
天山北坡蒿类荒漠植物群落数量特征及其多样性
2023-10-11 02:44周时杰董乙强阿斯太肯居力海提聂婷婷姜安静安沙舟
新疆农业科学 2023年9期
周时杰,董乙强,2,阿斯太肯·居力海提,聂婷婷,姜安静,安沙舟,2
(1.新疆农业大学草业学院,乌鲁木齐 830052;2.新疆草地资源与生态自治区重点实验室/西部干旱区草地资源与生态教育部重点实验室,乌鲁木齐 830052)
0 引 言
【研究意义】生物多样性在物种水平上的表现形式是物种多样性,体现了生物之间及其与环境之间复杂关系及生物资源丰富性[1-2]。物种多样性与空间尺度的关系是植物生态学研究热点之一[3],而不同尺度下甚至是不同地区的植物种群分布格局也呈现出差异,种群内部的空间关联也对尺度具有依赖性[4]。因此,开展不同区域植物群落数量特征及其多样性的研究,对提高物种多样性的认识及推进生物多样性保护有重要意义。【前人研究进展】何志斌等[3]研究表明,荒漠植被物种多样性随空间尺度的增大,空间依赖性减弱,与草原和森林群落随尺度增加而增加的结论是相反的。鲁为华等[5]研究结果表明,重度退化区域的绢蒿种群总体上偏离随机分布呈聚集分布,聚集强度在不同尺度上有所变化,在一系列空间尺度上出现明显的“聚集波”现象。陈毅萍等[6]研究表明,在水平方向上草地植物群落多样性从中心到边缘表现出“中心高边缘低”的变化趋势。天山北坡荒漠带处于亚欧大陆腹地,地势多样且物种丰富,蒿类荒漠在天山北坡海拔800~1 600 m的低山区和冲积-洪积扇广泛分布,形成一条脆弱的荒漠带,为春秋牧场,占全疆春秋牧场总面积的28%,并以菊科绢蒿属植物为其主要种群[7]。此外,蒿类植物是荒漠和半荒漠地区羊和骆驼的主要饲用植物。天山北坡蒿类荒漠相关研究,包括短期禁牧对天山北坡蒿类荒漠群落特征及其稳定性的影响[9]、蒿类荒漠草地退化演替的特征[10]、蒿类荒漠草地土壤变化与植被退化的关系[11]、天山北坡围栏封育条件下伊犁绢蒿幼苗分布格局及数量动态变化的规律[12]等。【本研究切入点】空间上,对于天山北坡蒿类荒漠分布规律的系统研究鲜见报道。需开展天山北坡蒿类荒漠植物群落数量特征及多样性研究。【拟解决的关键问题】选取天山北坡西段、中段和东段蒿类荒漠植物群落为对象,研究荒漠草地植物群落数量特征及多样性,分析天山北坡不同区域蒿类荒漠群落特征的空间差异,为新疆荒漠草地的合理利用和科学经营提供数据支撑。
1 材料与方法
1.1 材 料
依据许鹏等[13]文献,将天山北坡荒漠区分为西段、中段和东段。其中,西段是指沙湾市巴音沟以西的北天山山脊线以北的荒漠区,此区域包括新源县、巩留县和博乐市等县(市);中段是指沙湾市以东到乌鲁木齐市的荒漠区,此区域包括玛纳斯县、呼图壁县和乌鲁木齐市等县(市);东段是指奇台县以东的北天山山脊线以北的荒漠区,此区域包括奇台县、木垒县和巴里坤县等县(市)。
研究区地处欧亚大陆腹地,属典型的大陆性干旱气候,夏季炎热干燥,冬季寒冷,年均温3~10℃,年均降水量55~300 mm,荒漠生态系统极其脆弱。建群种主要为伊犁绢蒿(Seriphidiumtransiliense)和博洛塔绢蒿(S.borotalalense),伴生种有木地肤(Kochiaprostrata),羊茅(Festucaovina)、新疆针茅(Stipasareptana),短柱苔草(Carexturkestanica)以及角果藜(Ceratocarpusarenarius)、猪毛菜(Salsolacollina)等,春季有大量早春短命、类短命植物出现,如鳞茎早熟禾(Poabulbosa)、鸢尾蒜(Ixioliriontataricum)、弯果胡卢巴(Trigonellaarcuata)、伊犁郁金香(Tulipailiensis)等。表1
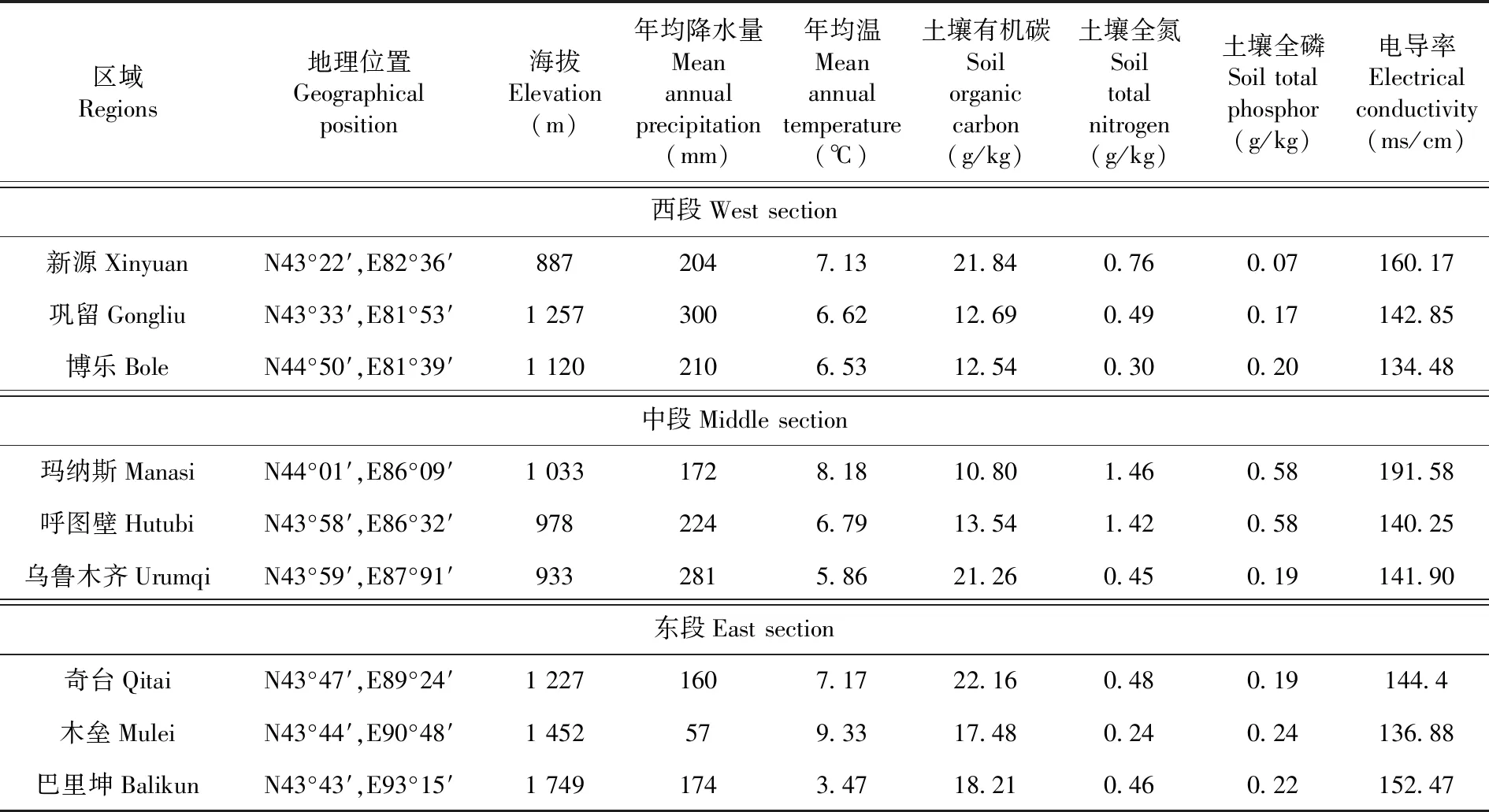
表1 研究区概况
1.2 方 法
1.2.1 试验设计
2021年6月中旬进行野外植被调查与取样,9个研究区分别布设3条样带,样带间距大于50 m,于每条样带上随机设置5个1 m×1 m的样方,样方间距10 m,样方数量共计135个(9个区域×3条样带×5个样方)。记录样方中出现的所有物种种类,用卷尺量取每种植物的自然高度(cm);针刺法测定物种盖度(%);计数法记录每个物种的株丛数(密度,株/m2);齐地面刈割法测定生物量(g/m2)。将植物样品带回实验室置于105℃烘箱杀青30 min,于80℃烘至恒重(24 h)后称重。表1
土壤样品通过5点法用土钻采集0~20cm的土样,常温风干保存,用于土壤养分测定。土壤养分测定参照鲁如坤[14]方法,测定指标包括:土壤电导率、有机碳、全氮、全磷含量。
1.2.2 测定指标
植物群落多样性用α多样性来表征,分别采用Patrick丰富度指数、Pielou均匀度指数、Shannon-Wiener多样性指数和Simpson优势度指数来表示。
重要值(Pi)=(相对密度+相对盖度+相对高度+相对生物量)/4.
Patrick指数(R):R=S.
Shannon-Wiener指数(H):
H=-∑PilnPi.
Pielou均匀度指数(E):
E=(-∑PilnPi)/lnS.
Simpson优势度指数(D):
D=1-∑Pi2.
式中,S为样地总物种数量;Pi为第i个物种的重要值。
1.3 数据处理
采用Excel 2010软件对原始数据进行整理,利用SPSS20.0统计软件对群落特征和多样性指数进行差异性分析,用Pearson相关系数对群落特征、多样性指数等进行相关性分析,运用利用Origin 2018绘图软件进行图表绘制。数据均以均值±标准误的形式表示。
2 结果与分析
2.1 天山北坡蒿类荒漠植物经济类群数量特征变异
研究表明,从天山北坡西段到东段,半灌木盖度和生物量呈逐渐增加趋势,且东段比西段显著高了49.93%和266.51%(P<0.05),而中段的半灌木高度显著高于西段和东段(P<0.05),半灌木的密度虽有一定的增降变化,但差异不显著(P>0.05)。从西到东多年生草本盖度、生物量和重要值呈逐渐降低趋势,且西段比东段显著高了285.58%、270.65%和157.14%(P<0.05),而中段的多年生草本高度显著高于西段和东段(P<0.05),密度虽有一定的增降变化,但差异不显著(P>0.05)。从西到东一年生草本和短生、类短生草本的数量特征均呈先增加后降低趋势,即中段最大,其中除了短生、类短生草本的盖度和生物量差异不显著(P>0.05),其他指标均呈现中段显著高于东段。表2

表2 蒿类荒漠植物经济类群数量特征的变异规律
2.2 天山北坡蒿类荒漠植物数量特征变异
研究表明,天山北坡中段的群落高度、盖度和密度显著高于西段和东段(P<0.05),且天山北坡中段和东段的群落生物量显著高于西段(P<0.05)。图1

图1 蒿类荒漠植物数量特征变异规律
2.3 天山北坡蒿类荒漠植物多样性变异规律
研究表明,空间上Patrick丰富度指数、Shannon-Wiener物种多样性指数和Pielou均匀度指数均具有较大的差异性。西段、中段、东段的Patrick丰富度指数差异显著(P<0.05),数值上中段>西段>东段。Simpson优势度指数方面,西段、中段、东段均无显著差异(P>0.05),数值上也较为相近。天山北坡西段和中段的Shannon-Wiener物种多样性指数显著高于东段(P<0.05),西段的Pielou均匀度指数显著高于中段和东段(P<0.05)。图2
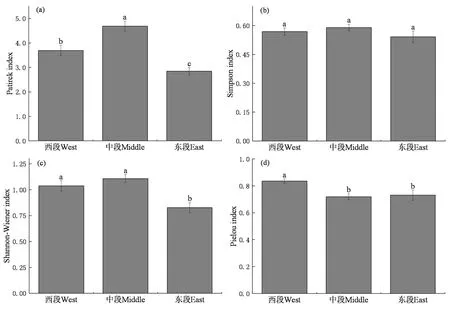
图2 蒿类荒漠植物多样性变异规律
2.4 天山北坡蒿类荒漠植物群落数量特征与多样性之间的相关关系
研究表明,天山北坡不同区域草地之间在西段部分,植物群落高度与植物Simpson指数(-0.306*)、Shannon-Wiener指数(-0.314*)呈现显著负相关(P<0.05);植物群落盖度与Patrick指数(0.517**)、Shannon-Wiener指数(0.455**)呈现极显著正相关(P<0.01),随着植物群落盖度的增加,植物多样性均随之增加,有利于植物生长发育。在中段部分,植物群落高度与Patrick指数(-0.460**)、Shannon-Wiener指数(-0.456**)、Pielou指数(-0.573**)呈现极显著负相关(P<0.01)。在东段部分,植物群落高度与Patrick指数(-0.680**)、Shannon-Wiener指数(-0.657**)、Pielou指数(-0.740**)呈现极显著负相关(P<0.01);植物群落密度与Patrick指数(0.509**)、Shannon-Wiener指数(0.543**)呈现极显著正相关(P<0.01)与Pielou指数(0.353*)呈现显著正相关(P<0.05);植物群落盖度和生物量分别于Pielou指数(0.339*)、Patrick指数(0.303*)呈现显著正相关(P<0.05)。植物群落盖度、密度和生物量对植物多样性呈正相关,而植物高度对植物多样性呈负相关,随着盖度、密度和生物量的不断增加,植物多样性随之增加,而随着高度的增加,植物多样性可能会随之降低。表3
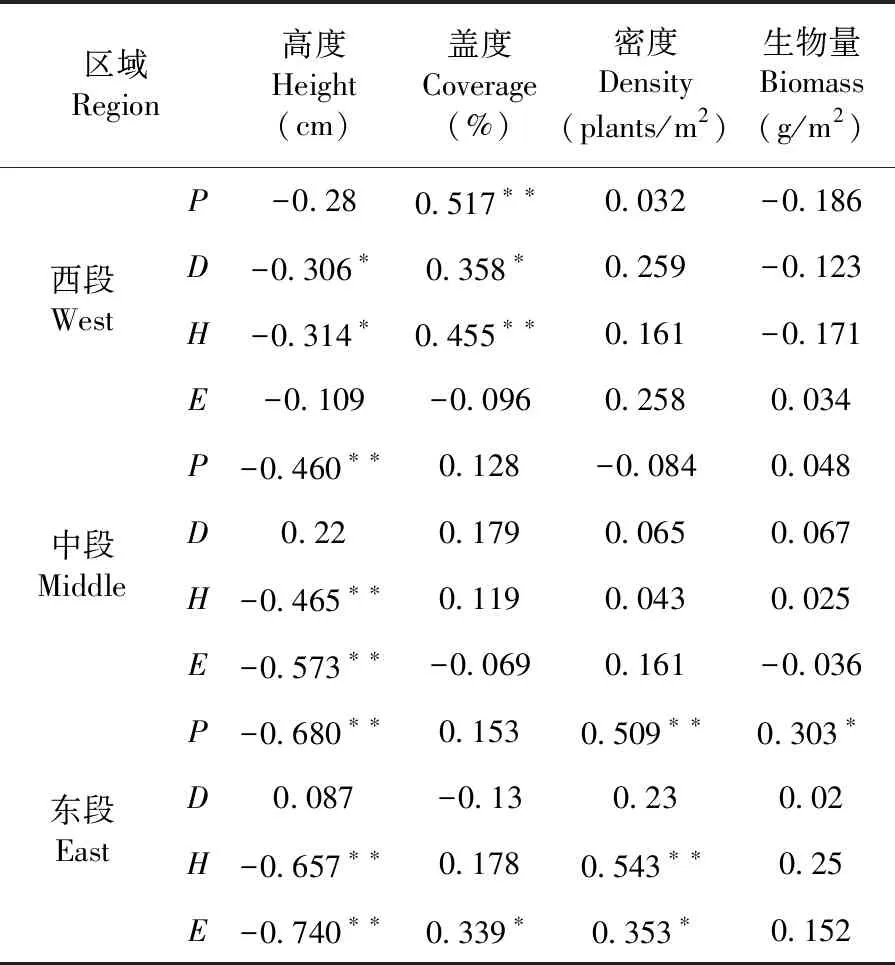
表3 蒿类荒漠植物群落数量特征与多样性之间的相关关系
2.5 天山北坡蒿类荒漠植物群落与环境因子RDA分析
研究表明,轴1与轴2样点环境关系方差的累计解释量达到84.99%,植物生物量与土壤全氮、土壤全磷、经度均呈正相关关系,与土壤有机碳、土壤电导率、年均温度、 年均降水量呈负相关关系。单个环境因子对植物群落的解释能力依次为海拔(38.1%)>土壤有机碳(13%)>年均温度(12.6%)>经度(12.4%)>土壤全氮(9.8%)>土壤全磷(7.5)>年均降水量(3.6%)>土壤电导率(2.8%)。引起植物群落空间变异的主要影响因素是海拔、土壤有机碳、年均温度和经度,其他环境因子对植物群落的影响相对较弱。图3
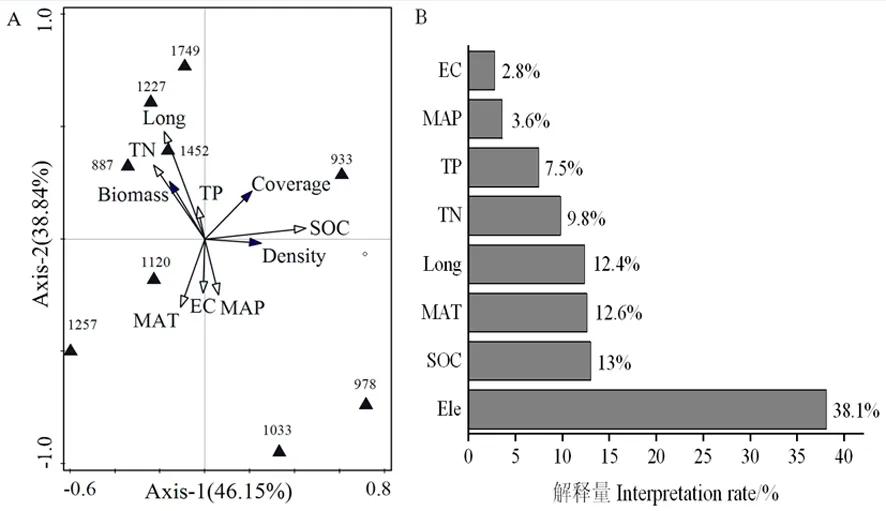
注:Ele-海拔;SOC-土壤有机碳;MAT-年均温度;Long-经度;TN-土壤全氮;TP-土壤全磷;MAP-年均降水量;EC-土壤电导率。A图中三角形上方数字代表海拔
3 讨 论
3.1通过依照植物个体的形态、生活史等生物学特性来划分植物经济类群的方法代替具体物种来描述植物群落的构成更具有一定的简便性[14]。植物经济类群对草地植物群落生产力、稳定性和退化恢复具有重要的影响。研究中,天山北坡4种蒿类荒漠生态经济类群重要值的空间差异较为明显,半灌木表现为由西向东先降后增,多年生草本呈现逐渐降低的变化规律,而一年生草本和短生、类短生草本重要值在东段达到最低值。可能是天山北坡东段在冬季降雪量及夏季降雨量小于西段和中段,春季植物返青时土壤中的水分相对较少,导致一年生草本和短生、类短生植物萌发的数量和种类较少,致使天山东段的半灌木的重要值较高,植被的高度、盖度、密度和生物量均低于天山西段和中段。群落数量特征能够反映出草地的健康状况和草场质量,其变化在一定程度上还能反映出群落受外界影响的程度。天山北坡中段的群落高度、盖度和密度显著高于西段和东段(P<0.05),且天山北坡中段和东段的群落生物量显著高于西段(P<0.05)。植物群落数量特征均表现为中段大于西段和东段,即由西向东呈现先增高后降低的变化趋势,天山北坡蒿类荒漠中段植物种类与西段和东段相比较多,其中包括弯果葫芦巴、落芒草和葶苈,致使中段的植物群落数量最高。
3.2α多样性是草地群落物种丰富度、均匀度和多样性的重要测量指标,能够充分反映物种间通过竞争而产生对外界环境的适应能力[15]。空间异质性(地形、地貌)会影响植物多样性,同时环境因子的改变也是影响植物多样性差异的重要原因[16-17]。研究表明,天山北坡西中东段植物的丰富度指数、物种多样性指数和均匀度指数均具有较大的差异性,而优势度指数则均无差异。另外,研究也发现,丰富度指数、优势度指数和物种多样性指数在空间上均表现为由西向东先增后降的变化趋势,而均匀度指数在空间上的变异规律不同于其他3个多样性指数,其呈现的规律表现为由西向东逐渐降低。原因可能是(1)天山北坡由西向东降水量逐渐降低,生态环境愈加脆弱,所以植物多样性均在东段呈现最低值。(2)在植物群落物种多样性的海拔分布格局中,干旱地区的植物物种多样性随海拔的升高而降低[18-19]。(3)因试验样地当年所属区域水热条件、人为干扰程度等方面的不同,导致某一段的植物多样性偏高或偏低进而影响研究结果,因此需要多维度且长期观测研究天山北坡不同区域的植物多样性的变化规律。此外,在整个研究区,蒿类荒漠植物群落结构和多样性主要受海拔、土壤有机碳含量和年均温度的影响,其原因主要为荒漠气候干燥、日照长,造成荒漠植被稀疏、结构和营养级简单,导致荒漠土壤养分含量较低,所以荒漠植物群落结构及多样性主要受3个环境因子调控。
4 结 论
4.1天山北坡蒿类荒漠半灌木生物量由西至东呈逐渐增加趋势,且东段比西段显著高了266.51%(P<0.05),而多年生草本从西到东则基本呈现逐渐降低的趋势,且西段显著高于东段(P<0.05),一年生草本和短生、类短生植物在数量特征上均在中段呈现最大值。
4.2天山北坡蒿类荒漠群落高度、盖度、密度和生物量均呈现由西向东先增后降的变化规律,其中在中段达到最大值。Patrick丰富度指数、Simpson优势度指数、Shannon-Wiener物种多样性指数在空间上均表现为由西向东先增后降的变化趋势,而Pielou均匀度指数则呈现由西向东逐渐降低趋势。天山北坡中段植物群落的生长较西段和东段旺盛。
4.3天山北坡不同区域草地之间植物群落数量特征与多样性的相关性各有差异,蒿类荒漠草地群落盖度、密度和生物量对植物多样性呈正相关,而植物高度对植物多样性呈负相关。
4.4引起蒿类荒漠植物群落空间变异的主要影响因素是海拔、土壤有机碳、年均温度。
猜你喜欢
矿产勘查(2021年3期)2021-07-20
绿色科技(2021年4期)2021-04-06
西部交通科技(2021年9期)2021-01-11
矿产勘查(2020年2期)2020-12-28
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
震灾防御技术(2019年3期)2019-06-02
石油工程建设(2014年5期)2014-03-20
新疆农垦科技(2014年10期)2014-02-28
地质找矿论丛(2014年2期)2014-02-27
城市道桥与防洪(2014年7期)2014-02-27
