
黄脊竹蝗种群肠道微生物的多样性分析
2023-10-20 12:07杨丽君李红卫李向永谌爱东于艳雪
环境昆虫学报 2023年4期
杨丽君,李红卫,李向永,谌爱东,于艳雪*
(1.中国检验检疫科学研究院植物检验与检疫研究所,北京 100176;2. 中国农业大学植物保护学院,北京 100193;3. 云南省农业科学院农业环境资源研究所,昆明 650205)
蝗虫灾害是国际性的自然生物灾害,涉及多个国家和地区,对全球经济和环境造成严重影响(涂雄兵等, 2021)。
昆虫肠道是昆虫存储食物、消化和排泄的场所,其中定殖着大量的微生物,与宿主在长期协同进化过程中形成了紧密的互利共生关系(Engel and Moran, 2013)。一方面,肠道微生物不仅影响昆虫对植物组织的消化吸收和信息化合物的合成,而且还会为宿主提供重要的营养物质、提高宿主防御和解毒能力,进而影响宿主昆虫的选择、繁殖和发育周期等行为(Fischeretal., 2017; 周帆等, 2020)。另一方面,肠道微生物种群结构的多样性与昆虫种类、龄期、生存环境、食物的喂养条件和消化道形态等因素息息相关(鲁迎新等, 2016; 程代凤等, 2021)。近年来,随着高通量测序技术的发展,对昆虫肠道微生物的研究逐步加深,利用肠道微生物控制害虫的新理论和新策略也越来越受到广泛关注,并且其中部分研究已成功付诸于实践(王四宝和曲爽, 2017; 侯璐文等, 2019)。
鉴于昆虫肠道微生物的重要性,相关研究已越来越受国内外学者重视。叶国浚等人通过对草地贪夜蛾Spodopterafrugiperda肠道微生物宏基因组初步分析,鉴定出其优势菌属(叶国浚等,2021);章雨璐等人通过对椰心叶甲BrontispalongissimiGestro肠道共生菌进行分离培养,鉴定其相关功能并探究肠道共生菌对椰心叶甲环境适应性作用(章雨璐等, 2021);罗曼等人采用纯培养法对云南地区的思茅松毛虫DendrolimukikuchiiMatsumura 3龄幼虫进行肠道细菌多样性的研究,发现芽孢杆菌属为该虫的优势菌群(罗曼等, 2021);魏博帆等人通过研究不同肠道微生物对无菌果蝇和正常果蝇交配行为的影响,证明植物乳杆菌可通过延长交配潜伏期影响果蝇的交配行为(魏博帆等, 2021)。
黄脊竹蝗CeracriskiangsuTsai是竹蝗属昆虫中分布广泛及危害严重的林业害虫之一,其食性杂、分布广、迁飞性强,食物不足或大发生时也可为害农田粮食作物(程佳等, 2010;仓晓燕等, 2020)。其防治技术多为利用人尿进行诱集和大量使用化学药剂,但随着可持续防治理念逐渐深化,生物防治方法的研究与应用越来越广泛(李红梅等, 2021)。
根据现有文献记载,气温、降水量、日照时数、光照和风速对黄脊竹蝗的生长、产卵和迁飞具有显著影响(李建江, 1998; 张贤开和左玉香, 2005; 张卫东等, 2012),较少报道关于揭示性别和非生物因子对黄脊竹蝗肠道微生物的作用机制。因此,本文通过利用Illumina NovaSeq测序平台对新入侵地区和国内常发地区的黄脊竹蝗肠道微生物多样性进行研究,分析不同地理分布、性别及环境因子对黄脊竹蝗肠道微生物的影响,探寻蝗虫地理溯源,为开发微生物制剂和提高综合防治技术提供新思路,以保障世界粮食生产安全。
1 材料与方法
1.1 材料
1.1.1供试昆虫
采集了3个地区的黄脊竹蝗,其中云南省普洱市江城县是2020年新入侵地区,湖南省益阳市桃江县和湖南省邵阳市新宁县是国内常发地区。具体信息见表1。
表1 黄脊竹蝗采集信息表
1.1.2地理环境因子数据
在国家气象数据中心下载黄脊竹蝗采集地的5年平均气相数据并进行分析(表2)。其中,平均气温的代号为T;降水量的代号为P;相对湿度的代号为RH;日照时数的代号为SUN;风速的代号为WVEL;蒸发量的代号为EVA;平均气压的代号为Pa;地面温度的代号为GND。

表2 不同地理位置2017-2020年平均气象条件数据统计
1.2 方法
1.2.1黄脊竹蝗样品收集
随机选取不同地理位置的黄脊竹蝗做6个重复处理,其中每头为一个处理。每个地理位置蝗虫雌雄虫各3头,用酒精和PBS溶液分别冲洗两遍,每次1 min;将清洗后的虫体放在蜡盘上进行解剖,取出后肠放入离心管中,每头放入一个2 mL离心管中,然后在研磨机上研磨,研磨结束放入冰箱。
1.2.2肠道微生物基因组提取及检测
按照试剂盒的步骤提取样本组织DNA:
(1)将研磨结束放置冰箱的样品取出置于冰上。
(2)向每个样品中加入1 mL Inhibit EX Buffer,涡旋持续1 min,直至样品混均。
(3)将混合液放入70℃的水浴锅中加热 5 min,涡旋15 s,室温下20 000 xg离心样品1 min。
(4)向一个新的1.5 mL离心管中加入15 μL Proteinase K,然后从步骤3中转移200 μL上清液加入。
(5)在混合液中加入200 μL Buffer AL,涡旋混匀,将混合液放入70℃的水浴锅中孵育10 min。
(6)向混合液中加入200 μL(96%~100%)酒精,涡旋充分混合。
(7)将步骤6中的全部混合液滴到新QIAamp离心柱的中央,离心1 min,丢弃沉淀物。
(8)加入500 μL Buffer AW1,离心1 min,丢弃含滤液的管。
(9)加入500 μL Buffer AW2,离心3 min,丢弃含滤液的管。
(10)将离心柱转移到一个新的1.5 mL微量离心管中,将200 μL Buffer ATE滴到QIAamp膜中央位置。室温孵育1 min,然后20 000 xg离心1 min洗脱DNA。
(11)对提取的DNA进行凝胶电泳检测。
1.2.3PCR扩增和纯化
将提取的黄脊竹蝗肠道基因组DNA作为PCR的模板,采用16S V3-V4扩增引物(341 F: 5′-CCTAYGGGRBGCASCAG-3′和806 R: 5′-GGACTAC NNGGGTATCTAAT-3′)进行PCR扩增。使用2%琼脂糖凝胶进行电泳检测。然后根据PCR产物浓度进行等浓度混样,充分混匀后使用1×TAE浓度为2%琼脂糖胶电泳纯化PCR产物,割胶回收目标条带。
1.2.4文库构建和上机测序
使用Illumina公司TruSeq DNA PCR-Free Library Preparation Kit建库试剂盒进行文库的构建,构建好的文库经过Qubit定量和文库检测,合格后进行上机测序。测序得到的原始数据存在一定的干扰数据,为了使信息分析的结果更加准确可靠,首先根据barcode进行拆分获得每个样品的原始数据,并去除barcode和引物,使用FLASH软件数据进行拼接。随后通过质控得到Clean Tags,再进行嵌合体过滤,得到可用于后续分析的有效数据。最后基于有效数据通过DADA2进行降噪,并过滤掉丰度小于5的序列,从而获得最终的ASVs(Lietal., 2020)。
对于得到的ASVs(Callahanetal., 2017),一方面对每个ASV的代表序列做物种注释,得到对应的物种信息和基于物种的丰度分布情况,进行Alpha多样性分析,以得到样本内物种丰富度和均匀度信息等。另一方面,可以对ASVs进行多序列比对并构建系统发育树,通过无度量多维标定法(NMDS),探究不同样本或组别间群落结构的差异。最后,进一步挖掘分组样本间的群落结构差异,选用LEfSe统计分析方法对分组样本的物种组成和群落结构进行差异显著性检验,用Spearman秩相关来研究环境因子与微生物种丰富度之间的相互变化关系,得到两两之间的相关性。
2 结果与分析
2.1 序列拼接组装与OTU聚类分析
通过Illumina NovaSeq测序平台对不同地理分布的黄脊竹蝗肠道微生物16S的V3~V4区域进行测序,发现不同地理分布和性别的黄脊竹蝗肠道菌群测序所得的条带数都有所差异(表3)。其中来自湖南省邵阳市新宁县的雌成虫(HSXA-F)样品得到的优化序列数最多(104 993条),湖南省邵阳市新宁县的雄成虫(HSXA-M)样品优化序列数量最少(95 742条)。6个样品通过测序平均得到103 136对Reads,经拼接、过滤后共平均产生100 824条Clean Tags,有效序列所占比例为97.7%,样品的测序准确度较好,满足分析标准。每个样品分别获得的OTU数目如表3所示:对比不同地理位置的成虫,OTU数目最多的是湖南省益阳市桃江县的成虫,最少的是湖南省邵阳市新宁县的成虫;对比不同性别的成虫,3个地区的黄脊竹蝗雌雄虫肠道OTU个数没有显著差异。各个地理分布的黄脊竹蝗肠道微生物在不同的分类阶元注释出的物种数目如表3所示:其中云南省普洱市江城县雄虫(YPJA-M)样品在各个分类阶元所得到的注释数均略高于其他样品,有22个门,34个纲,82个目,145个科,282个属,362个种。湖南省邵阳市新宁县雄虫(HSXA-M)样品在各个分类阶元的注释数均略低于其他样品。

表3 黄脊竹蝗肠道细菌16S rDNA高通量测序基本信息
2.2 黄脊竹蝗肠道微生物的组成分析
黄脊竹蝗肠道样本在门分类阶元水平上的微生物群落组成如图1所示。主要有绿弯菌门Chloroflexi、酸杆菌门Acidobacteriota、Planctomycetota、粘球菌门Mvxococcota、梭杆菌门Fusobacteriota、蓝细菌门Cyanobacteria、拟杆菌门Bacteroidota、放线菌门Actinobacteriota、厚壁菌门Firmicutes、变形菌门Proteobacteria等。湖南省益阳市桃江县(HYT)、湖南省邵阳市新宁县(HSX)和云南省普洱市江城县(YPJ)3个地理位置样品的优势菌门均为变形菌门(46.7%、77.0%、64.3%)。其中湖南省邵阳市新宁县(HSX)的次优势菌门为放线菌门(19.0%)和厚壁菌门(18.5%),云南省普洱市江城县(YPJ)的次优势菌门为厚壁菌门(20.2%),湖南省益阳市桃江县(HYT)的次优势菌门为厚壁菌门(11.4%)和放线菌门(11.3%)。3个地区不同性别样品的肠道菌群门水平组成差异不显著。
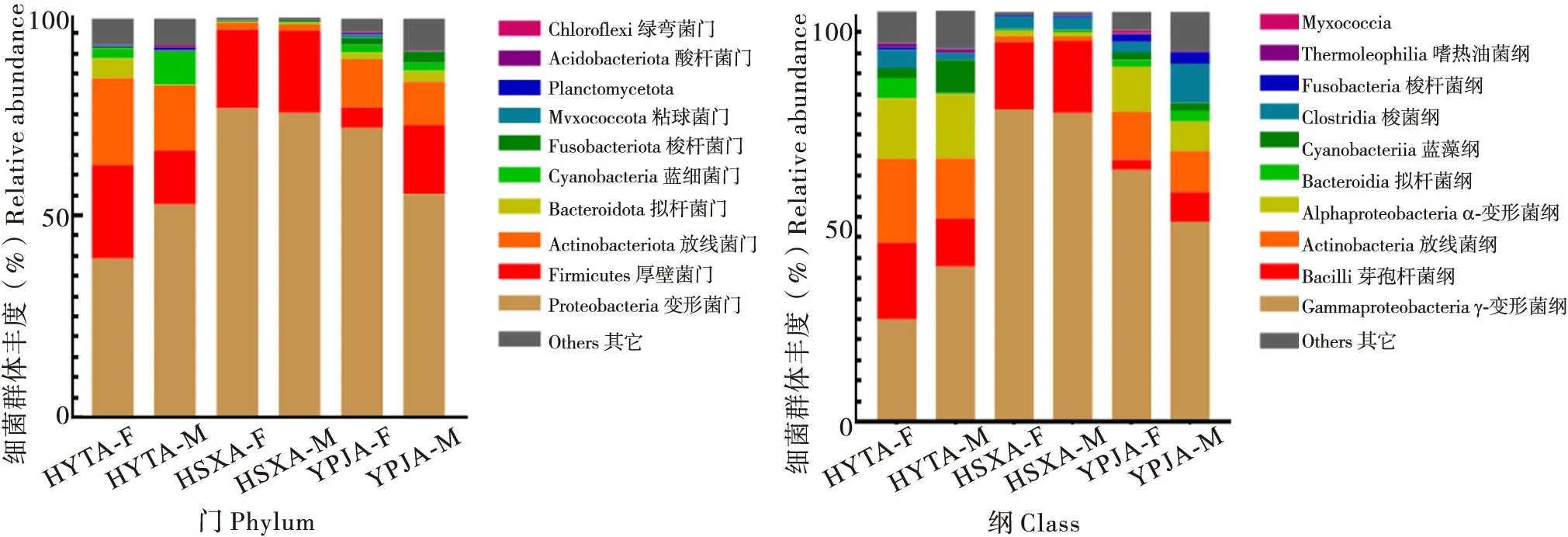
图1 不同地理种群的黄脊竹蝗前十个肠道菌群组成Fig.1 Composition of the top ten gut microbiota of Ceracris kiangsu in different geographical populations
在纲分类阶元水平上,黄脊竹蝗肠道微生物主要有嗜热油菌纲Thermoleophilia、梭杆菌纲Fusobacteria、Myxococcia、梭菌纲Clostridia、蓝藻纲Cyanobacteriia、拟杆菌纲Bacteroidia、α-变形菌纲Alphaproteobacteria、放线菌纲Actinobacteria、芽孢杆菌纲Bacilli、γ-变形菌纲Gammaproteobacteria等。湖南省益阳市桃江县(HYT)、湖南省邵阳市新宁县(HSX)和云南省普洱市江城县(YPJ)3个地理位置样品的优势菌纲均为 γ-变形菌纲(31.4%、75.9%、55.1%)。其中湖南省邵阳市新宁县(HSX)的次优势菌纲为芽孢杆菌纲(17.0%),云南省普洱市江城县(YPJ)的次优势菌纲为放线菌纲(10.8%),湖南省益阳市桃江县(HYT)的次优势菌纲为放线菌纲(17.5%)、芽孢杆菌纲(15.3%)和 α-变形菌纲(15.3%)。3个地区不同性别样品的肠道菌群纲水平组成没有显著差异。
在目分类阶元水平上,黄脊竹蝗肠道微生物主要有伯克氏菌目Burkholderiales、小单孢菌目Micromonosporales、黄杆菌目Flavobacteriales、虫原体目Entomoplasmatales、微球菌目Micrococcales、根瘤菌目Rhizobiales、乳杆菌目Lactobacillales、假单胞菌目Pseudomonadales、肠杆菌目Enterobacterales等。湖南省益阳市桃江县(HYT)、湖南省邵阳市新宁县(HSX)和云南省普洱市江城县(YPJ)3个地理位置样品的优势菌目均为肠杆菌目(25.7%、60.4%、50.2%)。其中湖南省邵阳市新宁县(HSX)的次优势菌目为乳杆菌目(16.3%)和假单胞菌目(14.3%),云南省普洱市江城县(YPJ)的次优势菌目为其他(10%),湖南省益阳市桃江县(HYT)的次优势菌目为乳杆菌目(12.6%)和根瘤菌目(11.2%)。3个地区不同性别样品的肠道菌群目水平组成没有显著差异。
在科分类阶元水平上,黄脊竹蝗肠道微生物主要有黄色杆菌科Xanthobacteraceae、周蝶菌科Weeksellaceae、微杆菌科Microbacteriaceae、螺原体科Spiroplasmataceae、欧文菌科Erwiniaceae、链球菌科Streptococcaceae、肠球菌科Enterococcaceae、莫拉氏菌科Moraxellaceae、肠杆菌科Enterobacteriaceae等。其中湖南省益阳市桃江县(HYT)、湖南省邵阳市新宁县(HSX)和云南省普洱市江城县(YPJ)3个地理位置样品的优势菌科均为肠杆菌科(24.3%、53.6%、39.5%)。其中湖南省邵阳市新宁县(HSX)的次优势菌科为莫拉氏菌科(12.1%)和肠球菌科(9.7%),云南省普洱市江城县(YPJ)的次优势菌科为其他(5%),湖南省益阳市桃江县(HYT)的次优势菌科为链球菌科(6.1%)、梭菌科(5.3%)和肠球菌科(4.6%)。3个地区不同性别样品的肠道菌群科水平组成没有显著差异。
在属分类阶元水平上,黄脊竹蝗肠道微生物主要有Pseudocitrobacter、李长文氏赤水河菌属Chishuiella、短小杆菌属Curtobacterium、螺原体属Spiroplasma、泛菌属Pantoea、乳球菌属Lactococcus、肠球菌属Enterococcus、不动杆菌属Acinetobacter、肠杆菌属Enterobacter等。湖南省益阳市桃江县(HYT)样品的次优势菌属有乳球菌属(6.0%)、肠杆菌属(4.8%)和肠球菌属(4.6%)。云南省普洱市江城县(YPJ)样品的优势菌属为肠杆菌属(27.5%),次优势菌属为其他(5%)。湖南省邵阳市新宁县(HSX)样品的优势菌属为肠杆菌属(16.8%),次优势菌属为不动杆菌属(12.1%)和肠球菌属(9.7%)。3个地区不同性别样品的肠道菌群属水平组成和丰度具有显著差异。其中,湖南省益阳市桃江县(HYT)的雌、雄样品优势菌属分别为乳球菌属(9.0%、2.9%)和肠杆菌属(3.7%、5.9%),云南省普洱市江城县(YPJ)的雌、雄样品优势菌属均为肠杆菌属(27.9%、27.0%),湖南省邵阳市新宁县(HSX)的雌、雄样品优势菌属分别为肠杆菌属(29.9%、3.5%)和乳球菌属(0.4%、12.0%)。
2.3 黄脊竹蝗肠道微生物Alpha多样性分析
Alpha多样性指数评估可以反映样本内菌群的多样性。本研究选用的Alpha多样性指数为Chao 1、Goods_coverage、Shannon和Simpson指数(表4)。其中,Chao 1值越大,表示样品群落中低丰度物种最多;Shannon和Simpson指数值越大,表示样品中群落丰富度和均匀度越高。结果显示:不同地理分布和性别的黄脊竹蝗肠道微生物多样性各不相同。其中Chao 1值最大的为湖南省益阳市桃江县的雄虫(HYTA-M),样品群落中低丰度物种最多,最小的为湖南省邵阳市新宁县的雄虫(HSXA-M),样品群落中低丰度物种最少。从Shannon和Simpson指数可以看出,湖南省益阳市桃江县雌虫(HYTA-F)的物种丰富度和均匀度最高,而湖南省邵阳市新宁县雌虫(HSXA-F)的物种丰富度和均匀度最低。从Goods_coverage来看,所有地理种群样品的Goods_coverage指数均接近于1,说明样品中序列检测率很高,该测序结果基本符合样品中肠道微生物的真实情况。
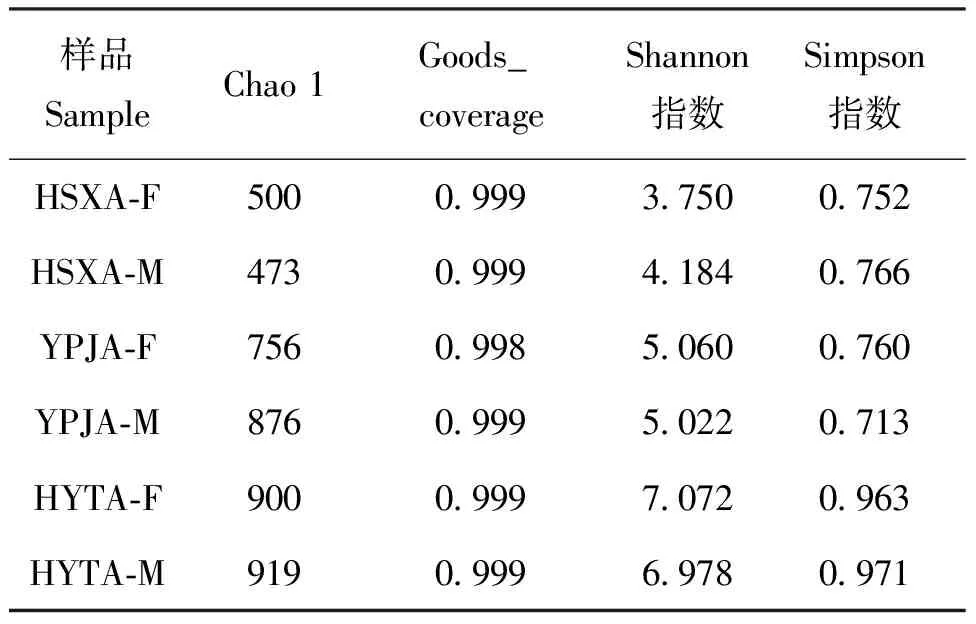
表4 黄脊竹蝗不同地理种群肠道微生物Alpha多样性分析
2.4 黄脊竹蝗肠道微生物Beta多样性分析
为探究不同地理分布的黄脊竹蝗肠道微生物组成差别,本研究根据所有样本在属水平的物种注释及丰度信息,选取丰度排名前35的属,绘制了物种丰度聚类热图。结果如图2所示:从性别来看,3个地区雌雄样品的菌群组成都具有显著差异;从不同地理位置来看,几乎没有3个地理品种共有且丰度较高的菌属,这说明不同地理种群的黄脊竹蝗肠道微生物组成具有明显差异。
为了更直观的对不同地理分布黄脊竹蝗肠道微生物的差异进行聚类分析,本研究采用了NMDS方法。该方法根据样本中包含的物种信息,以点的形式反映在多维空间上,反映样本的组间和组内差异,有效克服线性模型的缺点,更好地反映生态学数据的非线性结构(Rivasetal., 2013)。结果显示(图3-A),不同地理种群的黄脊竹蝗肠道微生物差异较大,其中差异最大的两组为湖南省邵阳市新宁县雌虫(HSXA-F)和云南省普洱市江城县雌虫(YPJA-F)。
进一步深入分析,采用LEfSe分析法(图3-B)。该分析方法可用于进行两个或多个分组的比较,强调统计意义和生物相关性,能够在组与组之间寻找具有统计学差异的Biomarker。结果显示湖南省邵阳市新宁县雌虫(HSXA-F)组中的o_Pseudomonadales、g_Acinetobacter、f_Moraxellaceae等细菌的丰度显著高于云南省普洱市江城县雌虫(YPJA-F),而云南省普洱市江城县雌虫(YPJA-F)中的g_Fusobacterium丰度显著高于湖南省邵阳市新宁县雌虫(HSXA-F)。
2.5 地理环境因子与黄脊竹蝗肠道微生物结构的相关性分析
地理位置的差异会导致环境因子不同,环境因子与蝗虫的生长发育息息相关。为研究不同地理位置的环境因素与黄脊竹蝗肠道菌群之间的关系,采用Spearman相关性分析方法对其进行研究。结果如图4,α多样性中的Chao 1、Dominance、Observed_otus、Pielou_e、Shannon、Simpson指数都与环境因子的蒸发量显著相关,且Dominance和Simpson指数与相对湿度和平均气压显著相关,说明这3个环境因子都影响了黄脊竹蝗肠道菌群多样性。
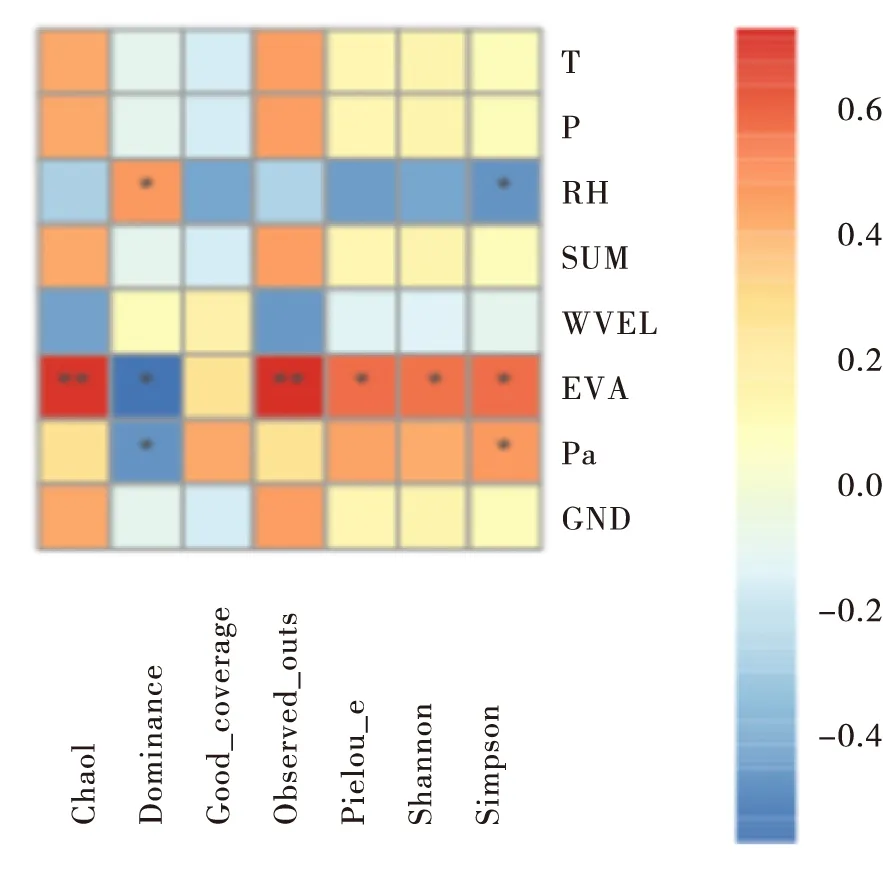
图4 不同地理种群黄脊竹蝗的环境因子分析Fig.4 Analysis of environmental factors of Ceracriskiangsu in different geographical populations
在分类阶元门水平上(图5),平均气温、降水量、日照时数、风速、蒸发量和地面温度与黄脊竹蝗肠道微生物中的梭杆菌门、蓝细菌门、放线菌门、谜古菌门Aenigmarchaeota、变形菌门、Desulfobacterota、广古菌门Euryarchaeota、蛭弧菌门Bdellovibrionota、髌骨细菌门Patescibacteria显著相关。此外,蒸发量还与变形菌门、绿弯菌门、疣微菌门Verrucomicrobiota和芽单胞菌门Gemmatimonadota显著相关。

图5 不同地理种群黄脊竹蝗的环境因子分析(门水平)Fig.5 Analysis of environmental factors of Ceracris kiangsu in different geographical populations (Phylum level)
在分类阶元纲水平上(图6),平均气温、降水量、日照时数、风速、蒸发量和地面温度与黄脊竹蝗肠道微生物中的放线菌纲、蓝藻纲、酸微菌纲Acidimicrobiia、甲烷杆菌纲Methanobacteria、Desulfovibrionia、脱硫单胞菌纲Desulfuromonadia显著相关。平均气温、降水量、日照时数、风速和地面温度与芽孢杆菌纲、厚壁菌纲Negativicutes、Oligoflexia显著相关。此外,蒸发量还与γ-变形菌纲、疣微菌纲Verrucomicrobiae、Saccharimonadia、芽单胞菌纲Gemmatimonadetes显著相关。

图6 不同地理种群黄脊竹蝗的环境因子分析(纲水平)Fig.6 Analysis of environmental factors of Ceracris kiangsu in different geographical populations (Class level)
在分类阶元目水平上(图7),平均气温、降水量、日照时数、风速和地面温度与黄脊竹蝗肠道微生物中的假单胞菌目、乳杆菌目、虫原体目、鞘脂单胞菌目Sphingomonadales、Peptostreptocoocale. Tissierellales、梭杆菌目Fusobacteriales、梭菌目Clostridiales、Veillonellales. Selenomonadales、假诺卡氏菌目Pseudonocardiales显著相关。相对湿度与乳杆菌目、Solirubrobacterales和梭杆菌目显著相关。而蒸发量与假单胞菌目、肠杆菌目、微球菌目、根瘤菌目、弗兰克氏菌目Frankiales、立克次体目Rickettsiales、丙酸杆菌目Propionibacteriales、醋杆菌目Acetobacterales、Solirubrobacterales、假诺卡氏菌目Pseudonocardiales显著相关。平均气压与乳杆菌目、Solirubrobacterales、梭杆菌目显著相关。

图7 不同地理种群黄脊竹蝗的环境因子分析(目水平)Fig.7 Analysis of environmental factors of Ceracris kiangsu in different geographical populations (Order level)Note:V.S.,Veillonellales. Selenomonadales;R.T.,Peptostreptocoocale. Tissierellales。
在分类阶元科水平上(图8),平均气温、降水量、日照时数、风速和地面温度与黄脊竹蝗肠道微生物中的肠球菌科、Streptoooccaceae、螺原体科、拜叶林克氏菌科Beijerinckiaceae、鞘脂单胞菌科Sphingomonadaceae、分支杆菌科Mycobacteriaceae、消化链球菌科Peptostreptocoocaceae、梭杆菌科Fusobacteriaceae、假单胞菌科Pseudomonadaceae显著相关。相对湿度与梭杆菌科Fusobacteriaceae、链球菌科、肠球菌科显著相关。蒸发量与根瘤菌科Rhizobiaceae、Microbacteriaceae、拜叶林克氏菌科Beijerinckiaceae、分支杆菌科Mycobacteriaceae、Mitochondria、德沃氏菌科Devosiaceae、假单胞菌科、类诺卡氏科Nocardioidaceae和醋杆菌科Acetobacteraceae显著相关。平均气压与肠球菌科、链球菌科和梭杆菌科显著相关。

图8 不同地理种群黄脊竹蝗的环境因子分析(科水平)Fig.8 Analysis of environmental factors of Ceracris kiangsu in different geographical populations (Family level)
在分类阶元属水平上(图9),平均气温、降水量、日照时数、风速和地面温度与黄脊竹蝗肠道微生物中的乳球菌属、肠球菌属、螺原体属、短小杆菌属、Peptoclostridium、Allorhizobium.Neorhizobium.Pararhizobium.Rhizobium、不动杆菌属、Mycobactenium、慢生根瘤菌属Bradyrhizobium、假单胞菌属Pseudomonas、Methylobacterium.Methylorubrum、Burkholderia.Caballeronia.Paraburkholderia、Lysinibacillus、梭杆菌属Fusobacterium显著相关。相对湿度和平均气压与乳球菌属、肠球菌属和梭杆菌属显著相关。蒸发量与短小杆菌属、类诺卡氏属Nocardioides、亮杆菌属Leucobacter、慢生根瘤菌属Bradyrhizobium、Allorhizobium.Neorhizobium.Pararhizobium.Rhizobium、假单胞菌属、Methylobacterium.Methylorubrum、Mitochondria、德沃斯氏菌属Devosia显著相关。
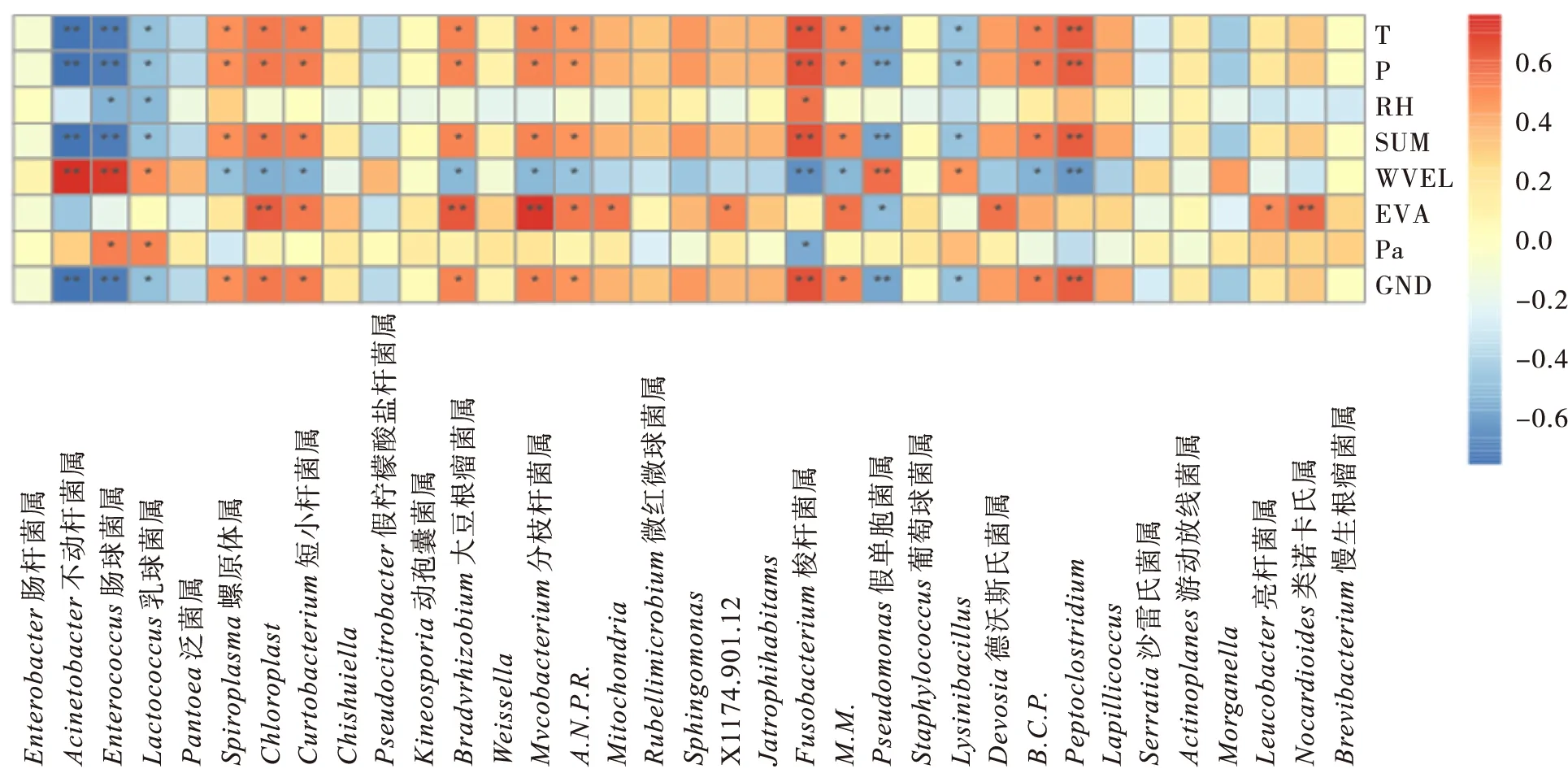
图9 不同地理种群黄脊竹蝗的环境因子分析(属水平)Fig.9 Analysis of environmental factors of Ceracris kiangsu in different geographical populations (Genus level)Note:B.C.P.,Burkholderia. Caballeronia. Paraburkholderia;M.M.,Methylobacterium. Methylorubrum;A.N.P.R.,Allorhizobium. Neorhizobium. Pararhizobium. Rhizobium.
3 结论与讨论
2020年6月黄脊竹蝗从老挝和越南等地迁飞进入我国云南边境地区,由于种群数量激增,取食范围扩展至大田作物,波及范围9 000 ha,其中农田约2 133 ha,林地约6 866 ha,对云南边境地区农业生产造成严重危害,引起社会广泛关注(李永华和刘勇波, 2021; 赵紫华等, 2021; 卓富彦等, 2021)。
蝗虫的群居型特征是造成蝗虫数量暴发成灾的特征之一,蝗虫肠道微生物与聚集信息素的合成与释放密切相关。通过分析昆虫肠道微生物的多样性及外在条件刺激下的菌群结构及动态变化,可以进一步了解宿主昆虫和肠道微生物间的关系(杭苏琴, 2007),对于昆虫的生命活动和生理代谢具有重要意义。关于蝗虫肠道微生物多样性的研究,近年来也可见一些报道。Dillon对沙漠蝗SchistocercagregariaForskal肠道微生物在饥饿和虫龄两个影响因子作用下的多样性进行分析,结果显示细菌多样性与蝗虫年龄成正比,而受饥饿的蝗虫通常容易发生疾病(Dillonetal., 2010);Su等人通过研究16种蝗虫的肠道共生菌,发现纤维素分解酶和肠道微生物群落可能反映了不同种类的蝗虫与其摄食方式的联系(Suetal., 2014);徐冲等人从东亚飞蝗LocustamigratoriamanilensisMeyen肠道内分离出12株产纤维素酶细菌,并对其中一株酶活性较高的菌株进行一系列实验,鉴定为蜡样芽胞杆菌(徐冲等, 2016)。
本研究通过分析我国新入侵地区和国内常发地区的黄脊竹蝗肠道微生物多样性,为研发微生物制剂和综合防控提供理论依据。研究发现,对于黄脊竹蝗来说,不同地理分布和性别的肠道内微生物多样性与丰富度不同。其中,地理位置影响最为显著,其次为性别因素。对比不同分布区域的黄脊竹蝗肠道微生物多样性,结果显示湖南省邵阳市新宁县和云南省普洱市江城县的差异最大。2020年黄脊竹蝗从境外迁入我国云南境内,由于该地气候环境适宜且食物充足,造成该虫大量繁殖危害(卓富彦等, 2020)。推测此次在云南省普洱市江城县采集到的黄脊竹蝗应该来自于境外迁入,因而两地的黄脊竹蝗肠道微生物多样性差异最大。通过对比不同性别的黄脊竹蝗,结果显示雌雄蝗虫的肠道菌群在属水平组成上具有显著差异。有研究表明不同性别的昆虫体内微生物多样性与丰度存在差异,例如中华通草蛉ChrysopasinicaTjeder雌雄群体间的肠道微生物在属水平组成存在差异,该研究推测差异原因是由于雄雌成虫生理代谢的不同引起微生物菌群的差异(赵辉等, 2017)。目前仅有研究表明黄脊竹蝗食叶量随虫龄的增长而增加,雌蝗的食叶量远远大于雄蝗(练佑明等, 1995),有关黄脊竹蝗雌雄个体肠道微生物差异性机理还少有研究。
研究表明,不同的环境条件会影响栖居于昆虫肠道内的共生微生物与其宿主之间的相互作用,进而微生物的种类及分布也会受到影响(Schmidetal., 2015;Donkersleyetal., 2017)。本研究所选取的8个环境因子中对黄脊竹蝗肠道微生物影响最大的为蒸发量、相对湿度和平均气压。以往研究发现,尽管不同种类的昆虫肠道菌优势门存在差异,但变形菌门均是最主要的肠道细菌群体之一(Yunetal., 2014),本研究中关于黄脊竹蝗优势菌门的分析也支持上述结果。
近年来,由于环境气候不断变化,黄脊竹蝗入侵频率逐年增加,防控形势依然严峻(卓富彦等, 2020)。蝗虫的群居觅食活动给农业生产带来巨大的破坏,有研究表明蝗虫粪球中所含的芳香族化合物聚集信息素是维持昆虫群体聚集的关键因素之一,而肠道微生物的次生代谢产物愈创木酚和少量苯酚是合成聚集信息素的必要组分(Dillonetal., 2010)。因此,通过进行蝗虫肠道微生物的相关性研究,可为害虫生物防治提供理论指导。作为生态系统中的因素,蝗虫还将长期存在,人类与蝗虫争夺粮食和牧草等斗争也将是长期的。因此,未来仍不能放松警惕,要彻底贯彻“改治并举”的治蝗工作方针,将治蝗准则牢记于心,努力实现“虫口夺粮”。
猜你喜欢
小主人报(2023年1期)2023-03-15
小天使·六年级语数英综合(2022年5期)2022-05-27
小学生优秀作文(低年级)(2022年3期)2022-03-29
矿产勘查(2020年11期)2020-12-25
音乐教育与创作(2019年10期)2019-12-26
国际呼吸杂志(2019年1期)2019-01-28
小布老虎(2017年6期)2017-07-31
现代检验医学杂志(2016年2期)2016-11-14
环境科技(2016年3期)2016-11-08
发明与创新(2016年30期)2016-08-22
