
海洋初级生产力与输出生产力的时滞效应研究进展
2023-06-13 13:36肖武鹏黄邦钦
厦门大学学报(自然科学版) 2023年3期
鲍 敏,肖武鹏,黄邦钦
(厦门大学环境与生态学院,近海海洋环境科学国家重点实验室,福建省海陆界面生态环境重点实验室,福建 厦门 361102)
海洋是地表系统最大的碳库,其储碳量是大气的60倍,是陆地生态系统的20倍,因此在全球碳循环中发挥重要作用[1].海洋在碳循环的过程中,吸收大气中的CO2,通过自身内部一系列的生物化学作用,将无机碳转换为有机碳并输送至海洋深层.随着全球CO2含量的增加,海洋的储碳量也经历了相当大的年际变化.从1998年到2011年,科学家预估海洋的碳吸收量每年都增加(1.00±0.12)Gt[2].近10年(2010—2019年)海洋每年平均吸收2.5 Gt碳(即10 Gt CO2),约占人为CO2排放量的26%,起到长期储存CO2和调节气候变化的作用[3-6].
目前,普遍认为海洋的储碳能力是由生物泵、溶解度泵、碳酸盐泵以及微生物碳泵共同决定的[7-8].其中生物泵和溶解度泵是海洋储碳的主要维持机制,它们共同调控着溶解无机碳(DIC)从海表到深海的垂直梯度[9-10].溶解度泵是将温度较低且富含DIC的高密度海水在重力作用下输送至深海的物理化学过程,主要发生在高纬度海区.生物泵则是将颗粒有机碳(POC)从海表输送到深海的一系列生物过程,将大气中的CO2长期封存至深海,在全球海区都发挥作用[7,11].CO2从大气进入上层海洋,在真光层被浮游植物通过光合作用转化为POC,之后伴随着食物网中不同营养级的生物相互作用,进行物质和能量运输.在此过程中,一些未被消耗的浮游植物形成的聚集体和浮游动物代谢产生的粪便颗粒迅速下沉,在下沉过程中进一步被微生物群落再矿化,最终只有少部分有机物进入深海,实现碳向深海的输出.
生物泵主要通过生物重力泵(BGP)、迁移泵、混合泵3个途径运行[12].BGP是指POC以聚集体、浮游动物的尸体和粪便颗粒等形式发生的被动沉降.普遍认为,BGP贡献了垂直DIC梯度的90%左右,在上层海洋的碳固定、中层生物群落的维持以及海洋内部的碳存储上发挥重要作用[9-10,13].迁移泵是通过浮游动物和鱼类的昼夜垂直迁移将有机碳主动运输至深海的过程[14-16].混合泵包括通过垂直混合、季节性混合层加深、涡流和大尺度海洋环流对悬浮POC和溶解有机碳(DOC)的物理输送[10,17-19].据估计,生物泵每年可从海洋上层清除100亿t以上的碳,其驱动力来自于上层海洋的沉降有机颗粒[20].这些颗粒物的最初来源是浮游植物的初级生产力(PP),即浮游植物通过光合作用将无机碳(CO2)转换为POC,其中大部分被上层海洋的代谢过程再矿化,仅有一小部分输出真光层进入弱光层(100~1 000 m),即输出生产力(EP).碳从上层海洋输出真光层的效率称为生物泵输出效率,用e-ratio表示,等于EP与PP的比值.e-ratio在一定程度上决定了CO2的海气分配[21],因此,量化e-ratio是理解全球碳循环的一个重要因素.尽管国际学界已知e-ratio的估算受EP与PP变化不同步,即EP与PP之间时滞的影响,但是二者时滞的估算存在较大不确定性,且不同海域造成时滞的原因尚不明确[22-23].在地质时间尺度上,总PP和EP的变化会极大地影响大气中的CO2水平,从而影响全球气候,并为可持续的渔业捕捞设定了上限[20].而由于海洋中浮游植物碳的平均周转时间约为一周或更短,PP和EP对外部因素的变化极其敏感,很少处于稳定状态[20].因此,阐明EP与PP之间的时滞效应对于理解海洋生物如何应对自然气候变化,以及海洋生态如何在大气强迫和气候反馈的背景下发挥作用至关重要,并可为未来10年如何管控人类活动对海洋产生的影响提供思路.
1 EP与PP的时滞效应
自1984年在美国开始的全球海洋通量研究及1987年的全球海洋通量联合研究之后,科学家们发现了生物泵在调控海气CO2平衡中的重要性,越来越多的研究开始探寻如何确定生物泵的效率及碳输出过程和通量.然而在实际研究中,由于PP与EP的测量时间尺度不一致,想要同步测定EP与PP来获得e-ratio的现场估算相当困难.因此,开发通过遥感参数估算e-ratio的简单经验算法,以估算全球尺度、高时空分辨率的有机碳输出通量,一直是生物泵研究的热点.目前已有的经验算法大多基于少量现场实测EP数据,并将e-ratio 表示成海表温度(SST)、PP或叶绿素的函数[22],其预测结果通常显示EP随PP同步变动,e-ratio 随着PP的增加而提高[24-28].Dunne等[24]基于卫星数据推算PP并分析了不同海区的EP变化情况:在极地、沿海和环流边界区域,EP易发生显著变化,且在南大洋受海冰的影响,总EP的年内变化率超过30%;在所有极地和亚极地海区,EP的年内变化都比年际变化更显著,只有在赤道太平洋和大西洋以及印度洋的亚热带流涡区,EP才具有较强的年际变化.Laws等[27]通过构建食物网模型,对全球多个典型海洋生态系统中PP、EP和环境变量之间的关系进行了定量分析,发现e-ratio与PP之间存在双曲线关系:当温度高于25 ℃时,e-ratio对PP变化的响应相对不敏感,其范围在0.1~0.2之间;但当温度低于20 ℃时,e-ratio随着PP的增加而增加.当PP的范围在10~100 mg/(m3·d)时,e-ratio对PP变化响应不敏感,并且与温度呈负相关;当PP<10 mg/(m3·d)时,PP与e-ratio之间存在负相关关系.该预测结果也与在北太平洋亚热带流涡区ALOHA站(22°45′ N,158° W)的实测数据一致.随后,Laws等[26]进一步以SST和PP为自变量建立函数关系估算EP和e-ratio,发现:e-ratio与SST呈线性负相关,与PP呈非线性正相关,并且在高纬度海区e-ratio的季节变化最大;近岸上升流区域的e-ratio较大,寡营养流涡区的e-ratio最小;同时,与PP的季节性变化趋势相同,EP一般从冬季到夏季呈上升趋势,高纬度海区的EP较大,寡营养流涡区的EP最低,在秘鲁和本格拉洋流系统等生产力较高的上升流区域,EP通常都较大.
然而,近年来越来越多的研究结果开始挑战以上观点.不管在高纬度还是在低纬度海区,PP与EP的不耦合都有报道,显示两者之间存在明显的时滞效应[23,29-35].特别是在对南大洋的一系列研究中发现e-ratio与PP呈负相关关系[36-40]之后,PP与EP的不耦合成为研究者关注的热点问题.Baumann等[29]在评估2008—2010年白令海东陆架断裂带附近海冰边缘海域e-ratio的季节性模式时,观测到春季冰缘附近PP值较高但EP值较低,导致e-ratio<0.25;而在初夏,各站点的EP>PP,导致e-ratio>1,由此发现了春、夏两季PP与EP存在时间上的解耦.此外,Lepore等[30]和Moran等[31]研究了北极楚科奇海的e-ratio,发现:PP和EP在季节上的时间解耦更加明显,2002年春季的e-ratio介于0.02~0.1之间,夏季的e-ratio介于0.2~0.6之间;同时,所有站位都显示POC通量存在显著的季节和空间差异性,春季的POC通量平均为(2.9±5.3) mmol/(m2·d),夏季增加了2.6倍,平均为(10.5±9.3) mmol/(m2·d).Maiti等[38]在对南大洋的e-ratio进行研究时发现PP与e-ratio之间呈反比关系,这也与此前科学家进行模型预估的结果相背,并发现该海区的e-ratio对温度变化的响应不敏感.此外,EP也有明显的区域差异.例如,Buesseler等[41]在对全球海洋的垂直运输(VERTIGO)项目研究中,通过中性浮力沉积物捕获器测量了颗粒物的下沉通量,发现颗粒通量衰减呈显著区域差异:与亚热带太平洋环流相比,西北太平洋有更高比例的净初级生产力(NPP)到达海洋深层(500 m).
Henson等[22]通过构建全球生物地球化学模型,发现在温暖海区e-ratio随PP的增加而增加,而在寒冷海区则相反.亚热带海区和亚极地海区PP和EP的关系存在明显差异:在亚极地海区(SST<7 ℃),低PP对应着高EP;而在亚热带海区,低PP对应着低EP.实测数据也显示,高纬度海区的e-ratio一般从春季到夏季呈上升趋势[29,42-43],而低纬度海区的e-ratio相对恒定[32,44],但在百慕大大西洋时间序列站(BATS)的研究中,在PP非常低的时期,偶尔也会观测到高e-ratio[33].他们同时发现,高纬度和低纬度海区不同的颗粒沉降速度对e-ratio有明显的影响:在低纬度海区,高比例的缓慢下沉颗粒和低e-ratio之间的联系相当直观,缓慢下沉的小颗粒在海洋上层停留的时间较长,从而可以在相对较浅的深度(<100 m)被迅速再矿化,因此不会产生较大的EP;但在高纬度海区(年均SST<3 ℃),下沉较快的大颗粒会导致较高的EP.总的来说,其研究表明,e-ratio的年际变化确实存在,但变化程度与区域有关.此外,Henson等[22]的模型预测结果与Maiti等[38]和Laws等[27]的观测结果相符,这进一步验证了PP和e-ratio之间的关系存在显著的区域差异性.Henson等[22]还指出在亚极地海区e-ratio与PP的这种负相关关系与EP和PP之间的时滞现象有关.同时,他们估算出全球海区的EP与PP的时滞范围:在上升流以及低纬度海区EP与PP的时滞较短,为0~10 d;北太平洋的时滞为10~15 d;南大洋和北大西洋有20~30 d的时滞;而在营养贫乏的环流边缘和北极,EP与PP的时滞达到30~40 d.在高纬度海区,PP呈现出明显的季节性变化,PP从春季到夏季持续增加,且在夏季会出现一个持续时间较长的单峰,而在这些海域中夏季PP的增加并不会立即被输出,导致EP与PP出现时间解耦,使得e-ratio存在很大的季节变异性.随后,Laws等[45]重新分析了南大洋的历史资料,发现如果假设EP与PP的时滞为25 d,PP与e-ratio之间的负相关关系就转变成正相关关系,再次证实了PP的季节变化及时滞现象对e-ratio的估算存在显著影响,并证明只有在保证EP的积分时间大于EP滞后PP的时间基础上才可能获得e-ratio的有效计算.
2 EP与PP的时滞效应研究进展及存在的问题
2.1 造成时滞差异的原因
PP通过食物网的传递转换为EP,但该过程并非瞬时发生的.在一个生产周期内,当PP迅速增加时,颗粒需要在上层海洋中经过一段时间的处理,达到一定的密度和尺寸后才能转换为EP,因此在一个周期刚开始时e-ratio较低而周期结束时e-ratio较高.现场实测数据也验证了PP和EP之间的时滞与食物网的结构有关[29,32-35].Henson等[22]解释PP与e-ratio呈负相关关系的基本假设就是基于食物网过程的季节性演替:在春季藻华暴发初期,PP迅速增加,但浮游动物的摄食等碳处理过程并没有迅速增加,因而EP仍然较低,导致出现高PP对应较低的e-ratio;在藻华暴发后期,PP已经下降,但由于上层海洋仍残留较多的碳,碳输出可能仍在进行,因而EP仍然较高,导致低PP对应较高的e-ratio.
但对不同海区而言,EP与PP之间的时滞成因可能存在差异.在高纬度海区,其时滞长短可能主要由外部环境因素对食物网结构产生的影响大小来决定.Baumann等[29]对白令海东陆架断裂带附近海冰缘的的研究中,认为海冰消退时间差异导致的PP增加与大型浮游动物出现的时间差变化是决定时滞的重要因素.在春季,太阳辐射的季节性增加以及春季海冰的消退,使得陆架断裂带和白令海东陆架海区营养丰富的海域PP较高[46-48],而同一时期浮游生物生长的变化与PP的变化并不同步[49],初级和次级生产者之间的生长脱钩,导致上层海洋残存大量未经浮游动物处理的浮游植物[50-51];随着温度逐渐回升,夏初浮游动物的摄食活动变强,上层海洋中的残存颗粒也逐步转换为EP.Brown等[48]也发现,在白令海东南部,年NPP在海冰消退较早的年份比海冰消退较晚的年份高40%~50%,验证了海冰消退的时间对初级生产力的大小起着重要的控制作用.
在亚极地和亚热带海区,浮游植物与浮游动物的耦合程度以及浮游动物群落的季节性动态对PP与EP的时滞影响更大[22].在亚极地海区,春季浮游植物生物量迅速增加,而浮游动物的生长相对滞后,因此该阶段只产生缓慢下沉的颗粒,EP与PP之间的滞后很大.随着浮游动物生物量的增加,快速下沉的颗粒也在增加,因而EP随之增加.随着浮游植物和浮游动物的生物量在藻华后期的耦合越来越紧密,EP与PP之间的滞后逐渐缩小.而在亚热带海区,缓慢下沉的颗粒主导了全年的总POC通量.浮游植物和浮游动物生物量的季节变化范围非常小,耦合较紧密,因此EP和PP之间几乎没有滞后.可见,EP和PP之间的滞后主要是由于初级生产者和他们的消费者之间的季节性脱钩,以及随之产生的缓慢沉降和快速沉降颗粒生产的脱钩.浮游动物群落的季节性动态也可能会影响EP与PP之间的时滞.例如,许多桡足类和磷虾类会在深海越冬,并在藻华之前到达表层,从而在POC向下输出的过程中发挥作用.水生动物的换皮、自然死亡和越冬群落的捕食者死亡都能提高POC水平,而这些都与浮游植物暴发的时间无关.
此外,导致高PP低EP的机制还有很多,如营养结构差异、再矿化效率、高细菌活性(异养细菌会将POC酶解为DOC[52-53])以及真光层DOC输出的增加等.例如,le Moigne等[54]将南大洋的低PP、高e-ratio解释为强浮游动物摄食和弱细菌活性的结果:一方面,浮游动物的强摄食行为将浮游植物进行“打包”和“聚集”,加快了颗粒物的沉降速度;另一方面,低细菌活性降低了有机碳在上层的再矿化速率.Henson等[23]进一步整合分析了全球范围内PP与e-ratio的关系,并从浮游生物(浮游植物、浮游动物和细菌)丰度之间的耦合程度解释了PP与e-ratio之间的不耦合关系.其结果显示,造成PP与e-ratio不耦合的原因具有很大的区域差异,其原因很可能与浮游生物的群落组成及食物网的结构有关,但缺乏足够的观测资料支撑,因此造成不同生态系统时滞差异的原因尚不明确.但可以明确的是,预测POC通量随PP增加而增加只是最简单的碳输出模型,而针对不同的海洋生态系统,可能需要附加更多描述食物网结构的参数、调整现有与温度相关参数的权重,来构建适应不同海洋生态系统的PP与EP时滞算法.
2.2 估算PP与EP及二者间时滞的方法
2.2.1 估算PP的方法
目前,实测PP主要是采用14C培养法[55-57],间接估算PP则常用叶绿素含量来估计[58-60].在14C 培养法中,培养开始时以 NaH14CO3的形式将一定量的无机14C添加到已知CO2总含量的海水中,培养时间通常为12~24 h.若假设14CO2仅通过光合作用被浮游植物吸收,而且14CO2与12CO2是以相同的速率被光合作用同化,通过测定培养结束后浮游藻类中14C的含量,即可确定总碳同化量(将测定的14C含量乘以培养开始时海水中CO2总含量和14CO2含量之间的比值),再将总碳同化量除以相应的海水体积及时间,即可推算出PP.
间接估算PP的方法中,Ryther等[58]通过海水中的叶绿素含量、每日到达海面的总太阳辐射和水柱中可见光的消光系数来估计单位海面下的有机物产量.该方法的优点是不需要直接测定光合作用以及其他相关参数,节省了大量的时间,并且对于日常调查工作而言,辐射数据以及消光系数都易获得,过滤后的叶绿素样品也可储存数月.Platt[59]认为PP和到达海面的太阳光之间存在稳定的关系,并提出在确定无量纲参数的偏差后,两者之间是线性相关的.该方法还可以用于通过遥感数据估算大面积海域的PP.Behrenfeld等[60]开发了一个依赖光学深度分辨碳固定的模型(VGPM),该模型将影响PP的环境因素分为影响PP相对垂直分布的环境因素和控制PP最佳同化效率的环境因素,并表明能越精确地描述PP最佳同化效率在空间和时间上的变化,则模型的算法越准确.
2.2.2 估算EP的方法
有机碳的向下输出通量可以利用营养盐的吸收、同位素示踪法、卫星遥感经验算法、水下视频系统等间接方法估算,也可以通过各种沉积物捕获器或海雪捕获器进行直接测量[61].目前,EP的测定最常用的还是同位素示踪法[25,33,61]和中性浮力沉积物捕获器法[13,62].同位素示踪法依赖于附着在颗粒物上的天然同位素的放射性子同位素产物,例如234Th或210Po.随着颗粒物的下沉,母同位素和子同位素之间出现不平衡,进而通过测得的POC与同位素活度比值来估计EP.这些方法具有与子同位素半衰期相关的积分时间尺度,范围为24 d(234Th-238U不平衡法)到135 d(210Po-210Pb不平衡法)不等.中性浮力沉积物捕获器通常与传统停泊式沉积物捕获器的结构类似,但该捕获器可在预定深度自由漂流,将下沉颗粒的样品收集于捕获管中,并分析POC含量.这些类型的沉积物捕获器通常布放时间只有0.5~5 d[22].由此可知,研究中造成e-ratio偏差的一个很重要原因是,EP的积分时间少于EP与PP的滞后时间.因此,准确估算EP与PP的滞后时间,是指导EP的现场测定,尤其是沉积物捕获器现场布放时间的必要前提,对有效估算e-ratio 至关重要.
2.2.3 估算PP与EP时滞的方法
由PP和EP的估算方法可知,PP的积分时间通常为12~24 h,而EP的积分时间则为几天到几个月不等[13,63].如前所述,理论上PP不太可能瞬间转化为沉降 POC.不论是单个颗粒的聚集,还是浮游动物的摄食,以及随后作为粪便颗粒的代谢等过程,颗粒物都必须达到足够的密度或尺寸才能下沉,而这些过程都需要时间[22,45].因此,EP的积分时间很可能少于EP与PP的滞后时间,造成e-ratio的偏差.但在实际研究中,很难通过现场观测数据来量化EP与PP之间的这种时间滞后.一部分原因是实测数据通常没有覆盖整个季节周期(除时间序列分析站如BATS外),另一部分原因是测量EP和PP的时间尺度通常都不一致.
模型模拟是目前估算EP与PP时滞的最有效手段.Henson等[22]提出的生物地球化学模型是目前少有的估算EP与PP时滞的模型.该模型由浮游生物模型MEDUSA(涵盖生态系统动力学、碳利用、固碳及酸化模块)嵌套海洋环流模式 NEMO 组成,但真正决定EP与PP时滞的是生物模型MEDUSA[64].该模型对了解全球尺度EP与PP的时滞分布趋势有重要参考价值.然而,在某些局部区域有些结果却难以解释,如其结果显示缓慢沉降的小颗粒在有些区域可100%沉降,但类似结果在现场观测中从未出现过[22].造成这种偏差的原因很可能在于其生物模型对食物网的描述过于简单.MEDUSA模型模拟了氮、硅和铁的循环,并将浮游生物群落分为“大”和“小”两部分.其中生态系统的“小”部分代表微型浮游植物和小型浮游动物的微生物循环,“大”部分代表小型浮游植物(硅藻)和中型浮游生物.在MEDUSA模型的食物网中最后形成的碎屑也分为小颗粒和大颗粒,其中大颗粒碎屑可迅速沉降;而小颗粒碎屑以缓慢速率(3 m/d)沉降,其矿化速率受温度的调控,即小颗粒碎屑的矿化速率=0.016×1.066t×小颗粒碎屑浓度(其中,温度t的单位为℃;小颗粒碎屑浓度以氮浓度表示,单位为mmol/m3).且其食物网并没有直接考虑对有机碳矿化起重要作用的细菌的影响.因此,估算的时滞完全取决于一个简单的函数,其中温度和小颗粒碎屑浓度是决定输出效率的关键因素.但事实上,从PP到最终转换为EP的过程复杂,其中营养盐吸收、浮游动物摄食、细菌呼吸等很多过程都可能受温度调控[65-67],进而影响 POC的形成、矿化和沉降速率,这种复杂关系是难以通过一个简单函数来表达的.因此,在区域尺度EP与PP的时滞需要采用更加精细的食物网结构模型估算.
与复杂的生物地球化学模型不同,单纯的食物网模型可更聚焦于食物网结构与有机碳输出的关系,是更直接的用于估算EP和e-ratio的方法[27,68-69].例如,与MEDUSA模型相比,Laws等[27]提出的食物网模型(Laws模型)对食物网结构的描述更为全面.其原理是假设食物网行使类似电路中电容器的功能,通过改变稳态模式下的输入值以扰动系统,对稳态方程进行积分,以了解扰动对输出的影响.Laws模型将食物网假设为初级生产按两条食物链传递:其中一条以大粒级浮游植物为基础,其食物链较短;另一条以小粒级浮游植物为基础,其食物链较长,且直接考虑细菌的作用.在以大粒级浮游植物为基础的食物链中,生物体产生碎屑颗粒有机物(POM),而不产生溶解有机物(DOM);在以小粒级浮游植物为基础的食物链中,小粒级浮游植物、鞭毛虫、纤毛虫产生DOM,但不产生碎屑POM.这两条食物链在中型浮游动物处汇合,中型浮游动物由捕食纤毛虫和大粒级浮游植物的滤食动物组成.此外,Laws模型假设所有组分(细菌、浮游植物、鞭毛虫、纤毛虫、滤食性和肉食性浮游动物)的饱和摄食或吸收速率都是温度的函数,因而所有状态变量都受温度的影响,进而影响沉降速率.因此,类似 Laws模型的单纯食物网模型可能理论上更适于估算EP与PP的时滞问题,但目前无人尝试.
以上的模型模拟都主要聚焦于BGP对EP的影响,但Henson等[70]提出颗粒的碎裂和迁移泵也会对EP产生重大影响.他们认为小颗粒的表面积与体积比较大,这会促使大颗粒碎裂成小颗粒进而促进POC的再矿化[71-72].Briggs等[73]利用生物地球化学Argo浮标收集光学数据,同时跟踪中层水中大小颗粒的浓度变化,发现颗粒的碎裂可能会驱动中层水50%的再矿化.此外,他们发现由浮游动物和游泳动物主导的垂直迁移也是EP的重要组成部分;如果将迁移泵包括在内,目前全球计算EP模型的估计值会增加14%~40%[14-15,74],在特定地点可能增加更多[75].由于颗粒碎裂和迁移泵的驱动因素目前尚不确定,许多模型都没有将其考虑进去或缺乏准确描述,但可以确定的是,这些因素会对EP产生显著的影响.尤其是随着气候变化,这些过程对温度的依赖性导致其强度发生的改变,都会是未来研究需要突破和关注的问题.
3 南海EP与PP时滞效应研究展望
南海是西太平洋最大的边缘海,本身环境梯度变化大且具有高度的空间异质性,其营养水平和浮游植物生物量从陆架到海盆呈递减趋势,且陆架区季节变化大于海盆区[76-78],这种变化趋势与全球尺度上高纬度到低纬度的变化模式非常相似.过去10多年,南海生物泵研究基本上与全球其他海域同时期开展,并取得较好的进展,如浮游植物群落结构和PP等自养过程参数[55,78-80],浮游动物摄食、群落呼吸、细菌呼吸等异养过程参数[65,81-83],以及EP或e-ratio等碳输出参数[84-88].南海由于地理优势独特且历史资料丰厚,是进一步研究EP与PP时滞效应的理想试验场.
由于营养水平和生物量的显著差异,南海陆架区PP的年平均值和季节变化都显著大于海盆区[89].根据Henson等[22]的假设,若EP与PP时滞较强,PP的强季节变化会导致PP与e-ratio的不耦合,因此南海陆架区比海盆区更有可能存在PP与e-ratio不耦合的现象.然而南海北部实测数据显示,e-ratio在陆架区为0.34,在海盆区为0.24,且陆架区的e-ratio与PP基本呈正相关,而海盆区的e-ratio与PP则呈负相关[90],这说明南海陆架区EP与PP的时滞可能较弱,而海盆区反而可能存在较强的时滞.Laws等[27]认为食物网以类似于电路中电容器的功能通过响应输入电压变化的方式响应PP的变化.在食物网中,EP与PP的滞后时间是食物网动力学的函数,食物网动力学与温度有关.由此可见,南海浮游生物的食物网结构对EP与PP时滞的调控作用可能大于PP(自养过程)本身的变化,导致观测到的南海海盆区EP与PP的时滞强于陆架区.南海陆架区与海盆区的浮游植物群落关键组成存在明显差异,陆架区以大粒级的硅藻为主,而海盆区以粒级较小的聚球藻(Synechococcus)、原绿球藻(Prochlorococcus)、定鞭藻(Haptophytes)和青绿藻(Prasinophytes)为主[78].小型浮游动物是浮游植物的主要摄食者,并可以快速响应浮游植物增长率的增加[91].此前Calbet等[92]在比较全球数据后,发现浮游动物的摄食率在贫营养盐的水域(70%)高于河口和近岸水域(60%),这可能是由于在营养丰富的水域浮游植物的粒径更大,小型浮游动物难以摄食.Chen等[93]利用全球表层水的14C吸收以及稀释实验的数据验证了浮游动物的摄食率与浮游植物平均大小呈显著负相关.然而,在南海夏季的观测数据中,从陆架区到海盆区,小型浮游动物对浮游植物的摄食率呈下降趋势,且在海盆区小型浮游动物的摄食对浮游植物的影响较小,这表明小型浮游动物摄食与浮游植物生长不耦合[81];Chen等[82]比较了南海北部不同地区和不同季节中型浮游动物对浮游植物的摄食率和每日消耗量,发现中型浮游动物在南海北部陆架区对浮游植物有更高的摄食量,因而陆架区中型浮游动物的摄食压力也比海盆区大.本课题组也有研究表明海盆区单位PP的细菌呼吸比陆架区高,即海盆区细菌再矿化对POC衰减的影响较大(未发表数据).
基于以上分析,可以结合Laws模型,进一步研究南海EP与PP时滞的时空分布模式以及浮游食物网的结构如何决定不同生态系统的时滞差异,并提出如下科学假设(图1):在南海陆架区,大粒级浮游植物比例较高,PP较大,碳流过程以短食物链为主,中型浮游动物对浮游植物的直接摄食作用较强,其主导的异养过程可迅速响应初级生产过程;加上细菌再矿化对POC衰减的影响较弱,因而颗粒沉降较快,EP与PP较同步变化,时滞较弱.相对地,在南海海盆区小粒级浮游植物比例较高,PP较小,碳流过程以长食物链为主,中型浮游动物对浮游植物的直接摄食作用较弱,导致其对小型浮游动物的摄食不能迅速响应初级生产过程;加上细菌再矿化对 POC 衰减的影响较强,因而颗粒沉降较慢,EP与PP的变化不同步,时滞较强.
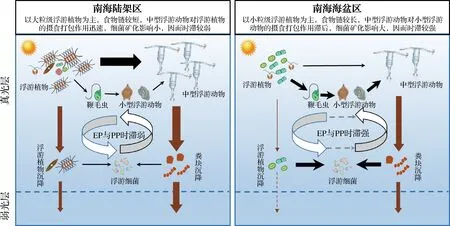
图1 简化的南海不同生态系统真光层生物泵输出示意图Fig.1 Simplified schematic diagram of the export of biological pumps in the euphotic zone of different ecosystems in the South China Sea
随着全球气候的逐渐变化,有研究预测随着未来海洋的混合层深度变浅以及海洋环流减缓,海洋表层向下输送的营养物质会逐渐减少,从而导致海洋生物碳泵的强度逐渐下降,海洋碳储量减少,这意味着大气中的CO2水平将逐渐增加[94-96].而南海作为西太平洋的一个大型半封闭边缘海,其海盆深达5 000 m,周围环绕着广阔的大陆架.虽然南海近岸和陆架受珠江、湄公河等河流输入的影响,但是海盆区通常为全年贫营养,这一特点使南海成为研究显著环境梯度下生物泵碳封存的理想场所[97].因此,研究南海EP和PP的时滞,有助于进一步揭示未来全球气候变化对海洋造成的影响以及海洋生物碳泵的强度变化.
猜你喜欢
海洋通报(2022年2期)2022-06-30
石油地球物理勘探(2022年3期)2022-06-11
海洋信息技术与应用(2021年3期)2022-01-17
石油钻探技术(2020年3期)2020-08-24
海洋学报(2020年7期)2020-08-06
建筑热能通风空调(2018年5期)2018-07-09
海洋信息技术与应用(2017年2期)2017-06-21
海洋渔业(2016年6期)2016-04-16
中国工程咨询(2016年9期)2016-02-13
应用海洋学学报(2014年1期)2014-11-22
