。人单核细胞白血病细胞系THP-1(中国医学科学院基础医学研究所细胞资源中心)。
1.2 方法
1.2.1 细胞的分离和培养:提取和培养人脂肪MSCs的方法参见文献[10]。
1.2.2 利用佛波酯将THP-1诱导为巨噬细胞:将THP-1以5×105/mL接种到6孔板中,佛波酯的终浓度为100 μg/L,培养24 h后细胞由悬浮状态转为贴壁增殖,待与间充质干细胞共培养使用。
1.2.3 间充质干细胞的瞬时转染:使用Lipofectamine 3000,按照说明书进行脂肪MSCs的瞬时转染,转染后的细胞命名为si-NC、si-IDO1、si-IL4I1和si-AHR,供与巨噬细胞共培养使用。
1.2.4 间充质干细胞与巨噬细胞共培养:在间充质干细胞对巨噬细胞活化的抑制实验中,使用LPS刺激巨噬细胞活化,LPS的终浓度为100 μg/L,同时使用0.4 μm孔径的Transwell小室将间充质干细胞或其条件培养基加入培养体系中,共培养后收集巨噬细胞以待检测。
1.2.5 RT-qPCR检测mRNA:收集未经处理的巨噬细胞、单独使用LPS活化的巨噬细胞、经LPS活化并与间充质干细胞共培养的巨噬细胞,检测炎性因子IL-6、TNF-α、IL-1β、IL-10的表达水平,引物序列见表1。

表1 RT-qPCR引物Table 1 Primer sequences for RT-qPCR
1.2.6 蛋白免疫印迹(Western blot)检测胞内蛋白:间充质干细胞芳香烃受体的蛋白水平检测方法参见文献[10]。
1.2.7 免疫荧光染色检测胞内蛋白:间充质干细胞色氨酸代谢关键代谢物的免疫荧光染色方法参见文献[11]。
1.2.8 酶联免疫吸附测定(ELISA)检测细胞分泌蛋白:收集巨噬细胞上清液,按照ELISA试剂盒中的说明书进行检测。
1.3 统计学分析
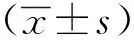
2 结果
2.1 不同炎性因子对间充质干细胞色氨酸代谢通路关键基因表达的影响
MSCs未经炎性因子刺激时,几乎不表达IDO1和IL4I1蛋白(图1A,B),而AHR表达较高,免疫荧光显示AHR在细胞核中也有表达(图1C)。IFN-γ、TGF-β、poly(I:C)和CZ[12]刺激MSCs后,IDO1、IL4I1、AHR和CYP1B1在不同时间表达上调(P<0.05)(图2A~D),IFN-γ的效果最显著(P<0.001)(图2A)。因此,后续实验选择IFN-γ诱导方案验证IDO1、IL4I1及AHR对MSCs免疫调节功能的影响。

A-C.immunofluorescence analysis of MSCs, stained with anti-IDO1, anti-IL4I1 (green) and anti-AHR (red) antibodies; nuclei were stained with DAPI (blue).图1 正常培养的MSC色氨酸代谢关键酶IDO1和IL4I1表达水平低,AHR表达水平高Fig 1 Low expression of IDO1 and IL4I1 and high expression level of AHR in normal cultured MSCs

A-D.RT-qPCR analysis of tryptophan metabolism related gene IDO1, IL4I1, AHR and CYP1B1 in MSCs stimulated by IFN-γ(A), TNF-α(B), poly(I:C)(C) and CZ(D); *P<0.05, **P<0.01, ***P<0.001 compared with untreated MSCs (0 hour).图2 炎性细胞因子刺激MSCs不同时间点色氨酸代谢酶及AHR表达水平上调Fig 2 Inflammatory cytokines stimulated the upregulation of tryptophan metabolizing enzyme and AHR expression levels at different time in
2.2 确定巨噬细胞经LPS活化后,炎性因子表达水平最高的时间
研究MSCs对巨噬细胞的免疫调节作用时,选择THP-1细胞系进行实验,使用PMA将THP-1诱导成巨噬细胞,经LPS活化后与MSCs共培养。首先,检测巨噬细胞活化的特征性炎性因子的表达时相。巨噬细胞活化的3个特征基因IL-6、TNF-α和IL-1β在LPS活化后3 h左右开始上调(P<0.001),TNF-α持续到12 h都维持较高的表达水平(P<0.05),IL-6和IL-1β持续到72 h都维持较高的表达水平(P<0.001)(图3)。因此,在后续共培养的实验中,观察时间点可以选择刺激后3~12 h之间。

RT-qPCR analysis expression of IL-6, TNF-α and IL-1β in macrophage stimulated by LPS at 0, 1, 3, 6, 12, 24, 48, 72 hours, *P<0.05, **P<0.01, ***P<0.001 compared with untreated MSCs (0 hour).图3 LPS刺激巨噬细胞后炎性细胞因子表达上调Fig 3 Upregulation of inflammatory cytokine expression after LPS stimulation of
2.3 间充质干细胞中芳香烃受体表达情况
收集正常培养的MSCs上清,作为条件培养基加入LPS刺激的巨噬细胞中,定量分析巨噬细胞炎性因子的表达,发现未经炎性因子刺激的MSCs培养上清可以显著抑制巨噬细胞炎性因子IL-6(P<0.001),TNF-α(P<0.01)和IL-1β(P<0.01)的表达(图4A)。

A.RT-qPCR analysis inflammatory cytokines IL-6, TNF-α and IL-1β in macrophage stimulated by LPS and co-culture with MSCs condition medium,*P<0.01, **P<0.001 compared with LPS, #P<0.01, ##P<0.001 compared with M0; B.Western blot detected AHR in MSCs stimulated by IFN-γ on 24, 48,72 hours, **P<0.001 compared with untreated MSCs (0 hour); C.immunofluorescence analysis of MSCs, stained with anti-AHR (red) antibodies.nuclei were stained with DAPI (blue).图4 IFN-γ刺激后AHR表达及核分布增加Fig 4 Increased AHR expression and nuclear distribution after IFN-γ
由于MSCs未经炎性因子刺激时,几乎不表达IDO1和IL4I1蛋白,而AHR表达较高且存在核分布的情况,这意味着不经过炎性细胞因子的激活,就存在Kyn-AHR的活化,因此推测MSCs中可能存在除了IDO1和IL4I1以外的色氨酸代谢酶,或者其他的AHR激活剂。IFN-γ刺激后AHR蛋白表达水平上调(图4B),细胞核分布增加(图4C)。因此,未经炎性因子刺激的MSCs可能由于AHR的表达和活化从而发挥免疫调节作用。
2.4 IDO1和IL4I1的表达对间充质干细胞免疫调节作用的影响
分别使用si-IDO1、si-IL4I1及IFN-γ处理MSCs,转染后IDO1、IL4I1的表达水平明显下调(P<0.01)(图5A)。收集上述处理后的MSCs条件培养基,分别加入LPS刺激的巨噬细胞体系中。结果表明,与对照组(LPS+si-NC)相比,IDO1和IL4I1干扰后的MSCs抑制巨噬细胞炎性因子的能力减弱(P<0.05)(图5B)。因此,IDO1和IL4I1可能都参与调控MSCs的免疫调节功能。

A.the expression of IDO1 and IL4I1 was determined by RT-qPCR in MSCs transfected with si-IDO1, si-IL4I1 and si-NC, **P<0.01, ***P<0.001 compared with si-NC; B.RT-qPCR analysis inflammatory cytokines IL-6, TNF-α and IL-1β in macrophage stimulated by LPS and co-culture with MSCs condition medium which transfected with si-IDO1, si-IL4I1 and si-NC, *P<0.05, **P<0.01, ***P<0.001 compared with LPS+si-NC, #P<0.01,##P<0.001 compared with M0,ΔP<0.01, ΔΔP<0.001 compared with LPS.图5 低表达IDO1和IL4I1的MSCs免疫抑制功能减弱Fig 5 Reduced immunosuppressive function of MSCs with low expression of IDO1 and
2.5 AHR对间充质干细胞免疫调节作用的影响
使用si-AHR进行转染,收集对照组及AHR干扰组的间充质干细胞,与对照组相比,干扰组的AHR在mRNA水平显著下调(P<0.05)(图6A),在蛋白水平也明显下调(图6B)。收集上述处理后的MSCs条件培养基,分别加入LPS刺激的巨噬细胞体系中。结果表明,在mRNA表达水平上,与对照组(LPS+si-NC)相比,AHR干扰后的MSCs抑制巨噬细胞炎性因子的能力减弱(P<0.05)(图6C);ELISA检测巨噬细胞培养上清,得到与基因表达水平一致的结果(P<0.001)(图6D)。因此,AHR对MSCs发挥免疫调节功能具有重要作用。

A.the expression of AHR was determined by RT-qPCR in MSCs transfected with si-AHR and si-NC, *P<0.05 compared with si-NC; B.Western blot detected AHR in MSCs transfected with si-AHR and si-NC, ***P<0.001 compared with si-NC; C.RT-qPCR analysis inflammatory cytokines IL-6, TNF-α and IL-1β in macrophage stimulated by LPS and co-culture with MSCs condition medium which transfected with si-AHR and si-NC, *P<0.05, **P<0.01, ***P<0.001 compared with LPS+si-NC, #P<0.001 compared with M0, ΔP<0.01, ΔΔP<0.001 compared with LPS; D.ELISA detected inflammatory cytokines IL-6, TNF-α and IL-1β in macrophage stimulated by LPS and co-culture with MSCs condition medium which transfected with si-AHR and si-NC, ***P<0.001 compared with LPS+si-NC, #P<0.001 compared with M0, ΔP<0.01, ΔΔP<0.001 compared with LPS.图6 低表达AHR的MSCs免疫抑制功能减弱Fig 6 Reduced immunosuppressive function of MSCs with low expression of
3 讨论
间充质干细胞具有免疫调节功能已成为共识,但人们对MSCs发挥免疫调节功能的机制仍没有明确的认识[13]。时玉舫等人[6]于2014年提出了“IDO是人MSCs免疫调节功能开关”的假说:高水平的促炎因子存在时,MSCs会表达高水平的IDO,展现出更强的免疫抑制能力。IDO在MSCs中发挥免疫调节作用的下游分子尚不清楚,那么在MSCs中,IDO1是否可以通过代谢产物作用于AHR发挥免疫调节作用?此外,新报道的色氨酸代谢酶IL4I1是否也参与MSCs的免疫调节作用?带着这两个问题,进行了刺激和未刺激的MSCs培养上清对巨噬细胞的调节作用的分析。研究发现正常培养条件下的MSCs上清就具有免疫抑制功能,免疫荧光染色证实正常培养的MSCs高表达AHR,且部分分布在细胞核,但几乎不表达IDO1和IL4I1。下调正常的MSCs的AHR表达,可以显著削弱MSCs的免疫调节作用。上述结果提示AHR参与正常MSCs的免疫调节作用,但AHR的激活机制有待进一步研究。
有研究表明,使用炎性因子或药物处理的方法模拟炎性微环境,可以启动MSCs的免疫抑制功能,使其发挥强大的免疫调节作用[14-15]。在研究中,发现色氨酸代谢关键酶IDO1和IL4I1都参与炎性因子刺激对MSCs免疫调节功能增强的过程。正常的MSCs几乎不表达色氨酸代谢酶,经不同炎性因子或药物刺激的MSCs的色氨酸代谢酶表达上调, 其中IFN-γ刺激可同时显著上调IDO1和IL4I1表达,TGF-β,poly(I:C)和CZ只能激活IL4I1表达上调,提示IDO1和IL4I1两个色氨酸代谢酶都参与了不同刺激后MSCs的免疫调节功能增强,但不同刺激剂诱导代谢酶表达的分子机制尚有待进一步研究。
综上所述,AHR正向调节MSCs的免疫功能,下调AHR可以抑制MSCs的免疫调节作用,炎性因子刺激通过上调色氨酸代谢酶的表达增强MSCs的免疫调节能力。正常MSCs中AHR的激活机制也有待于后续的研究。
猜你喜欢
北方牧业(2022年9期)2022-11-22
宁夏医学杂志(2020年3期)2021-01-21
动物营养学报(2018年12期)2018-12-13
天然产物研究与开发(2018年4期)2018-05-07
中成药(2017年5期)2017-06-13
天然产物研究与开发(2016年6期)2016-06-05
中成药(2016年8期)2016-05-17
中国医疗美容(2015年5期)2015-02-03
饲料博览(2014年11期)2014-05-04
现代检验医学杂志(2014年3期)2014-02-02
