
冻融作用对黄河源区曲流河岸土体抗剪特性的影响
2023-12-04 08:03刘国松朱海丽刘亚斌李国荣
干旱区研究 2023年10期
刘国松, 朱海丽,2, 张 玉, 刘亚斌,2, 李国荣,2
(1.青藏高原北缘新生代资源环境重点实验室,青海 西宁 810016;2.青海大学地质工程系,青海 西宁 810016)
黄河源区冻土的分布孕育了源区大面积高寒沼泽湿地和高寒草甸,其间河网密布,湖泊和湿地较多,成为青藏高原重要的水源涵养地。近几十年来,黄河源区高寒草甸在自然气候条件和人为活动的双重影响下,冻土环境发生着显著变化,导致草地退化,高寒草甸生态屏障作用削弱,进而导致河岸稳定性降低、河岸崩塌频繁发生[1]。由于河岸带根土体的力学特性是影响河岸稳定的关键因素,因此,明确冻融作用下源区河岸带高寒草甸土体力学特性的变化规律对定量分析河岸稳定性、解释河岸崩退机制并预测曲流横向演变趋势具有重要意义。
冻融循环对土体力学性能的影响主要是由于土体的三相介质在强烈的冻结和融化过程中具有不同的热物理特性造成的[2-3]。不同土体结构和含水条件的土体在冻融循环作用下的冻胀变化存在明显差异[4],当冻胀力超过一定数值,则会改变土颗粒之间的排列和连结,进而在宏观上表现为土体的剪切强度、弹性模量和黏聚力等力学性能指标降低[5-7],而土体内摩擦角变化规律不一致[8-12]。当土体内加入纤维或根系等材料时,其对土体的力学性能有较大的影响,能显著提升土体黏聚力。申紫雁等[13]、杨馥铖等[14]通过直剪试验研究提出含根土较素土黏聚力提高2.47%~94.1%;Mahya 等[15]、Orakoglu 等[16]通过对掺入纤维材料的土体开展冻融循环前后的剪切试验,发现随冻融循环次数增加,加筋土体强度衰减幅度较素土减弱;Gong 等[17]通过研究冻融循环作用下聚丙烯纤维对路堤边坡土体稳定性的影响,提出纤维的加入提高了冻融循环作用下土体的黏聚力,改善了土体稳定性,并随着冻融循环次数的增加,其改善作用降低;王恒星等[18]研究了结缕草(Zoysia japonica)、牛筋草(Eleusine indica)、狗尾草(Setaria viridis)的根-土复合体在冻融前后的抗剪特性,提出冻融循环之后根-土复合体黏聚力损失较大,损失幅度在28.6%~55.0%之间,但其黏聚力仍高于素土,内摩擦角先增大后减小。黄河源区曲流河岸土体力学特性,既受到季节性冻融作用影响,又存在滨河草甸植物根系作用影响,且随着高寒草甸植被的退化,其地下有效根系减少[19],分析二者共同作用下,高寒草甸土体抗剪力学特性的研究相对较少。因此,本文以黄河源区曲流河岸高寒草甸含根土体为研究对象,通过室内重塑冻融直剪试验深入分析不同冻融循环作用、不同含根量变化对滨河土体抗剪强度指标的影响,为评价黄河源区河岸带不同退化程度的高寒草甸植被根系护岸作用提供数据支撑,同时为分析区内曲流河岸稳定性及其横向演变趋势提供理论依据。
1 研究区概况及研究方法
1.1 研究区概况
选定黄河源二级支流兰木措曲作为研究区,其位于青海省黄南蒙古族藏族自治州河南县,地理坐标为34°26′~35°02′N,101°23′~101°35′E(图1),属于典型的高寒草甸型弯曲河流,其流程长,流域大,水文地质稳定,水力蕴藏量十分丰富。试验区位于兰木错曲中游,该段河床主要为砾卵石河床,土壤类型主要为高山草甸土及沼泽化草甸土。研究区内海拔为3400~4200 m,受季风的影响,高原大陆性气候特点明显,冷季较长且多大风。根据青海省河南 县 气 象 站1981 年7 月1 日—2018 年6 月30 日 气温、降水、日照、冻土等观测资料可知,多年平均降水量为560 mm,多年平均蒸发量为1349 mm,年平均气温0.4 ℃,属于季节性冻土区,年平均冻土深度为137.8 cm,冻土深度最深为162 cm,5—6 月为消融期,11 月至翌年4 月为冻胀期。研究区内河岸带植被以高寒草甸为主,其中未退化草甸群落以华扁穗草(Blysmus sinocompressus)、线叶嵩草(Kobresia capillifolia)和垂穗披碱草(Elymus nutans)等草本植物为优势植物种;随着河岸带草甸植被退化,优势植物种减少,鹅绒委陵菜(Potentilla anserina)、美丽风毛菊(Saussurea superba)、矮小火绒草(Leontopodium pusillum)等典型退化物种增多,地下根量减少。
图1 研究区位置示意图Fig.1 Location map of the study area
1.2 试验材料
为了给室内重塑冻融试验提供物理参数及根系参数设定依据,在研究区随机选取3 个河湾凹岸做为试验和取样点位。由于研究区高寒草甸植物根系主要集中于地表以下0~30 cm 深度范围,且含根土体密度、含水率均随土层深度增大而变化,因此将原位取样的深度确定为0~30 cm,分三层取样(0~10 cm、11~20 cm 和21~30 cm)、同一深度范围重复3 次取样并综合求其平均值,最终设定室内重塑根-土复合体的密度、含水率及根系含量,使其数值具有代表性。其中,土体含水率采用小铝盒取样,烘干法测定;密度采用小环刀(内径6.18 cm,高2 cm)原位制样,称重法测定;原状复合根-土体采用大环刀(内径10 cm,高6.37 cm)取样,室内采用洗根法分析单位干土中的根系质量。
试验所用的土体材料取自研究区取样点滨河凹岸地表以下0~30 cm 深度范围的土体,土质类别为粉土[20];重塑根-土复合体制样选用的根系材料为研究区优势滨河植物华扁穗草根系。结合课题组前期野外调查和原位植被样方试验结果[20],研究区河岸带华扁穗草分布广泛、覆盖度达60%~90%,因此,将其根系作为重塑根-土复合体根系材料。利用现场挖掘法在华扁穗草生长正常的区域进行根系取样,挖取花盆大小的根-土复合体土柱,将其装入花盆并带回实验室。回实验室后,采用浸泡、冲洗的方法,获得干净的植物根系,放入恒温箱4 ℃保存待用。
1.3 试验设计
试验得知,研究区滨河土体的密度范围在1.34~1.78 g·cm-3之间、含水量在37.2%~53.4%之间,地表植被覆盖度不同,其物理参数也相应变化。取实测土体物理参数的平均值,设定根-土复合体密度为1.55 g·cm-3,含水量为45%。统计大环刀内根系直径及含根量(植物根系质量与干土质量之比)可知,试样中根系平均直径为0.11~1.69 mm,根系含量范围为4.1%~11.7%。基于实测根系参数,试验按照逐级增加的方式,含根量设置为0%、4%、7%、11%4个梯度,试验所用根系材料平均根径为0.58 mm,根长为30 mm。结合研究区近10 a每年10月至次年4月的气温和日照时长等气象数据,将本试验冻结温度和融化温度分别设定为-18 ℃和10 ℃,冻、融时长各为12 h,完成一次冻融24 h。试验设置冻融循环次数分别为0、3、6、9、12、15、20次,7个试验梯度。
1.4 试样制备及直剪试验方法
根据试验设计进行重塑直剪试样的制备,制取过程根据规范《土工试验规程》进行。具体操作过程如下:(1)将野外所取得的土体风干碾碎后过2 mm 筛;(2)在获得的干净华扁穗草根系中挑选出根径为0.58±0.05 mm 的根系,并将其剪至长约为30 mm 左右的根段;(3)首先按设计的含根量梯度将剪好的根段与素土干土充分拌匀,然后按设计的含水率和密度配置素土及不同含根量的根-土复合体试样,盖上保鲜膜后静置12 h;(4)待根-土复合体完全浸透,再次搅拌均匀,按质量将其均分为4 份,依次装入三瓣模中,利用分层击实法分4 层击实;(5)将制备好的复合根-土体试样用保鲜膜密封,以减少水分的流失;(6)对试样进行编号后,放入冰箱并在-18 ℃进行冻结,冻结完成取出后,再放置在10 ℃条件的恒温恒湿箱内消融12 h,此过程为一次冻融循环。根据试验设计,依次完成不同含根条件和冻融次数的试样冻融循环试验;(7)对完成既定冻融循环次数后的试样,立即按照《土工试验规程》展开直剪试验。
采用试验室ZJ 型应变控制式直剪仪开展直接剪切试验,剪切速率设定为0.8 mm·min-1,采用逐级加压的方式,每组试样施加的法向应力分别为50 kPa、100 kPa、200 kPa、300 kPa,每组试样试验结束后,记录所得黏聚力c值和内摩擦角φ值。
2 结果与分析
由表1可知,复合根-土体黏聚力在含根量最高(11%)、未经冻融时达到极大值11.9 kPa,在不含根、冻融循环6次时,达到极小值6.37 kPa,其变化幅度较大;内摩擦角在含根量为7%、冻融循环6次时,达到极小值15.53°,其余均在17.22°~20.3°范围内浮动,变化幅度不大。
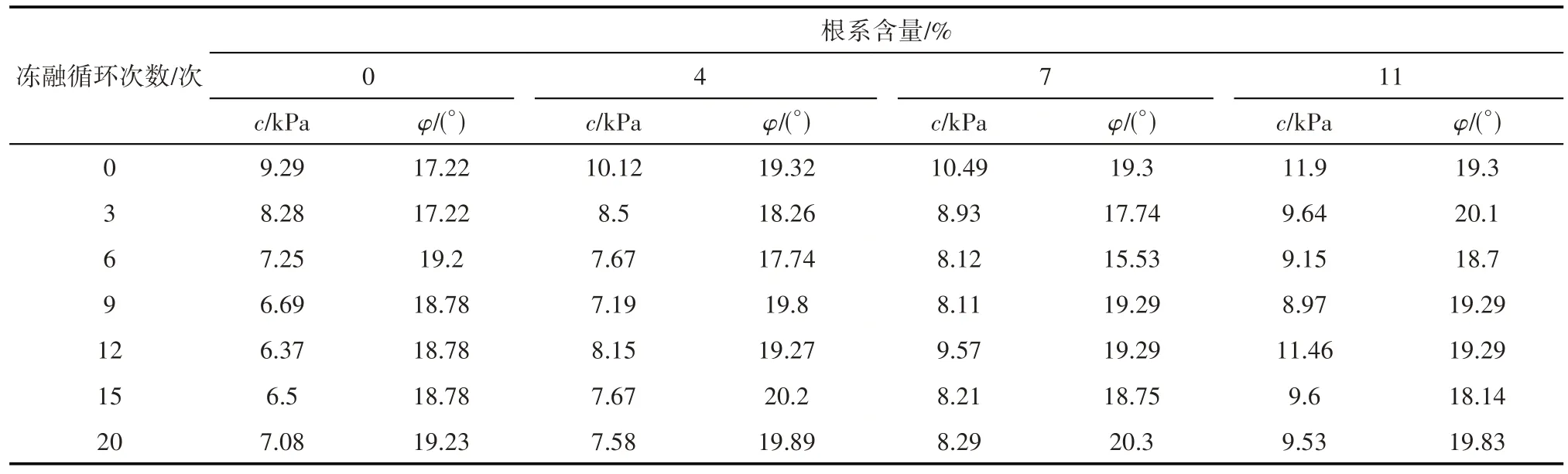
表1 冻融试样抗剪强度指标Tab.1 Shear strength index of remolded freeze-thaw sample
采用SPSS软件对以上数据进行数据整理分析,得到不同含根量、不同冻融循环次数对抗剪强度指标黏聚力和内摩擦角影响的方差分析。其中黏聚力方差分析结果如下,含根量F=34.3,显著性P=0,差异极显著;冻融循环次数F=13.1,显著性P=0,差异极为显著,且决定系数R2=0.91,说明不同含根量、不同冻融循环次数对黏聚力影响极显著。其中,内摩擦角方差分析结果如下,含根量F=1.3,显著性P=0.3,差异性不显著;冻融循环次数F=2.0,显著性P=0.1,差异性不显著,且决定系数R2=0.46,说明不同含根量、不同冻融循环次数对内摩擦角影响不显著。因此,需对不同含根量、不同冻融循环次数对抗剪指标的影响规律及影响机制进行详细探究。
2.1 冻融作用下含根量对根-土复合体抗剪强度指标的影响
由图2可知,不同冻融循环次数作用下,土体黏聚力均随含根量的增加,呈现增大的趋势,增大幅度明显,且不同冻融循环次数的增大幅度有所差异。结合表1 数据可知,黏聚力增加幅度在冻融循环3 次时最低,曲线最缓,相较于不含根土体,含根量为4%、7%、11%的复合根-土体黏聚力增幅依次为2.7%、7.9%、16.4%;黏聚力在冻融循环12 次时增幅最大,曲线最陡,相较于不含根土体,含根量为4%、7%、11%的复合根-土体黏聚力增幅依次达到27.9%、53.0%、77.9%。可以看出,植物根系增强土体黏聚力的效果显著,这主要是由于植被根系穿插在土体中,可将较小颗粒的土组成较大的团聚体,形成具有网络结构的复合根-土体,在剪切受力过程中,植物根系可发挥其抗拉特性以抵抗土体的剪切破坏,同时通过根系的缠绕、加筋作用使土体黏聚力得到较大提升[21],即根系含量高可有效提高根-土复合体黏聚力,表现为未退化和轻度退化的高寒草甸河岸崩退现象不明显。
此外,由图2 还可知,在冻融循环条件下,土体强度相较于未冻融土体会有所降低,但其中的根系能有效减少土体黏聚力降低幅度,与王恒星等[18]得出的结论相一致。表明在根系的作用下,土体间的连结断开,改变了土壤热量的收支、传递,从而对土体间的热量传递起到调节作用。土体内根系可在冻期内延缓热量支出,融期内阻挡热量输入,进而在一定程度发挥阻隔温度变化的作用[22],类似于泡沫阻隔热量的传递。由此,减少冻胀作用对土体结构的破坏,从而表现为含根土体在冻融循环作用下,土体黏聚力下降幅度相对减小。
由图3 可知,土体内摩擦角随含根量的变化规律如下:未冻融、冻融循环9 次、12 次和20 次的变化规律类似,均随含根量的增加内摩擦角呈现先增大、后趋于基本不变的变化特征;冻融循环3次试样的内摩擦角呈现先增大-减小-再增大的变化特征,而冻融循环6 次试样呈现先减小后增大的变化趋势,且可以看出冻融循环6次时,试样内摩擦角变化幅度相较于其他冻融循环次数变化相对较大,当含根量为7%时,相较于不含根土体,内摩擦角下降幅度最大,达到19.1%。总体来看,当含根量为7%、冻融循环达6次时,其内摩擦角显著减小,其余含根量条件的土样在冻融循环作用下其变化幅度都不大,在一定范围内呈波动状态。产生原因可能是由于根系的加入使得部分土-土接触转化为根-土接触,相较于原土体,其土体颗粒间的接触面积以及孔隙大小会有所变化,但由于高寒草甸植物根径较小,平均仅为0.58 mm,因此,其根土接触面积相对较小,从而导致根土间摩擦力较小,表现为内摩擦角有小范围的变化[22]。对比王恒星等[18]以平均半径为0.88 mm 狗尾草为试验材料的冻融直剪试验结果,其内摩擦角呈现较大的变化幅度,可知根系半径会对内摩擦角产生影响。
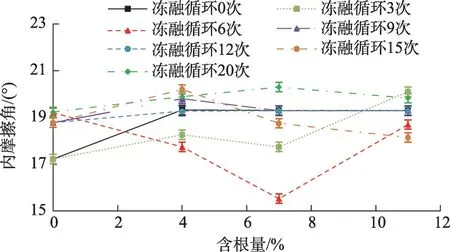
图3 不同冻融循环次数下内摩擦角随含根量变化关系Fig.3 Variation of internal friction angle with root content under different freeze-thaw cycles
2.2 冻融循环次数对根-土复合体抗剪强度指标的影响
为分析冻融循环次数对根-土复合体抗剪强度指标的影响,根据表2 数据绘制土体黏聚力和内摩擦角随冻融循环次数变化关系图(图4、图5)。图4表明,当试样的冻融循环次数由0 次依次增加至3、6、9、12、15、20 次,其黏聚力总体表现出降低的变化趋势。且可以看出,其降低趋势随冻融循环次数增加逐渐变缓,对比4 种含根量土体试样抗剪强度指标的降低趋势,发现4 种不同含根量的试样在冻融循环3次时,试样黏聚力降低幅度均达到最大,分别降低了1.01 kPa、1.62 kPa、1.56 kPa 和2.26 kPa。这主要是因为在冻融初期,试样经过冻融循环,土体冻结过程中,生长的冰晶由于体积膨胀破坏了土颗粒间的联系,而融化时,被破坏的土颗粒间连结不可恢复,因此反复冻融作用使土体结构呈现弱化,土体原有土颗粒的结合形式被破坏,进而导致黏聚力下降[8],而随着冻融循环次数的增加,冻融作用对土体的土颗粒间的影响越来越小,黏聚力降低速度也逐渐变缓[10]。在冻融循环达到12次时,素土试样的黏聚力降到最低,降幅达31.4%;随着冻融循环次数增加,其黏聚力变化趋于平缓;而含根试样在9次冻融循环时,其黏聚力降到最低。含根量为4%、7%、11%的试样,在达到9 次冻融循环时,降低幅度分别为29.0%、22.7%、24.6%,均小于不含根试样的降低幅度。含根试样的黏聚力在冻融循环12 次时会有一个较为明显的回升,其黏聚力较素土试样黏聚力降低幅度要小27.7%,随后降低并趋于平稳。结合苏谦等[5]、左小锋等[23]学者的研究分析,可能是根土体随着冻融循环次数的增加,由于穿插在土体中根系作用,原网状根土结构被破坏并进行重组,土颗粒大小、形状、颗粒间的孔隙及根系与土颗粒间的连结状态在重组过程中逐渐稳定,根-土体结构达到新的平衡状态,并表现为受冻融循环影响变小,根-土体抗剪强度有所增加,土体黏聚力在冻融循环12次时有明显的回升,而后变化趋于平缓。

图4 黏聚力随冻融循环次数变化关系Fig.4 Relationship between cohesion and freeze-thaw cycles
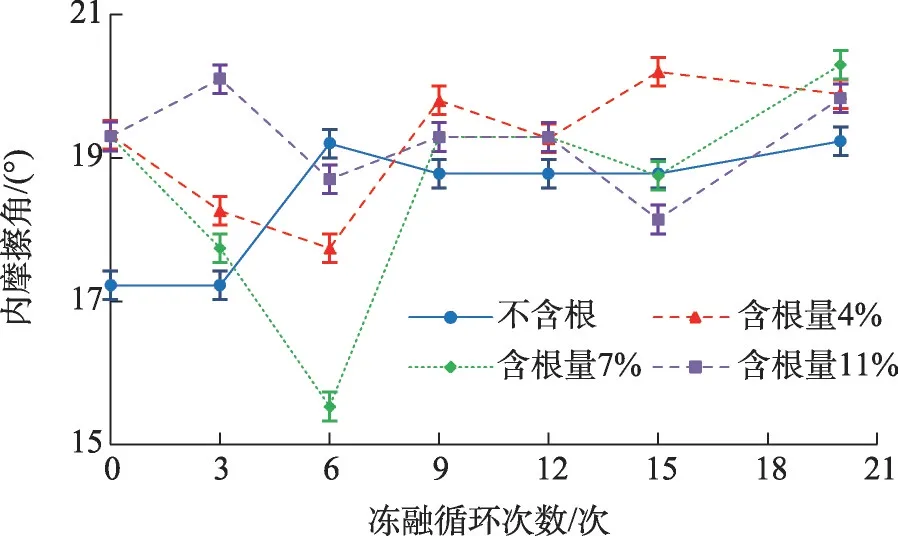
图5 内摩擦角随冻融循环次数变化关系Fig.5 Relationship between internal friction angle and freeze-thaw cycles
由图5 可知,冻融循环对试样内摩擦角的影响与黏聚力不同,总体来说呈现略微加强的作用。未经冻融循环的素土试样内摩擦角显著小于含根试样,冻融循环6次以内,不含根试样内摩擦角有明显的增加,然后随冻融循环次数的增加而变缓;而含根试样在6 次冻融循环以内,内摩擦角则表现为降低趋势,其中以含根量为7%的试样降低幅度最大,达到19.5%,表现为随冻融循环次数增加,呈波状起伏增加的变化趋势。分析可能是由于不含根试样土体孔隙相对较多,土体结构较为疏松,冻融循环作用之后,其土体大颗粒破碎,大孔隙减少使得土颗粒间的接触更为充分,从而使得内摩擦角增大,随着冻融循环次数的增大,土颗粒间结构形成新的平衡态,内摩擦角逐渐趋于平稳[8];而试样在加入根系后,整个试样土体颗粒间的接触变得更加充分,而在冻融循环作用下土体结构被破坏,土体颗粒间接触变少,因而内摩擦角减小[10];随着冻融循环次数的增大,和不含根土体类似,含根土体内部结构破坏过程停止,在9 次冻融循环以后内摩擦角呈现小范围的波动,变化较小[18]。
3 结论
(1)在相同冻融循环作用下,重塑根-土复合体黏聚力随着根系含量由4%增加至7%、11%,呈逐渐增大趋势;较素土试样,含根土体黏聚力最大增幅可达77.9%,而根系对土体内摩擦角影响幅度相对较小。根系的加入使土体形成紧密的网络结构,抵抗剪切破坏的能力增强,进而提高河岸稳定性。
(2)随着冻融循环次数的增加,含根和素土试样的黏聚力均呈现不同程度的先减小后趋于平缓的变化趋势。在冻融循环3 次时,土体黏聚力降幅达到最大,为19.0%,表明冻融初期对土体结构破坏较大,土体抗剪强度明显下降;随着冻融次数增加,其破坏能力减弱,土体结构达到新的平衡,在冻融循环达到12次时,冻融循环对黏聚力的减小幅度达到最低,仅为3.7%。
(3)冻融循环对研究区高寒草甸植被河岸抗剪强度具有一定衰减作用,当滨河高寒草甸植物退化,其地下根量减少,由于缺少根系对土体的网络、缠绕和阻隔温度变化的作用,河岸土体抗剪强度指标有不同程度的降低趋势,土体黏聚力降幅最高可达31.4%。表现为河岸稳定性降低,因此,在退化河岸带应尽量减少人类和牲畜活动,并适当开展植被恢复工作,保护河流生态。
猜你喜欢
重庆建筑(2021年3期)2021-03-31
数理化解题研究(2019年22期)2019-08-26
水利科技与经济(2017年12期)2017-04-22
西安建筑科技大学学报(自然科学版)(2016年5期)2016-11-10
铁道建筑(2016年4期)2016-10-17
科技视界(2016年7期)2016-04-01
新高考·高一物理(2015年11期)2015-12-24
遥感信息(2015年3期)2015-12-13
湖南大学学报·自然科学版(2015年5期)2015-06-16
四川建筑(2014年6期)2014-09-03
