
结合洄游模式的中西太平洋鲣资源评估
2023-10-19 06:05黄雨辰耿喆冯佶石一茜朱江峰
大连海洋大学学报 2023年4期
黄雨辰,耿喆,3,冯佶,3,石一茜,3,朱江峰,3*
(1.上海海洋大学 海洋科学学院,上海 201306;2.大洋渔业资源可持续开发教育部重点实验室,上海 201306;3.农业农村部大洋渔业开发重点实验室,上海 201306)
鲣(Katsuwonuspelamis)是一种大洋中上层高度洄游性鱼类,其在中西太平洋的产量占全球鲣总产量的70%[1],围网和竿钓为其主要捕捞方式。鲣种群具有高度洄游性,生产活动中具有渔业作业类型多、管理区域复杂等特点[1]。鲣资源管理由中西太平洋渔业委员会(Western and Central Pacific Fisheries Commission,WCPFC)负责,从20世纪80年代开始,在该海域对鲣的捕捞强度逐年增长,并于2019年达到历史高位(年产量为204.60万 t)。由于鲣具有较高的繁殖力和生长速度,故该资源未发生过度捕捞[2-4]。但考虑到逐年增长的捕捞压力和资源评估的复杂性,WCPFC建议各成员国和地区的学者进一步优化资源评估,以减少评估不确定性并降低过度捕捞风险[5]。
目前,鲣资源评估的问题主要表现在:1)种群繁殖力较高且年际波动大,导致种群的幼龄生物量占总生物量的比重较大,无年龄结构的资源评估方法因无法考虑选择性,难以准确解释种群动态与资源丰度的关联性;2)不同作业方式对个体体长的选择性差异较大,若不能对其进行细分则会低估渔业对种群年龄结构的影响,并增加资源丰度和体长结构数据拟合的不确定性[6];3)高度洄游会造成不均匀的资源分布,也会使不同季节和区域的观测数据间存在矛盾,增加评估模型的拟合误差[5]。
国内对中西太平洋鲣的研究多集中在生物学特性和渔场分布方面,如王学昉等[7]和李亚楠等[8]分别对鲣的生物学特性及气候因子影响渔场分布进行了研究,而资源评估方面鲜有研究报道。目前,对中西太平洋鲣资源的评估主要采用MULTIFAN-CL(MFCL)模型,对大西洋、印度洋和东太平洋鲣资源的评估则采用Stock Synthesis 3(SS3)模型[9-11]。MFCL模型可将多组渔业进行混合并直接生成预测数据进行似然方程拟合,而SS3模型将不同渔业划分为不同单元进行独立计算和加权拟合,因此,MFCL在拟合多组渔业数据时更简洁有效;但在考虑洄游情况时,SS3模型通过假设种群洄游服从多种分布方式,并赋予各区域的群体具有不同的生活史特征及更直观的生物学解释。评估中西太平洋鲣资源时,仅用标志放流数据分析个体移动,由于标志缺失和回捕率低等因素,这一研究存在较大不确定性。本研究中,采用SS3模型并结合分季节、分年龄的洄游模式参数化,重点分析了洄游过程对鲣资源评估的影响[10,12-14],以期为高度洄游鱼类的资源评估改进和制定管理策略提供科学参考。
1 材料与方法
1.1 鲣渔业及其数据
WCPFC将鲣渔业管理区域划分为8个,见图1,鲣渔业类型划分和数据时间跨度见表1,上岸渔获量见图2。所有数据均来自WCPFC与太平洋共同体(The Pacific Community,SPC)秘书处(http://www.wcpfc.int/statistical-bulletins)[5,15-18]。
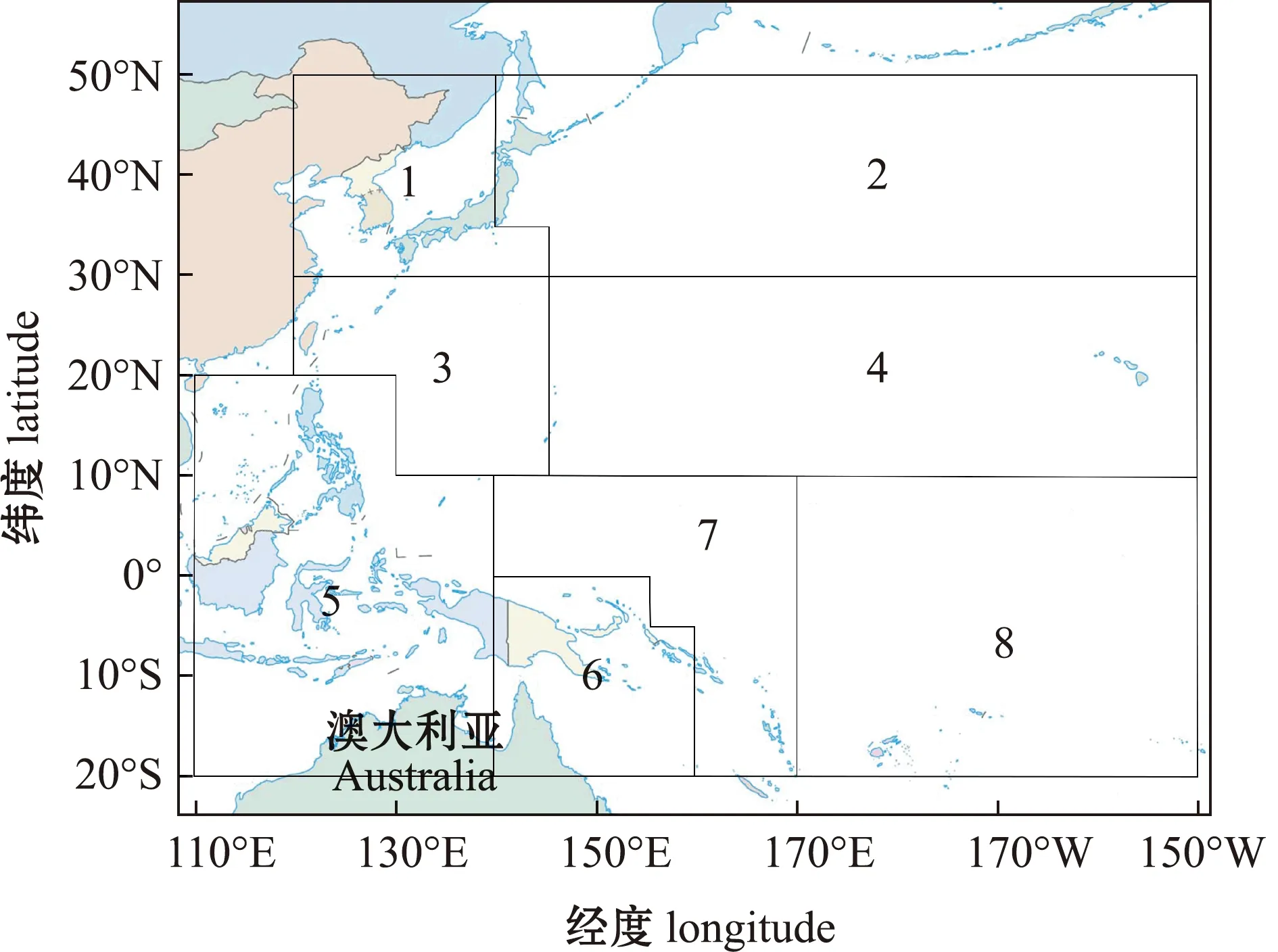
本图基于自然资源部标准地图服务网站GS(2016)1663号标准地图为底图,底图边界无修改。1~8为区域。The figure is based on the standard map GS(2016)1663 in the Standard Map Service website of Ministry of Natural Resources of the People’s Republic of China,with no modifications of the boundaries in the standard map.1-8,area.图1 中西太平洋鲣渔业管理区域划分Fig.1 Management area delineation for the Western and Central Pacific skipjack tuna fisheries
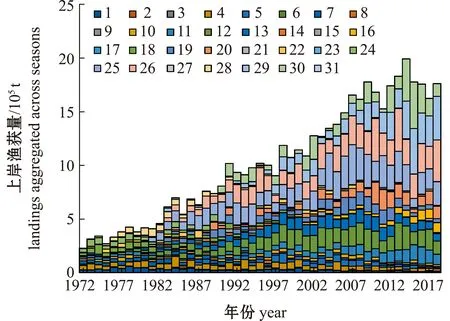
图例表示不同船队,详见表1。The legend indicates different fleets,see Table 1.图2 中西太平洋鲣历年渔获组成Fig.2 Historical catch of skipjack tuna in the Western and Central Pacific
1.2 方法
1.2.1 评估模型与假设 SS3模型有3个子模型,即:1)种群动态模型,用以描述种群生物学特征及渔业作用下的种群动态变化;2)观测模型,用来建立观测值的预测模型(如丰度指数与资源量关系);3)统计模型,用来统计模型预测值与观测值间的误差结构,并量化估算误差,模型构建基于C++,模拟通过R语言实现[19-21]。
1.2.2 种群动态模型 本研究中考虑的种群动态和渔业过程包括生长、繁殖、亲体与补充量的关系、自然死亡、种群洄游及渔具选择性等,指标的详细计算公式可参照朱江峰[11]和Methot等[19]的研究。其中,选择性的计算公式为
Sf,l=1/{1+exp[-ln19(l-Ls50)]/
(Ls95-Ls50)}。
(1)
式中:Sf,l为f船队中l体长组渔具选择的概率;Ls50和Ls95分别为f船队中50%和95%选择性的对应体长。
1.2.3 观测模型 SS3观测模型为
(2)
(3)
1.2.4 统计模型 具有相对资源丰度观测数据的渔业,对应的对数正态分布似然函数为
(4)
式中:L为目标函数;N为观测个数;I为渔获量丰度指数的观测值(即CPUE值);σ为CPUE对数化后正态分布误差结构的标准差。
具有体长组成观测资料的渔业对应的多项式分布似然函数为
(5)
1.2.5 洄游过程的建模 除基于个体模型(individual based model)的资源评估方法外,多数评估模型存在资源均匀分布的假设,由于计算机性能和过参数化的限制,对研究区域进行完全栅格化将会导致模型需要的评估参数数量呈指数化增长,因此,当前主流的单鱼种资源评估方法是通过区域划分来满足资源量化评估和渔业管理的需要,但在每个划分区域内资源仍维持均匀分布的假设[22]。其中,SS3模型就是可以结合空间结构,并在不同区域赋予种群不同生活史特征的模型,空间结构的考虑可以使模型更加灵活,同时,也可以更好地验证资源在开发和管理过程中关于洄游模式的各种假设。
本研究中的种群运动在每个季节开始时瞬时发生,采用“隐性过度”(implicit transition)的计算对拥有共同边界的区域参数进行估计[19],因此,鱼群可以在非毗连区域进行移动,达到划分区域内的全面覆盖(图1)。p′和p区域间的移动量随年龄a线性增加或减少,其计算公式为
(6)
(7)
式中:Xp′,p,a为p′和p区域间a龄鱼的移动量;xp′,p,a为p′和p区域间a龄鱼的移动概率;mα,p′,p为aα龄的鱼群从区域p′移动到区域p的概率,aα为鱼群初始移动的年龄,本研究中设为1龄;mω,p′,p为aω龄的鱼群从区域p′移动到区域p的概率,aω为鱼群结束移动的年龄,本研究中设为12龄;λp为常数。
季节性洄游概率通过最低年龄迁移参数p1和最高年龄洄游迁移参数p2进行调节,其余年龄段呈p1至p2的线性分布,具体计算公式为
(8)
式中:ri为年龄为i的迁移率;N为最终可能留在pi区域的鱼群总数,也包含其留在原地未发生洄游的可能。移动参数化定义为:在一个给定区域内移动到临近区域的鱼群比例,以区域1为例,该区域种群每个季度达到洄游年龄后可洄游至区域2和区域3,或者停留在区域1。当种群不发生洄游时,参数p=1,迁移相对发生比率数值无独立含义。
1.2.6 参数设置 基于以往研究结果[23-24],本研究中各渔业的可捕系数(catchability)服从对数贝塔分布。由于鲣渔业各作业方式差异较大,为赋予模型最大灵活性,本研究中渔业选择性曲线均采用双正态分布(double-normal),并对控制选择性曲线弯曲度参数和顶端峰值参数分别设定为基于均匀分布和正态分布的先验信息(表2)。

表2 鲣生活史信息和种群动态模型的参数设置Tab.2 Parameter settings of life history information and population dynamic models for skipjack tuna
本研究中,在考虑洄游的基准模型基础上设置了忽略种群洄游的情景,以探究洄游结构对资源评估结果的影响。此外,为考虑评估模型参数设置的不确定性,并探究各参数设置对模型结果的影响,设置了4种不同情景对模型进行敏感性分析(表3)。其中,陡度参数h是年龄结构模型中主要的不确定性来源,本研究中选择3种不同的陡度参数h进行敏感性分析[23],基准模型的陡度参数h取为0.8,其他h分别取0.65和0.95[5]。

表3 SS3模型敏感性分析情景设置Tab.3 Sensitivity analysis scenario settings of SS3 model
2 结果与分析
2.1 CPUE数据拟合
4组情景评估结果均收敛,梯度(gradient)均小于0.000 1。其中,考虑和忽略种群洄游的SS3模型CPUE拟合见图3,结果显示,忽略季节性洄游时拟合曲线较为平滑,难以准确捕捉季节性CPUE变动,而考虑季节性洄游时CPUE拟合曲线有明显的季节波动,且拟合效果得到显著提高。
左侧和右侧分别为情景1和2。Scenario 1 and scenario 2 are shown on the left and right respectively.图3 CPUE观测值(点圈)与预测值(实线)的拟合情况Fig.3 Observed (dotted circles)and predicted (solid lines)values of the CPUE
2.2 选择性拟合
以区域3的渔具选择性为例(图4),结果显示:围网渔业(PS_ALL_3)对鲣的开捕年龄较小且捕捞以小体长为主,集中分布为40~75 cm的鲣个体;延绳钓渔业(LL_ALL_3)的捕捞体长最为集中,主要捕捞65~70 cm的个体;而竿钓渔业(PL_ALL_3)集中捕捞体长为40~75 cm的个体,其渔具选择性范围最大。对相同渔具渔业的模拟有较好的表现(图5),如船队2、15和20的渔具选择性基本相同,对大于5龄的鲣个体具有完全选择,而船队10、11和16这3种沿海小型渔业则对小年龄鲣个体(2、3龄)有更强的倾向性。

图4 渔区3的围网、延绳钓和竿钓渔业渔具的选择性Fig.4 Gear selectivity in purse seine,longline and pole-and-line fisheries in the area 3
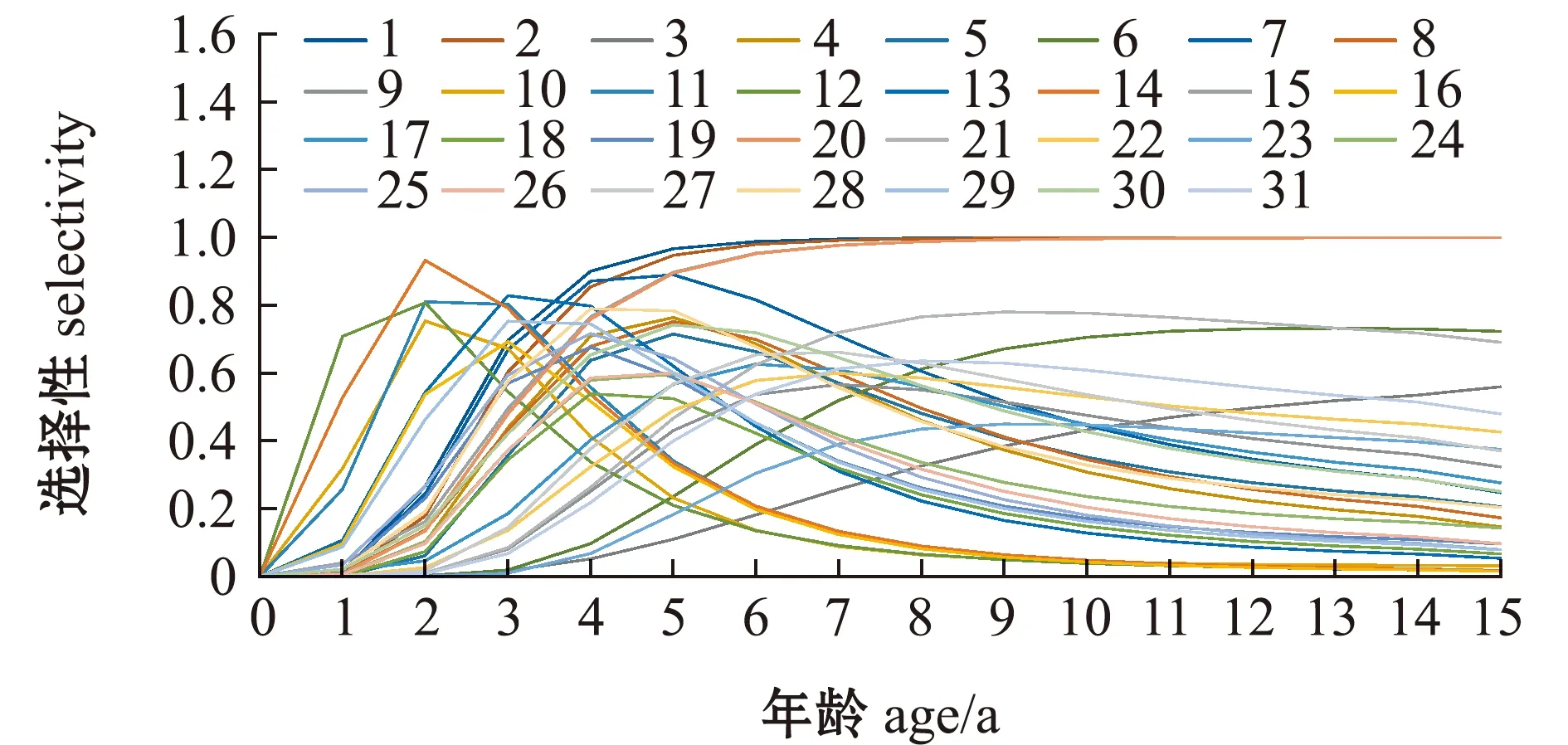
图例表示不同船队,详见表1。The legend indicates different fleets,see Table 1.图5 各渔业渔具对鲣的年龄选择性Fig.5 Selectivity at age of skipjack tuna in the Western and Central Pacific Ocean by different fishing gears
从图6可见,多数渔业拟合情况良好,仅有船队14(PS_ASSOC_5)和15(PS_UNASSOC_5)中自由鱼群和随附鱼群的围网拟合存在偏差。
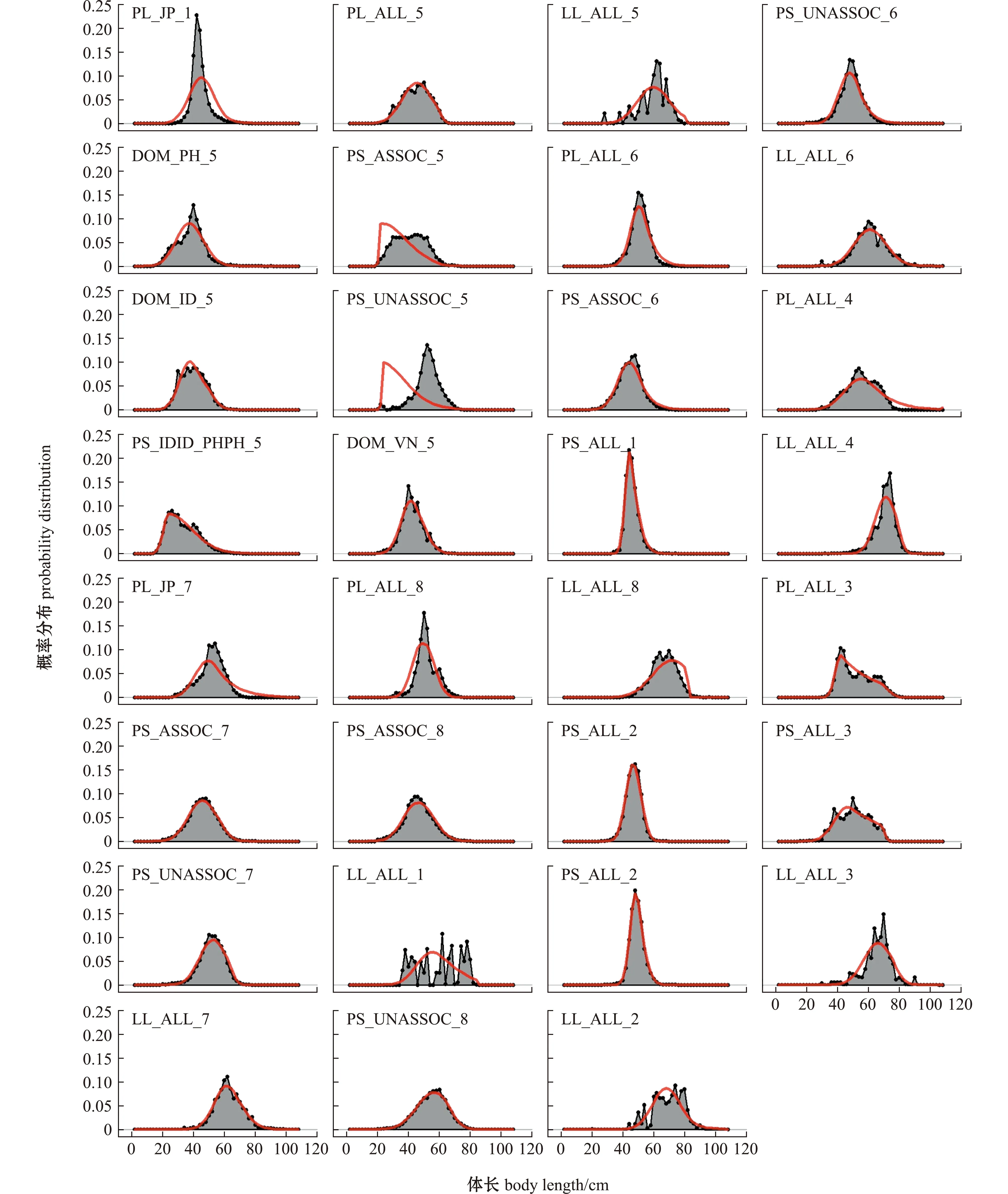
图中红色曲线为估算后验分布。The red curve in the figure means posteriori distribution.图6 不同渔业方式的体长组成Fig.6 Length composition in different fisheries
2.3 洄游模式
洄游模拟结果显示,不同季节下不同年龄段的鲣鱼群有着不同的洄游模式(图7)。

图7 中西太平洋1~12龄鲣不同季节的迁移率(迁移率均小于1)Fig.7 Seasonal movement rate of 1 to 12 years old skipjack tuna in the Western and Central Pacific Ocean (all of movement rates are less than 1)
春季(1—3月),1~8龄的鲣鱼群在1、2、3区域较为活跃,小部分4区域和大部分7区域的鱼群会向西北移动至3区域,并在该区域聚集。而区域4和8间存在部分鱼群的来回移动。大龄鱼群(9~12龄)则更倾向于驻留在原区域。
夏季(4—6月),所有年龄的鲣均有相似的洄游模式,大部分7区域和4区域的鱼群会向8区域进行东南向移动,并在8区域聚集,但是从8区域迁出的个体却较少。从6龄开始,逐渐有部分鱼群开始从6区域向5区域进行西向移动,并在8龄时迁移率达到最大值,一直持续至12龄。此外,还有少部分鱼群会从3区域向5、7区域进行移动,但从9龄(9~12龄)开始,3区域向5、7区域的移动逐渐消失。
秋季(7—9月),所有年龄的鲣鱼群总体呈现向南的运动趋势,大部分3区域鱼群会向5区域进行移动,2区域鱼群会向4区域进行移动,但是从4区域迁出的鱼群却很少。此外,还有少部分鱼群会从6区域经7区域向8区域进行东向移动,这些小部分鱼群的区域间移动在9龄时开始逐渐消失,但大范围的南向移动依然存在。
冬季(10—12月),1~8龄的鲣鱼群会从1、2、3区域向4区域进行移动,少部分4区域的鱼群会向8区域进行移动,但是从8区域迁出的个体几乎没有。9~12龄的鱼群则变得开始只在1、2、3区域活动。此外,所有年龄的鱼群均存在大量从6、7区域向5区域西向移动的情况。
总的来说,鲣鱼群在1、2、3区域非常活跃,并在4和8区域进行聚集,但从这些区域迁出的个体数量极少。此外,在5、6、7、8区域间存在季节性的东西向移动,在3、7区域间存在季节性的南北向移动。
2.4 资源状况
从图8可见,鲣资源有较为稳定且数量较大的补充量,1995年前后产卵生物量达到历史顶峰,随后在1995—2008年,产卵生物量虽有波动但整体趋势平稳,2008年后,产卵生物量呈逐年下降的趋势,于2020年下降到资源量低位。

图8 中西太平洋鲣产卵生物量补充量偏差和季节性趋势变化Fig.8 Seasonal variations in spawning biomass and recruitment deviation of Western and Central Pacific skipjack tuna
多数情景下,鲣的资源量处于健康状况,既未发生资源型过度捕捞(B2018>BMSY;Bcur/B40%=1.9>1),也未发生捕捞型过度捕捞(F2018
3 讨论
3.1 SS3模型敏感性分析
对陡度、洄游参数等主要不确定性来源进行敏感性测试,可以了解模型输出结果的稳健性,为正确选择参数、构建模型提供参考。本研究中,通过探究不同渔具的选择性差异和区域间种群洄游现象,显著提高了SS3模型对CPUE和体长数据的拟合度,使中西太平洋鲣资源评估模型的合理性进一步提升。从模型拟合结果来看,仅有船队14和船队15的体长数据拟合存在较大误差,虽然SS3模型具有较大的灵活性,但模型参数拟合过程受体长和CPUE数据的共同影响,该情况可能是因体长组成与丰度数据不匹配造成(如相对丰度指数数据倾向于捕捞小型个体,而相同渔具下体长数据却倾向于捕捞更大体长的个体)。此外,围网渔业在统计渔获时也存在将小型个体直接丢弃及未鉴定种类统称为“杂鱼”的情况,该部分小型渔获物可能未记入鲣数据中[14]。
本研究中,模型敏感性分析结果显示,鲣种群的补充量和陡度参数h对生物学参考点的计算结果影响较大,由于该参数定义为资源量接近于零值时补充量的斜率,因此,难以通过生物学进行直接估计。在当前资源评估中,通常基于以往研究和经验进行设置或将其设定为灵活参数,并通过模型拟合进行估计。Zhu等[25]研究表明,资源评估模型仅能在一定范围内对参数h进行估算,陡度参数对评估结果的影响较明显且容易造成参数过拟合的情况,使模型背离或失去生物学意义。种群统计学可以将生活史信息和种群动力学相结合,提出基于生物学研究的有信息的陡度参数先验分布,使资源评估模型在保持生物学意义的前提下依然可以对该参数进行估计[26]。
3.2 洄游模式的拟合状况
资源评估中通过增加空间结构估算鲣的空间移动率可以了解其洄游模式,一方面有利于了解鲣的移动路径及渔场分布,提高渔业生产效率;另一方面明确其具体位置,可以使养护和管理具有针对性,从而提高鲣的养护和管理效果。本研究中,使用SS3模型对中西太平洋鲣进行了不同年龄阶段的空间种群动态拟合,结果显示,鲣鱼群在1、2、3区域非常活跃,而且会在4和8区域进行聚集,但种群迁出的个体数量极少。此外,在5、6、7、8区域间存在季节性的东西向移动,在3、7区域存在季节性的南北移动。目前,全球范围内关于鲣洄游的研究均较少,仅发现分布在马绍尔群岛和加罗林群岛的鲣群体会向日本、菲律宾和巴布亚新几内亚洄游,而分布在马绍尔群岛和法属波利尼西亚附近的鲣群体会向非洲西岸和夏威夷群岛洄游[3,27-30]。Kiyofuji等[27]研究认为,日本南部海域(即本研究区域2西侧)是鲣的索饵场,夏季黑潮带来大量的日本鳀和沙丁鱼为附近鲣提供了大量的饵料,导致鲣有向该区域洄游的现象。Schaefer等[28]研究表明,东太平洋鲣会在冬末和春初的几个月内在中美洲(区域8东侧)和夏威夷群岛附近[29](区域4)进行产卵,这也与本研究结果基本吻合。此外,在5、6、7和8区域间的季节性东西向移动可能与赤道南北洋流的推动有关,而3、7区域的季节性南北移动可能是由于海水温度和饵料丰度影响所致。
除此之外,气候变动也是影响鲣分布的重要因素之一,鲣是一种对温度较为敏感的鱼类,当海表温度发生剧烈波动时(如厄尔尼诺或拉尼娜现象),其垂直和水平运动均会变得剧烈且会进行较长距离的移动,以寻求适宜栖息地[5]。杨胜龙等[31]通过对海洋环境因子(海温、盐度和叶绿素等)的变化研究发现,当厄尔尼诺发生时,中西太平洋鲣渔场中心会向东移,而当拉尼娜发生时,中心则会向西收缩。但是环境因子具体如何影响中西太平洋鲣的移动路径有待进一步研究。
中西太平洋鲣资源区域间的长距离大量迁移,一方面可能是因海表温度变化导致的季节性往返运动;另一方面热带地区的丰富饵料供给及适宜的产卵条件导致大量的鲣向该方向移动,而太平洋西侧黑潮的推动,导致鲣移动向南延伸。
3.3 SS3模型在渔业管理中的应用效果
对资源进行评估最主要的目的是了解目前资源状况并对其进行针对性管理,不同管理目标参照系下的结果会存在较大差异,因此,需要对不同管理目标下的评估结果进行讨论和比较。本研究中,基于SS3模型的评估结果显示,鲣资源群体未发生资源型过度捕捞(Bcur/B40%=1.9>1)和捕捞型过度捕捞(Fcur/F40%=0.712<1),该结果与中西太平洋渔业委员会基于MULTIFAN-CL模型的评估结果相一致。两个研究均采用了基于生物学的不同管理参考点体系,本研究中采用初始资源的40%作为参照系(B/B40%和F/F40%),而中西太平洋渔业委员会则采用了基于最大持续产量的产卵生物量和捕捞死亡率比值(SB/SBMSY和F/FMSY)。后者通常用于不同种群间的比较,通过模型迭代计算出种群平衡状况下的产卵生物量资源状况;而前者则更注重渔业影响下非平衡的资源状况,通过渔业管理者预设的目标参考点(通常为B40%),多直接用于渔业管理中,童玉和等[32]研究表明,后者对补充量的年际变动更为敏感,也使得管理目标更为保守。
4 不足与展望
目前,中西太平洋渔业委员会使用MCFL模型对鲣进行资源评估,虽然已解决了部分模型的不确定性,但由于MCFL模型本身的限制,其是否能在评估模型中较为全面地反映种群洄游模式仍待进一步研究。本研究中,利用SS3模型探究了洄游模式在中西太平洋鲣资源评估中的可行性与必要性,由于数据均来自于真实渔业,数据的观测误差不可避免,无法对MCFL和SS3模型的评估效果进行比较和评价。该问题可以在未来研究中通过建立模拟种群的方式,以量化评估模型的结构误差并分析其影响。此外,本研究表明,考虑季节性洄游对鲣资源评估的影响是显著的,该影响不但体现在模型的拟合过程中,同样也增加了实际渔业管理的难度。目前,中西太平洋渔业委员会主要通过限制鲣的捕捞努力量(包括在周边岛国专属经济区和20°N~20°S公海禁止使用集鱼装置作业,每艘围网船最多释放350个集鱼装置等)和渔获量(不得超过2001—2004年平均水平或2004年水平)[33]进行管理,但由于资源分布不均,集中捕捞可能会造成部分区域的生长型(集中捕捞幼鱼)或补充型(集中捕捞成鱼)过度捕捞,因此,对捕捞策略进行适当优化才能更充分地开发渔业资源。管理策略评价(management strategy evaluation,MSE)通过模拟数据收集系统、资源评估模型和管理策略制定与实施等环节可能产生的误差,帮助管理者依据不同管理目标和期望(如最大程度地可持续利用、保守开发或者种群重建)制定相应的管理措施[24,34]。结合国家正在实施的公海自主休渔制度,将季节性洄游与管理策略评价研究相结合将是筛选和优化渔业管理措施的重要研究方向。
5 结论
1)季节性洄游对鲣种群资源评估具有显著影响,忽略洄游模式会低估捕捞对资源的影响。资源评估过程应该考虑种群洄游模式。
2)不同季节下不同年龄段的鲣有不同的洄游模式。总的来说,鲣鱼群在1、2、3区域非常活跃,在4和8区域聚集,但从这些区域迁出的数量极少。此外,在5、6、7、8区域间存在季节性的东西向移动,在3、7区域间存在季节性的南北向移动,说明在资源评估过程中应该考虑不同季节下不同年龄段的种群洄游情况。
猜你喜欢
艺术品鉴(2020年3期)2020-07-25
世界农药(2019年4期)2019-12-30
中外文摘(2017年19期)2017-10-10
中国水产(2017年2期)2017-02-25
湖南畜牧兽医(2016年3期)2016-06-05
电测与仪表(2016年3期)2016-04-12
电测与仪表(2016年20期)2016-04-11
陶瓷科学与艺术(2016年1期)2016-02-28
河南城建学院学报(2015年4期)2015-02-27
江苏年鉴(2014年0期)2014-03-11
