
两种二倍体野生花生ABI3/VP1的全基因组鉴定与分析
2023-10-27 06:36王江丽张慧哲王希胤
华北理工大学学报(自然科学版) 2023年4期
王江丽,张慧哲,王希胤
(1. 华北理工大学 生命科学学院,河北 唐山 063210;2. 华北理工大学 基因组学与计算生物学研究中心,河北 唐山 063210)
引言
转录因子(Transcription factors, TFs)是一种序列特异性的蛋白,通过调控靶基因,在植物生长、发育、激素反应以及生物和非生物胁迫应答中起着重要作用[1]。与ABI3/VP1转录因子家族相关的RAV转录因子,编码一个B3结构域和一个APETALA2(AP2)结构域,属于APETALA2/乙烯响应元件结合因子(AP2/ERF)或B3超家族,在调节种子萌发、植物生长发育和响应生物或非生物胁迫中发挥重要作用[2-4]。首先在玉米中发现ABI3/VP1家族成员中的VP1基因,随后在模式植物拟南芥中也鉴定出其同源基因[5]。后来,为分析RAVs在植物生长发育过程中的作用,在苜蓿、大麦、蓖麻等许多粮食作物中做了一些研究[6-8]。比如在苜蓿中ABI3/VP1相关基因的表达能够导致拟南芥抗逆性增强和分枝增加,其研究结果为MtRAVs在豆科植物中的应用奠定了基础[6]。
花生是豆科重要的模式物种,也是世界上重要的粮食作物之一,对花生基因组内部各个功能基因的研究对提升其产量和品质具有重要意义[9-11]。研究表明,2个野生二倍体花生Arachis duranensis(后续缩写为Adu)和Arachis ipaensis(后续缩写为Aip)是所有栽培花生的二倍体祖先,其基因组测序的完成对花生相关生物信息学分析奠定了基础[12]。花生作为种植面积最大、产量最高的油料作物之一,其种植面积在国内非常广泛,而河北省是北方地区花生种植面积较大的省份之一[13]。花生不仅是一种营养价值高的食物,而且其药用价值较高,花生籽、种皮、种壳和花生油等均可入药。除了其营养上的重要性,花生也是一种在遗传学、功能基因组学和育种方面被许多作物领域研究人员所深入研究的植物。花生功能基因的挖掘是分子育种工作的关键[14-17],许多转录因子家族的基因鉴定和表达分析能够揭示花生在种子休眠或种子萌发过程中的关键作用,为花生品质和产量的提高提供参考[18-20]。
目前对于花生ABI3/VP1转录因子家族的相关研究还未见报道。本研究将对花生ABI3/VP1转录因子家族进行全基因组鉴定,并通过一系列生物信息学方法对其进行系统发育、理化性质,二级结构和亚细胞定位,启动子顺式作用元件分析,为后续开展花生ABI3/VP1基因调控脱落酸信号转导相关的研究提供参考。
1 材料与方法
1.1 二倍体野生花生ABI3/VP1的全基因组鉴定
从拟南芥基因组数据库(https://www.arabidopsis.org/)下载ABI3/VP1 转录因子家族的相关基因ID及其对应的cds与pep序列数据,从花生基因组数据库(https://www.peanutbase.org/)下载2个二倍体花生Adu及Aip的全基因组数据。利用拟南芥的ABI3/VP1基因的蛋白序列文件,使用BLASTP[21]搜索工具进行同源序列比对,得到花生的ABI3/VP1基因的蛋白序列文件。同时,在SMART网站(http://smart.embl-heidelberg.de/)对蛋白序列针对基因家族特有结构域进行筛选,在NCBI(https://www.ncbi.nlm.nih.gov/)网站对CDD结构域进行分析并利用TBtools软件[22]进行可视化分析得到候选基因的结构域情况,去除不完整结构域基因后得到最终花生ABI3/VP1的基因。
1.2 系统发育进化树的构建和基因结构分析
利用ClustalW[23]工具将鉴定出的蛋白序列进行多重比对,使用MEGA-X软件[24]构建系统发育进化树并用在线软件iTOL(https://itol.embl.de/)进行美化。利用TBTools软件展示ABI3/VP1家族基因结构。
1.3 花生ABI3/VP1编码蛋白的二级结构分析和亚细胞定位
利用SOPMA(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)和 PSORT在线网站(http://psort.hgc.jp/)分别预测花生ABI3/VP1编码蛋白的二级结构和亚细胞定位。
1.4 花生ABI3/VP1基因染色体定位和顺式作用元件分析
利用TBTools分析获得花生ABI3/VP1基因在染色体上的物理位置,并绘制其在染色体上的分布图。利用PlantCARE在(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)进行启动子顺式作用元件的预测并将结果整理。
2 结果与分析
2.1 二倍体野生花生ABI3/VP1的全基因组鉴定
利用拟南芥ABI3/VP1基因家族的11个蛋白序列建库,通过Blastp搜索同源序列,在2个花生Adu和Aip基因组中鉴定出18个ABI3/VP1候选基因(见表1)。同时将拟南芥的11个已鉴定基因与其所具有的功能制表,并在基因名称后添加相应功能注释(见表2)。为了检验ABI3/VP1候选基因结构域的完整性,进一步利用SMART、TBtools进行结构域搜索,发现花生ABI3/VP1候选基因Ad_01326结构域片段不完整,仅含有22个氨基酸残基,而它与拟南芥该基因家族的blast结果中相应匹配的基因为AT4G30080,该基因在该家族中并不具有重要作用,推测ABI3/VP1候选基因Ad_01326可能是假基因(见图1),不对它进行分析。在剩下的17个基因中,发现其全部位于花生2个种的2、4、5、6、7、8、9号染色体上,且大部分位于2、5、6和9号染色体。另外有7个基因位于正向链,10个位于负向链(见表3)。
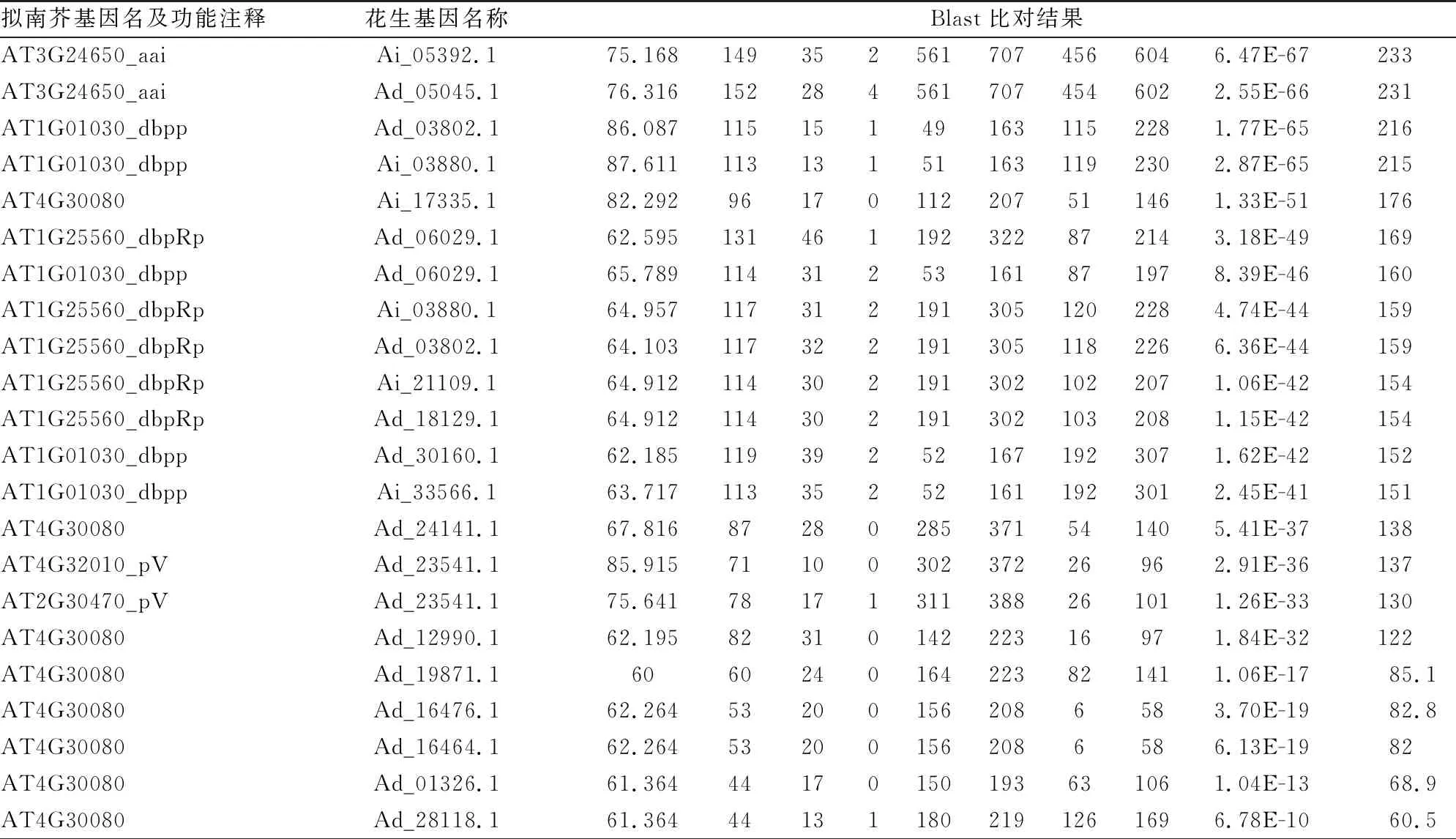
表1 拟南芥ABI3/VP1基因与二倍体野生花生blast比对结果

表2 拟南芥根据已知的功能修改名称
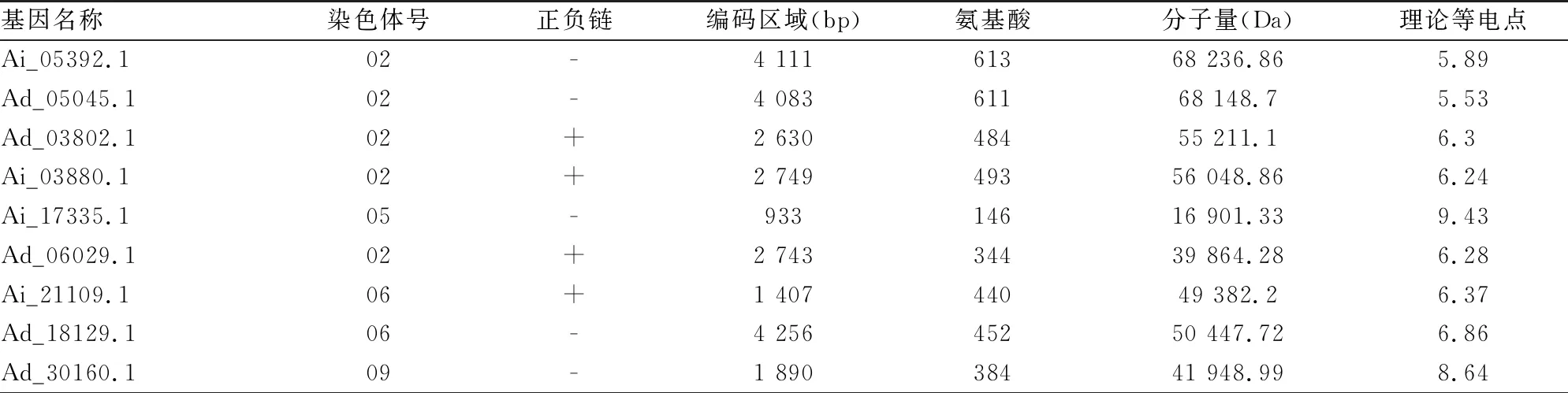
表3 花生ABI3/VP1基因家族成员信息
2.2 系统发育进化树的构建和基因结构分析
利用拟南芥和花生共28个ABI3/VP1基因编码的蛋白质氨基酸序列构建其ABI3/VP1基因家族进化树(见图2)。根据进化树拓扑结构,可将ABI3/VP1蛋白分为3个亚家族。在3个亚家族中均包含拟南芥和花生的ABI3/VP1基因,同时在进化树拓扑结构中发现,花生ABI3/VP1基因在现代支与中间支中分布很少,一共只有 4个ABI3/VP1基因,其余 13个ABI3/VP1基因均位于古老支。为了进一步研究花生ABI3/VP1基因结构的多样性和系统发生情况,对17个ABI3/VP1基因的蛋白质氨基酸序列单独构建系统发育进化树(见图3),结果也分为3个亚家族(分类方式同上)。对这些蛋白质的保守基序分析发现(见图4),共有20种motif被分析出来,其中motif 1、2、3、5、6、16、20在花生ABI3/VP1基因家族中广泛存在,Ad_16476.1、Ad_12990.1、Ad_19871.1、Ad_16464.1中仅有motif 1的存在,Ad_23541.1和Ad_24141.1中没有检测到motif的存在。古老支的特有基序有motif3、motif4、motif8、motif14、motif15、motif16、motif17,中间支特有基序有motif10和motif12。通过比较花生ABI3/VP1基因结构发现,除Ad_30160.1没有内含子外,其余ABI3/VP1基因均含有1~7个内含子。

图2 花生与拟南芥ABI3/VP1基因家族的系统进化树
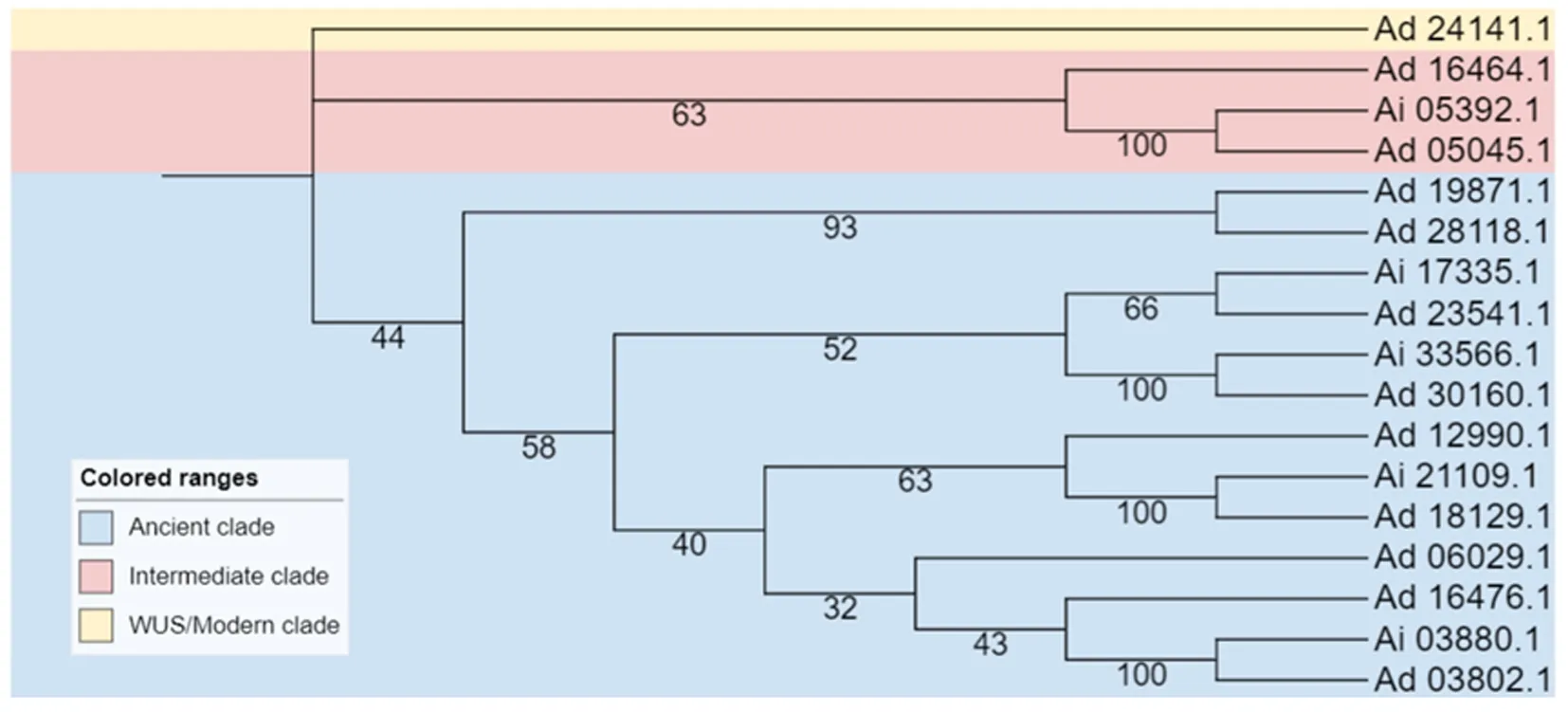
图3 花生ABI3/VP1基因家族的系统进化树

图4 花生ABI3/VP1基因家族的系统进化和基因结构
2.3 花生ABI3/VP1编码蛋白的二级结构分析和亚细胞定位
二级结构分析显示(见表4),花生17个ABI3/VP1基因的氨基酸序列均含有α-螺旋、延伸链、β-转角和无规则卷曲,其含量分布范围有所不同。如α-螺旋和随机卷曲在ABI3/VP1基因编码蛋白质二级结构中的含量范围分别为8.43%~40.27%、28.32%~66.81%,而拓展链和β-转角的含量范围分别为11.62%~34.85%、2.88%~15.15%,以上表明17个ABI3/VP1基因氨基酸序列二级结构主要以α-螺旋和随机卷曲为主。同时,亚细胞定位预测显示花生ABI3/VP1基因有11个定位在细胞核中,2个定位在叶绿体中,4个定位在细胞质中。

表4 花生ABI3/VP1基因编码蛋白的二级结构及亚细胞定位
2.4 花生ABI3/VP1基因染色体定位和顺式作用元件分析
染色体定位分析结果显示(见图5),花生的17个ABI3/VP1基因在11条染色体上呈不均匀分布,其中Ad种的12个基因分别位于2、4、5、6、7、8、9号共7条染色体, Ai种的5个基因位于2、5、6、9号共4条染色体。
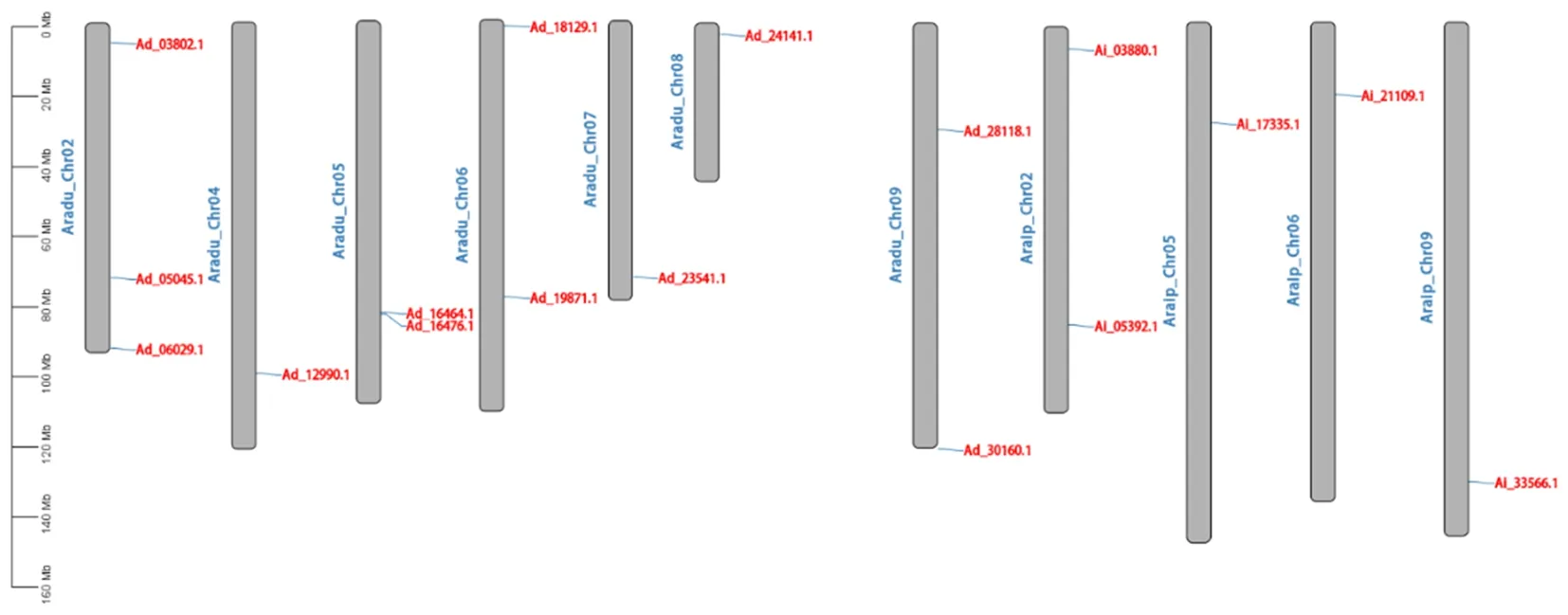
图5 花生ABI3/VP1基因的染色体定位
对花生17个ABI3/VP1基因启动子序列的顺式作用元件的预测结果显示,共有21种397个顺式作用元件,且多种顺式作用元件与植物生长过程中的相关激素应答、组织特异性表达等密切相关(见图6)。其中,光照响应元件在花生的17个ABI3/VP1基因中都存在,暗示花生ABI3/VP1基因表达可能受光照影响。
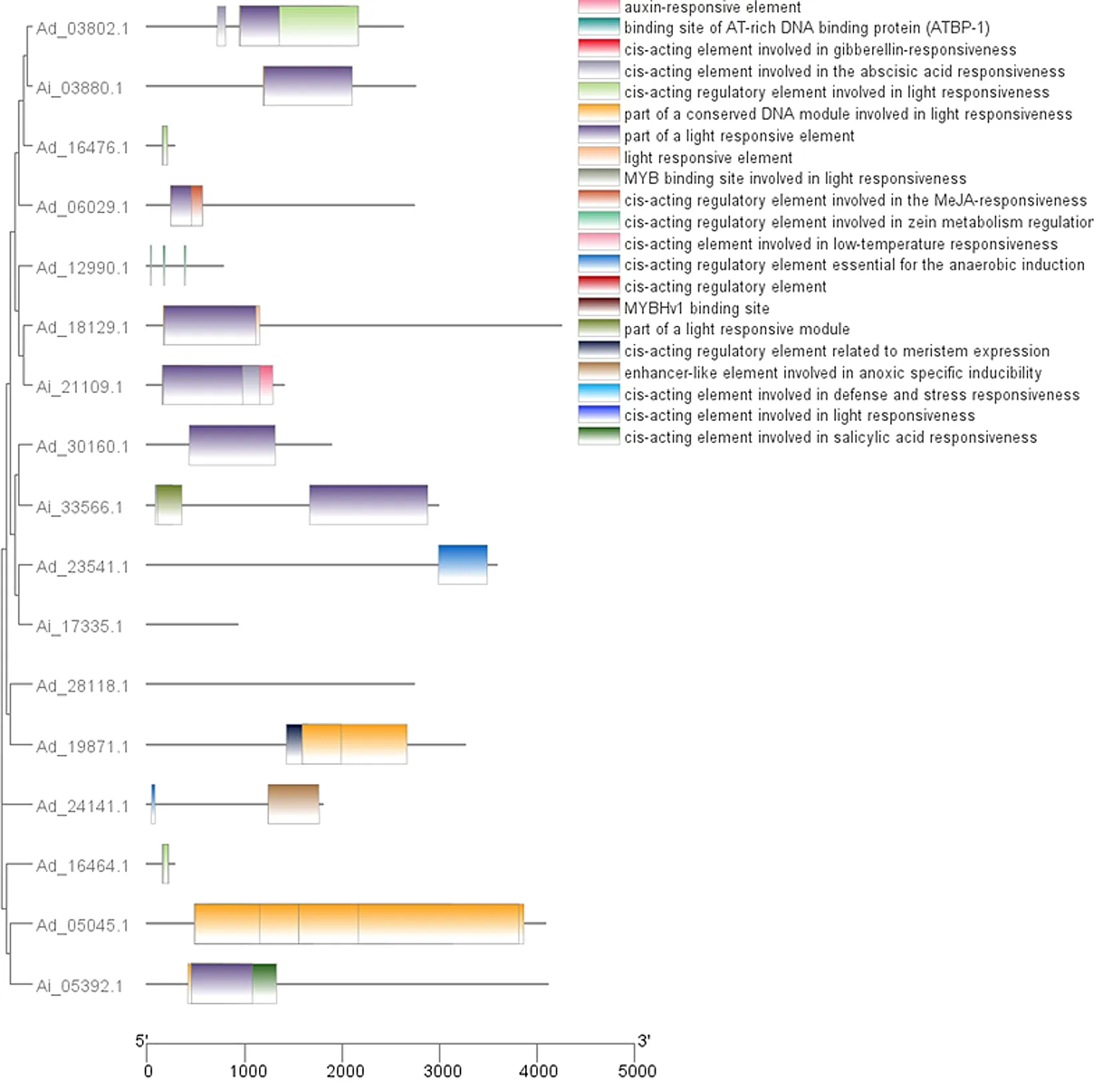
图6 花生ABI3/VP1基因的顺式作用元件分析
3 结论
(1)研究从花生基因组中成功鉴定出17个ABI3/VP1基因,并且不均匀地分布在11条染色体上,相比于作为十字花科的拟南芥具有的11个而言,豆科的花生具有了较多的基因,这可能是在两者祖先物种分歧时发生的基因丢失或加倍导致。
(2)系统发育表明,花生古老支中有较多基因与拟南芥亲缘关系较近,并且在观察花生2个不同种在3个亚家族中的分布后发现,在古老的分支中Adu和Aip 2个种的基因数量并无差别,而随着时间的推移,在中间支和现代支Aip的该家族基因数量明显小于Adu的数量,推测为地缘差异或者种间相关基因的差异造成。值得注意的是,Aip的该家族相关基因从中间支到现代支经历了从无到有的变化,推测与基因的丢失或古老基因的加倍与进化有关。综合理化性质、亚细胞定位及顺式作用元件分析,推测花生ABI3/VP1基因参与多个信号通路调控花生的生长发育过程和对环境刺激的响应。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17
天津市教科院学报(2021年5期)2021-11-10
生物学通报(2021年9期)2021-07-01
广州大学学报(自然科学版)(2019年1期)2019-05-07
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年3期)2017-04-10
江苏农业科学(2016年8期)2017-02-15
山东农业工程学院学报(2016年6期)2016-12-01
天津科技大学学报(2016年1期)2016-02-28
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10
