
敬信湿地弃耕稻田恢复演替年限对植物群落多样性的影响
2023-10-07 01:49汤钰琦尚钇君朱卫红曹光兰
生态与农村环境学报 2023年9期
汤钰琦,尚钇君,朱卫红,曹光兰,3①
(1.延边大学地理与海洋科学学院,吉林 珲春 133300;2.长白山湿地生态系统功能与生态安全吉林省重点实验室,吉林 延吉 133002;3.南开大学环境科学与工程学院,天津 300350)
湿地生态系统因具有维持生态系统平衡、调节气候、涵养水源等重要作用,是全球重要生态系统之一[1-3]。与其他生态系统类型相比,湿地生态系统具有独特的植物群落结构特征和丰富的生物多样性,是动植物生存的重要场所[4-5]。湿地植物作为湿地初级生产者,通过与湿地其他生物因素以及湿地土壤、水位等非生物因素间相互作用,对湿地功能产生重要影响[6]。随着社会经济的发展,湿地及其生物多样性遭受破坏的问题愈加严重,湿地恢复对区域生态、经济和社会可持续发展具有重要影响,因此湿地的保护和恢复被认为是全球重要的环境热点问题[7]。
稻田属于人工湿地,弃耕稻田保留了湿地原有植物种子,作为湿地植物潜在的再生地,弃耕稻田的自然恢复被认为为湿地历史植被恢复的机会。随着弃耕年限的增加,弃耕稻田可逐渐变成具有独特植物群落结构特征和较高生物多样性的天然静水湿地[5,8]。植被特征不仅是评估湿地恢复进展最简单和最常用的指标,也是维持湿地生态系统健康稳定的关键因素[9-11]。湿地植被恢复直接反映在植物群落组成和植物多样性变化上,因此植物群落组成和多样性被广泛地用于生态恢复评价[12-14]。弃耕稻田对生物多样性的影响比农业扩张所造成的持续负面影响更加复杂,弃耕稻田也被认为是全球生物多样性变化的关键驱动因素[15]。长期耕种导致湿地种子库有所损失,导致植物多样性显著下降,因此了解再生湿地自然恢复过程中植物群落的发展非常重要[5]。
吉林省是全国湿地类型较多的省份之一,但由于水资源短缺,许多湿地几乎干涸,生态环境持续恶化[16]。敬信湿地是图们江流域生态环境的重要组成部分,是东亚-澳大利亚候鸟迁徙带的重要中转站[17]。受国家政策调整、气候变化、人口增加、农业生产和水利工程建设等多个因素的综合影响,近年来敬信湿地面积呈现先减少后增加趋势,并呈现明显的湿地景观破碎化趋势,生物多样性严重下降,生态系统稳定性受到极大威胁[18-20]。随着经济发展和生态环境保护意识的提高,图们江下游敬信湿地的稻田在经过长期农业耕作之后逐渐被废弃,敬信湿地得到不同程度的恢复。对图们江下游敬信湿地地区不同弃耕年限稻田湿地的植物群落物种组成以及α和β多样性进行研究,明确敬信湿地不同弃耕年限湿地植物群落结构特点,揭示弃耕稻田湿地生态恢复过程,从而为敬信湿地弃耕稻田的管理以及湿地恢复与重建提供理论支持,也为其他同纬度湿地恢复提供理论依据。
1 材料与方法
1.1 研究区概况
敬信湿地位于吉林省延边朝鲜族自治州珲春市南部的第2大平原区,地处中、朝、俄三国交界地带,属于图们江水系下游,地理位置为42°27′ N~42°40′ N,130°25′ E~130°39′ E,海拔高度为5~15 m(图1)。由于距日本海较近,受海洋性气候影响,与同纬度其他地区相比,该地具有冬暖夏凉、春秋季风大、多阴寡照和降水充沛的气候特点,属于中温带近海洋性气候区,年降水量为600~800 mm,年平均气温为5.6 ℃,属于湿润区[21-23]。敬信湿地内河道蜿蜒曲折,湖泊(泡)棋布,具有河流型、湖泊型和沼泽型等多种湿地类型,是吉林省唯一近海湿地[24]。敬信湿地生物多样性丰富,湿地生产力高,分布在该地区的植物有59目119科314属537种。近年来,由于农业竞争力逐渐减弱,人口由农村向城市流动,延边州内朝鲜族人口向韩国等地流动,农业劳动力减少,使得敬信湿地内大部分水田被废弃[20,25]。
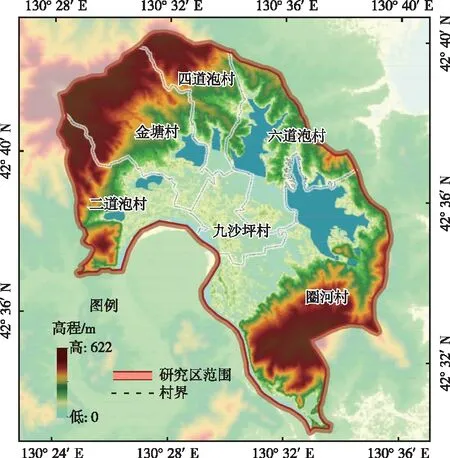
图1 研究区概况
1.2 试验设计
于2017年7月开展湿地植物群落野外调查工作,采用空间序列代替时间序列的方法对敬信湿地进行不同弃耕年限的划分,选取弃耕年限<5 a的湿地,设置7个样地;弃耕年限5~15 a的湿地,设置8个样地;弃耕年限>15 a的湿地,设置7个样地;同时选取自然湿地作为对照,设置11个样地。稻田弃耕年限信息通过对土地所有者和研究区当地村长走访调查得到。由于未能找到完全未受人为活动干扰的湿地,选取一块近50 a未被开垦的湿地作为自然湿地。为了能全面收集湿地植被信息,采用地块法和样方法[26-28]对植被群落进行调查。每个样地设5个样方,每个样方大小为1 m×1 m,为了避免样地之间的空间自相关,样地间隔至少为1 000 m,且同一样地各样方的植物景观保持一致。记录各湿地类型、样方内植物物种组成和各物种盖度等相关信息,并用GPS仪记录样方经纬度等位置信息。湿地样地信息见表1。
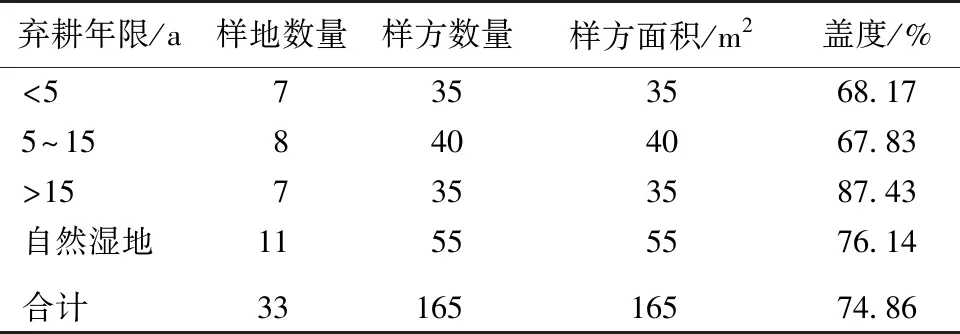
表1 敬信湿地样地信息
1.3 生物多样性的测度
根据样地调查数据,分别计算不同弃耕年限湿地α多样性指数和β多样性指数。其中,α多样性指数选取Patrick物种丰富度指数(R)、Shannon-Wiener多样性指数(H)、Simpson优势度指数(D)和Pielou均匀度指数(J)进行测度,各指数计算公式[29]为
R=S,
(1)
(2)
(3)
J=H/lnS,
(4)
Pi=S/N。
(5)
式(1)~(5)中,S为物种个体数;N为样地各物种个体总数;Pi为物种i个体数占所有个体总数的比例。H值越大,群落越复杂,植物多样性越高;D值越大,物种数越多,群落整体组成越均匀;J反映群落中不同物种多度分布的均匀度[30]。
β多样性采用Jaccard成对相异系数进行测度。参照BASELGA[31]的加和分解法,将Jaccard成对相异系数(βjac)分解为物种更替组分(βjtu)和嵌套组分(βjne),其计算公式为
(6)

(7)
(8)
式(6)~(8)中,a为群落A和B共有物种数,b和c分别为群落A和B的物种数。
1.4 统计方法
采用R 4.1.2软件计算α多样性指数。采用SPSS 26软件中方差同质性(homogeneity of variance)检验进行方差齐性检验(P>0.05)后,再用单因素方差分析(one-way Anova)和Duncan多重检验(P<0.05)比较不同弃耕年限湿地α多样性以及α多样性成分间的差异。采用最小显著差异法(LSD)对不同弃耕年限和自然湿地采集的数据进行差异显著性检验。采用R 4.1.2软件betapart包(http:∥CRAN.R-project/org/package=betapart)计算β多样性。采用R 4.1.2软件ggplot2包和Excel 2017软件绘图。
2 结果与分析
2.1 湿地植物群落物种组成
敬信湿地33个样地植物种类野外调查结果表明,共有植物99种,均为草本植物,隶属于33科64属。其中,弃耕年限<5 a稻田有61种植物,隶属于25科45属;弃耕年限5~15 a稻田有48种植物,隶属于20科34属;弃耕年限>15 a稻田有41种植物,隶属于20科27属;自然湿地有36种植物,隶属于15科24属。随着弃耕年限的增加,植物群落科、属、种数量均呈现下降趋势。不同弃耕年限稻田优势种见表2。
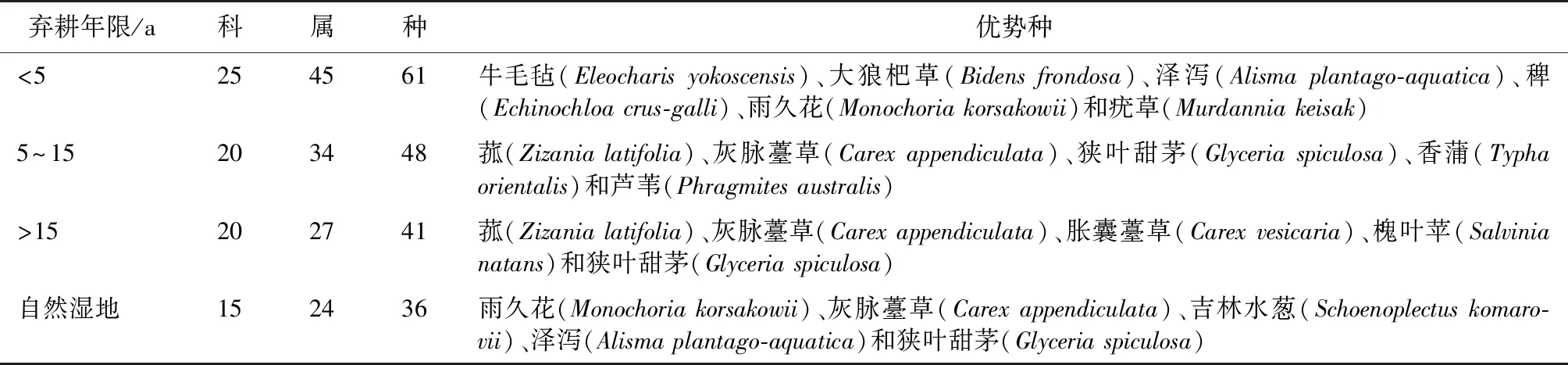
表2 敬信湿地不同弃耕年限稻田主要植物物种构成
调查结果显示,敬信湿地植物群落中莎草科(Cyperaceae)有16种,占总植物种数的16.2%;蓼科(Polygonaceae)有15种,占总植物种数的15.2%;禾本科(Poaceae)有12种,占总植物种数的12.1%;菊科(Asteraceae)有9种,占总植物种数的9.1%;唇形科(Lamiaceae)有6种,占总植物种数的6.1%。此外,千屈菜科(Lythraceae)、伞形科(Apiaceae)和十字花科(Brassicaceae)各有3种植物;车前科(Plantaginaceae)、豆科(Fabaceae)、木贼科(Equisetaceae)、通泉草科(Mazaceae)、雨久花科(Pontederiaceae)、泽泻科(Alismataceae)和香蒲科(Typhaceae)各有2种植物,上述植物共计21种,占总植物种数的21.2%。敬信湿地植物群落单科植物有18种,占调查植物种数的18.2%,单属植物有50种,占调查植物种数的50.5%,这表明敬信湿地植物仍具有多样性。
不同弃耕年限稻田草本植物数量见表3。弃耕年限>15 a稻田植物群落有41种草本植物,在3种弃耕年限稻田中最低,与对照自然湿地植物群落的36种草本植物最为接近。3种弃耕年限稻田与自然湿地植物群落均以湿地植物为主,占比分别为72%、83%、76%和83%,并且湿地植物以多年生植物为主。从功能群组成来看,弃耕初期,以一年生草本植物为先锋植物,随着弃耕年限增加,一年生草本植物地位逐渐下降,多年生草本植物在群落中的优势地位逐渐上升,逐渐由一年生草本植物群落向多年生草本植物群落方向演替,最终形成以雨久花(Monochoriakorsakowii)、灰脉薹草(Carexappendiculata)、吉林水葱(Schoenoplectuskomarovii)、泽泻(Alismaplantago-aquatica)和狭叶甜茅(Glyceriaspiculosa)等多年生湿生草本为优势种的植物群落。

表3 基于生活型的不同弃耕年限稻田草本植物数量
2.2 不同弃耕年限稻田植物群落α多样性分析
从敬信湿地不同弃耕年限稻田植物群落物种多样性(图2)可以看出,弃耕年限<5、5~15和>15 a稻田以及自然湿地植物群落物种多样性各指数变化规律较一致。随弃耕年限增加,植物群落Patrick物种丰富度指数(R)在9.10~21.71之间;Shannon-Wiener多样性指数(H)在0.08~2.65之间;Simpson优势度指数(D)和Pielou均匀度指数(J)随弃耕年限增加,在0~1.00之间变化,变化范围相对较小。随着弃耕年限增加,植物群落物种丰富度逐渐下降,植物群落物种数量逐渐变少,植物群落趋于单一化。除Patrick物种丰富度指数外,不同类型湿地植物Shannon-Wiener多样性指数、Simpson指数和Pielou均匀度指数均呈先增加后减少再增加的变化趋势,且均在弃耕年限5~15 a稻田达到最大,在弃耕年限>15 a稻田达到最小。整体来看,随着弃耕年限增加,弃耕稻田植被逐渐自然恢复,弃耕稻田植物群落α多样性指数呈现下降趋势,并最终接近自然湿地。

箱图上下横线分别表示最大值和最小值,方框表示50%变异区间,方框中横线表示对应指标的中值。英文小写字母不同表示不同类型湿地之间某指标差异显著(P<0.05)。
2.3 不同弃耕年限稻田植物群落β多样性特征
将总体β多样性(βjac)分解成物种更替组分(βjtu)和嵌套组分(βjne)。其中,更替组分是不同弃耕年限稻田之间植物群落β多样性存在差异的主导因素,表明β多样性主要是由湿地植物物种在空间上的相互替换形成,与总体β多样性变化趋势一致;而嵌套组分作用相对较小。图3显示,弃耕年限<5 a稻田植物群落总体β多样性显著高于弃耕年限5~15和>15 a稻田植物群落。随着弃耕年限增加,不同弃耕年限稻田β多样性差异逐渐减小。值得注意的是,弃耕年限5~15与>15 a稻田植物群落β多样性中的更替组分相同。

βjtu为更替组分,βjne为嵌套组分,βjac为总体β多样性。
3 讨论
笔者在敬信湿地调查了不同弃耕年限稻田和自然湿地,共发现湿地植物99种,隶属于33科64属,均为草本植物。同时,不同弃耕年限稻田植物群落优势物种具有一定差异。虽然植物在弃耕年限少(弃耕年限<5 a)时生长旺盛,但大多数为一年生或多年生杂草,如牛毛毡、大狼杷草和稗,此时湿地植物群落优势种并未出现。这是由于对于莎草科等湿地植物种子来说,弃耕稻田土壤水分较少,不能满足其萌发的必要条件[32]。在演替中期(弃耕年限5~15 a),中生植物和湿地植物成为优势种。随弃耕年限增加,植物种类逐渐减少,湿地植物群落优势种接近自然湿地,湿地植物处于植物群落的主导地位。这与次生演替理论一致,湿地被破坏后,植物群落可以通过明显的植被演替过程自然恢复[33]。在弃耕初期,由于稻田耕作措施形成的开放且多变的环境更易于一年生或多年生杂草种子入侵,但随着弃耕年限增加,一年生或多年生杂草群落由于过度消耗土壤表层养分而逐渐消亡,同时,长时间的稻田耕种可能会导致主要湿地物种从种子库中消失,这可能导致弃耕初期稻田物种多样性高于自然湿地,而弃耕年限长(弃耕年限>15 a)时物种多样性小于自然湿地[34-35]。
α多样性反映物种分布数量和均匀性、物种或生物性状的重要性以及群落内物种相似性[36]。笔者研究中随着弃耕年限增加,稻田植物α多样性越来越接近于自然湿地。同时,研究区不同弃耕年限稻田植物群落α多样性变化趋势与Patrick物种丰富度指数不完全一致,但总体变化趋势一致,均在弃耕年限<5 a稻田植物群落达到最大值,这有可能是因为当稻田被废弃后,稻田土壤水分减少,为中生植物生长提供了条件,进而提高了生物多样性[37-38]。Shannon-Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数均呈现先增加后减小趋势,这可能是由于弃耕初期稻田残留的耕种环境为一年生草本植物生长提供了条件,但随着弃耕年限增加,一年生植物达到承载上限后,开始出现竞争关系,逐渐被多年生植物所替代,这与王佳等[39]对青土湖不同年限退耕地植被物种多样性的研究结果相似。
β多样性反映群落在物种组成、进化关系和功能属性等方面的空间和时间尺度变异性[36]。物种嵌套组分(βjne)可反映群落构建过程中由于物种丢失或者获得而引起的物种组成变化,而物种更替组分(βjtu)可反映由于物种更替而引起的物种组成变化[40-41]。在笔者研究中,不同弃耕年限稻田植物群落β多样性中更替组分占比近90%,其中,以弃耕年限<5 a稻田植物群落为最大,这种β多样性组成表明环境梯度是稻田弃耕后湿地群落演替的主要驱动因素[42-43]。同时,这也说明在湿地更替的不同阶段会有不同湿地植物出现,更替时间越长,湿地植物种类就越多。
综上所述,由研究区植物物种、不同弃耕年限稻田群落多样性特征可知,研究区具有较多种类湿地草本植物,并以湿生和水生植物为主。研究区不同弃耕年限稻田与自然湿地之间植物群落特征具有显著差异,且随着弃耕年限增加,弃耕稻田与自然湿地之间植物群落构成、α多样性和β多样性差异逐渐减小,弃耕稻田植物群落特征逐渐接近自然湿地水平。这与对同纬度地区日本和韩国的废弃稻田研究结果不同,有关日本和韩国废弃稻田的研究认为以朝鲜柳群落为主的湿地是弃耕稻田演替的最终形态[44-47],而中国黑龙江三江国家级自然保护区湿地经过5年的自然恢复,退耕农田与自然湿地之间植物群落多样性无显著差异[48]。与这些研究区域相比,敬信湿地植物演替速率较慢,弃耕年限<5和5~15 a稻田植物群落的恢复状态尚未达到理想状态,需要更长时间才能达到自然湿地水平,建议开展湿地环境因素驱动下的植被生态演替特征研究。由于缺少弃耕稻田具体弃耕年限数据,笔者在进行弃耕年限划分时受到限制,比较粗略。稻田弃耕对湿地植物群落具有积极影响,但随着植被生长,其对速效养分的需求不断增加,因此建议在弃耕稻田管理中注意补充土壤水分和有机质,并持续加强管理和生态环境保护。
4 结论
在吉林珲春敬信湿地自然恢复过程中的弃耕稻田调查到33科64属99种植物。不同弃耕年限的稻田湿地植物生态特征存在差异,随着弃耕年限的增加,植物群落由一年生或多年生杂草占优势逐渐演变为湿地植物占优势,α多样性指数逐渐接近自然湿地,呈现符合次生演替理论的演替规律。敬信湿地植物演替速率较慢,弃耕<5、5~15 a的稻田植物群落恢复状态尚未达到自然状态。同时,β多样性中更替组分占比近90%,表明环境梯度是弃耕稻田植物恢复过程中群落演替的主要驱动因素。
猜你喜欢
现代园艺(2021年23期)2021-12-01
趣味(作文与阅读)(2021年5期)2021-08-19
现代畜牧科技(2021年4期)2021-07-21
林业勘查设计(2020年1期)2021-01-18
新农业(2020年18期)2021-01-07
作文大王·低年级(2019年2期)2019-01-23
创新作文(小学版)(2018年19期)2018-11-30
启蒙(3-7岁)(2018年8期)2018-08-13
中成药(2017年6期)2017-06-13
动物营养学报(2015年3期)2016-01-07
