
褪黑素诱导增强生防芽孢杆菌DZY 6715对油茶炭疽病的抑制作用*
2024-01-05 11:26刘地杨阔杜倩洁周慧琴邓佳伍健榕王芳
西部林业科学 2023年6期
刘地,杨阔,杜倩洁,周慧琴,邓佳,,伍健榕,王芳,3
(1.西南林业大学 林学院,云南 昆明 650224;2.西南山地森林资源保育与利用教育部重点实验室,云南 昆明 650224;3.西南林业大学 西南地区生物多样性保育国家林业和草原局重点实验室,云南 昆明 650224)
油茶(Camelliaoleifera)为山茶科(Theaceae)山茶属小型常绿乔木或灌木,是我国特有的木本油料植物[1]。油茶炭疽病是油茶的主要病害,重病区产量可缩减40%~80%[2]。根据研究报道,油茶炭疽病的病原菌有8种,如油茶果生炭疽菌(Colletotrichumfructicola)、胶孢炭疽菌(C.gloeosporioides)和暹罗炭疽菌(C.siamense)等,其病原菌在油茶整个生育期均可侵染[3-4]。目前,对于油茶炭疽病的主要防治措施是化学防治[5]。使用化学农药防治油茶炭疽病,虽然能高效降低油茶损失,但由于炭疽病病原菌具有较强的适应能力,容易使病原菌产生抗药性,还会对植物造成氧化胁迫使植物生长受抑[6]。因此,寻找安全无害的防治方法是研究的重要方向。目前,利用拮抗细菌控制病害的方法显示出了巨大的应用前景。国内外已有报道证明,芽孢杆菌(Bacillustequilensis)具有广谱抑菌性,抑菌机理较为复杂,通常是多种抑菌机制协同发挥作用[7-8]。芽孢杆菌防治病原菌的方式主要有:与病原菌进行营养竞争和生态位点竞争来防治病原菌;形成生物膜提高其生防效果[9];产生多种抗菌物质[10-11]来造成细胞畸变,使原生质发生凝集和外溢[11-12];分泌几丁质酶、葡聚糖酶等裂解酶来抑制病原菌菌丝生长;通过破坏细胞膜完整性和诱导孢子中活性氧的积累来抑制病原菌的生长[13-15]。此外,芽孢杆菌还可以通过促进植物生长并分泌次生代谢产物或诱导系统抗性(ISR)来防治植物病害[16]。如:徐睿等[3]发现以枯草芽孢杆菌YL 13发酵液对暹罗炭疽菌抑制效果可达到68.03%;曹娜等[17]发现芽孢杆菌发酵液具有较好的稳定性,能使病原菌菌丝发生形态变化。
芽孢杆菌DZY 6715是一株内生拮抗芽孢杆菌,对油茶炭疽病害有很好的生防效果[18],但是其防治效果与化学杀菌剂相比仍有很大差距。褪黑激素(melatonin,MT)在植物体生理调节、增强植物抗逆性方面起着非常重要的作用[19-20],可激活冷害[21]、干旱[22]和盐胁迫等多种非生物胁迫下植物的抗氧化系统,通过提高植物防御酶活性和其相对基因表达量等来提高植物的抗逆性[21]。刘亚萍[23]发现,当白菜(Brassicarapavar.glabra)灌施400 μmol/L的褪黑素溶液时,可有效降低根肿病的病情指数和发病率。目前,关于MT诱导的植物抗病内容较为丰富,但当MT作为一种诱导因子直接作用于拮抗细菌进而提高其生防效力的研究目前尚无报道。本研究以油茶为实验材料,研究特基拉芽孢杆菌DZY 6715对暹罗炭疽菌的生防效果,以探究褪黑素诱导芽孢杆菌DZY 6715抗油茶炭疽病的抗病机理。该研究结果将为探寻提高DZY 6715生防效力的途径提供理论依据。
1 材料与方法
1.1 供试材料
1.1.1 供试菌
炭疽病原菌 暹罗炭疽菌(CS)。
生防菌 特基拉芽孢杆菌DZY 6715(BacillustequilensisDZY 6715),由西南林业大学西南地区生物多样性保育国家林业和草原局重点实验室分离鉴定并保存。
1.1.2 油茶品种
2 a生的油茶幼苗(长林系列),购买于江西吉安,现种植于西南林业大学。
1.1.3 培养基
PDA培养基、LB培养基。
1.1.4 诱导物制备
褪黑素购于上海西格玛奥德里奇贸易有限公司。参考Jiang等[24]和何欢等[25]的方法配制不同浓度(0、50、100、200、400 μmol/L)褪黑素溶液并避光保存待用。
1.2 试验方法
1.2.1 不同浓度褪黑素对芽孢杆菌DZY 6715生长的影响
将预先培养并调整浓度为1×107CFU/mL的芽孢杆菌DZY 6715菌悬液(按1%接种量)加入至含有不同浓度MT(0、50、100、200、400 μmol/L)的LB液体培养基中培养(28 ℃、140 r/min),分别于12、24、48、72、96、120 h收集菌悬液并测定OD600值。以培养时间为横坐标,菌液OD600为纵坐标绘制其生长曲线。
1.2.2 不同浓度褪黑素诱导下芽孢杆菌DZY 6715对炭疽菌菌丝的抑菌活性测定
平板对峙方法参照周瑷婷[19]。处理组为不同诱导时间(24、48、72、96、120 h)和诱导物MT不同浓度(0、50、100、200、400 μmol/L),对照组为水。在7 d时测量对照组病原菌菌落半径(R1)和对峙组病原菌菌落中心到DZY 6715菌落边缘的距离(R2),并计算对病原菌菌丝的生长抑制率(R抑)。每个处理3个重复,公式如下。
1.2.3 不同浓度褪黑素诱导下芽孢杆菌DZY 6715生物膜成膜能力测定
使用结晶紫染色法,取单菌落接种到不同浓度MT(0、50、100、200、400 μmol/L)的LB液体培养基中,于28 ℃、140 rpm条件下诱导培养,并于72 h取样1 mL加入至24孔板中,后加入等体积的LB液体培养基静止培养。分别于24、48、72、96 h进行0.1%结晶紫染色,40%冰醋酸溶解,并在紫外分光光度计下测定在560 nm处的吸光值。每个处理3个重复。
1.2.4 褪黑素诱导下芽孢杆菌DZY 6715胞外水解酶活性测定
将芽孢杆菌DZY 6715接种到不同浓度MT(0、50 μmol/L)的 LB液体培养基中,于28 ℃、140 rpm条件下诱导培养,并于24、48、72、96、120 h收集菌悬液,测定几丁质酶(chitinase)和β-1,3葡聚糖酶(β-1,3-glucanase,β-1,3-GA)活性,每处理3个重复。操作步骤按照试剂盒说明书进行(试剂盒购买于苏州科铭生物技术有限公司)。几丁质酶酶活表示方法为每毫升培养液每小时分解几丁质产生1 mg N-乙酰氨基葡萄糖的酶量为一个酶活单位〔mg/(h·mL)〕,β-1,3葡聚糖酶酶活表示方法为每1 mL血清(浆)每小时产生1 mg还原糖定义为一个酶活性单位〔mg/(h·mL)〕。
1.2.5 褪黑素诱导下芽孢杆菌DZY 6715对炭疽病原菌菌丝破坏作用
将芽孢杆菌DZY 6715接种到不同浓度MT(0、50 μmol/L)的LB液体培养基中,于28 ℃、140 rpm/min条件下诱导培养,并于72 h收集菌悬液,加入CS病原菌菌丝,制成混合液,以无菌培养液为对照,每处理3个重复。28 ℃、180 rpm/min培养后,分别于24、48、72、96、120 h取样,测定电导率,10 000 rpm、4 ℃离心10 min后测定OD260,丙二醛含量(nmoL/g)和可溶性蛋白含量(mg/mL)测算按照试剂盒说明书进行(试剂盒购买于苏州科铭生物技术有限公司)。
1.2.6 不同浓度褪黑素诱导下芽孢杆菌DZY 6715对油茶炭疽病的抑制作用
选取健康且大小一致的2 a生‘长林系列’油茶叶片作为接种对象,用75%酒精将叶片消毒30 s,再用无菌水将叶片清洗3次后晾干;浸泡到MT诱导后的菌悬液(0、50、100、200、400 μmol/L)和不同浓度MT(0、50、100、200、400 μmol/L)溶液中,每个处理9个重复。放置2 h后,使用一次性注射器将叶片叶脉两侧挑破,接种直径为6 mm 的炭疽菌块,置于铺有灭菌滤纸的培养皿中,每天观察发病情况并测定病斑面积。
1.2.7 不同诱导时间下芽孢杆菌DZY 6715对油茶炭疽病抗病性的影响
叶片处理方法同1.2.6。叶片用菌悬液(50 μmol/L MT诱导72 h)浸泡处理,分别于放置2、6、12、18、24、30 h后接种直径为6 mm 的炭疽菌块,每天观察发病情况并测定病斑面积。
1.3 数据分析
利用SPSS 17.0软件对试验数据进行统计分析,应用Duncan氏新复极差法和t检验进行差异显著性检验(α=0.05)。采用Origin 2018软件进行数据处理和图形制作,Photoshop 软件进行图像处理。
2 结果与分析
2.1 不同浓度褪黑素对芽孢杆菌DZY 6715生长的影响
芽孢杆菌DZY 6715的OD600随着培养时间的延长逐渐升高,即其细胞浓度随着培养时间的延长逐渐升高并于72 h进入稳定期(图1)。进入稳定期后,50 μmol/L MT诱导下DZY 6715的细胞浓度显著高于其他浓度褪黑素诱导下的细胞浓度,且在50 μmol/L MT诱导下DZY 6715的细胞浓度在72、96、120 h分别是未诱导的1.57、1.17和1.52倍,表明50 μmol/L MT诱导处理显著提高了芽孢杆菌DZY 6715的细胞浓度。

图1 不同浓度MT处理的芽孢杆菌DZY 6715 的OD600
2.2 不同诱导时间和褪黑素浓度下芽孢杆菌DZY 6715对炭疽菌菌丝的抑菌活性
MT诱导的芽孢杆菌DZY 6715对炭疽菌的抑制效果显著高于未诱导菌株(表1),各处理抑菌率均高于50.00%。抑菌率随着诱导时间的延长呈先上升后下降趋势,其中褪黑素在50 μmol/L水平时,在不同诱导时间(24、48、72、96、120 h)下的抑菌率显著高于其他处理组,且不同浓度MT(50、100、200、400 μmol/L)在诱导72 h时抑菌率最高,分别为76.33%、72.33%、71.67%和68.67%。

表1 不同浓度MT和诱导时间下芽孢杆菌DZY 6715对炭疽菌菌丝的抑制率Tab.1 Inhibition rate of Bacillus tequilensis DZY 6715 on anthracnose mycelium under different concentrations of MT and different induction time
由图2可以看出,不同浓度MT诱导DZY 6715处理对炭疽菌菌丝并无明显影响,但在MT诱导72 h之后,可以明显看出芽孢杆菌DZY 6715在营养体上扩散面积最广,这可能是其抑制率增强的原因。

图2 不同浓度褪黑素诱导DZY 6715处理对炭疽菌菌丝生长的影响Fig.2 Effects of B.tequilensis DZY 6715 treatment induced by different melatonin concentrations on hyphal growth of anthrax
2.3 不同诱导时间诱导下芽孢杆菌DZY6715生物膜成膜能力
由图3可以看出,MT处理下的生物膜与未处理的均存在差异,所有处理培养时间与生物膜成负相关关系,培养时间越久生物膜形成能力越弱。未诱导菌株在24~48 h生物膜产量逐渐上升,72~96 h产量下降,且在静置培养96 h时,生物膜出现解离、浮游的状态。经MT诱导后的DZY 6715,其生物膜的产量明显提高且与未诱导菌株的变化趋势大致相同,但在96 h时未观察到解离状态;MT处理后在静置培养12~24 h、24~96 h分别处于最初的黏附、成膜的稳定的阶段。50 μmol/L MT处理的DZY 6715各时间段内生物膜产量均有上升,显著优于对照组,表明经50 μmol/L MT处理后的DZY 6715具有较强的生物膜形成能力,其菌悬液在静置培养24 h时OD560达到最大值,生物膜厚度明显变厚,褶皱变多,显著高于其他MT浓度处理组,这可能有助于其在植物表面定殖。


图3 不同MT处理对DZY 6715生物膜OD值的影响
2.4 50 μmol/L褪黑素诱导下芽孢杆菌DZY 6715胞外水解酶活性
由图4可知,经MT诱导过后的菌液内几丁质酶活性在不同时间段内均显著升高。不同诱导时间下50 μmol/L浓度菌液几丁质酶活性分别为2.13、2.23、2.19、2.27、2.30 U/mL。表明MT诱导处理可使菌体内几丁质酶活性升高,从而达到抑菌目的。菌液的β-1,3-葡聚糖酶活性呈现先升高后下降趋势,且MT处理组的该酶活性均较对照组显著升高。在72 h、 50 μmol/L浓度处理组的β-1,3-葡聚糖酶活性达到峰值4.5 U/mL,较对照组的酶活性3.46 U/mL显著升高了31%。表明经MT诱导处理后的发酵液β-1,3-葡聚糖酶活性显著升高,能使细胞壁降解,抑制菌丝生长。
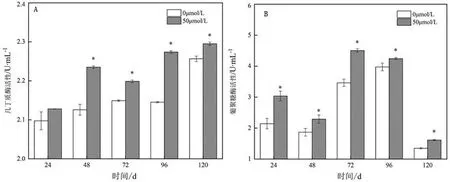
图4 褪黑素诱导下芽孢杆菌菌液胞外水解酶活性
2.5 芽孢杆菌DZY 6715对炭疽病原菌菌丝的破坏作用
如图5所示,对照组MDA浓度在120 h时间段内呈现先上升后下降趋势,处理间无显著变化;与对照组相比,相同时间50 μmol/L MT处理组菌丝体MDA浓度显著上升(P<0.05),均高于对照组和未诱导菌株,且MT处理组MDA浓度维持在较高水平,基本呈现拮抗越久,菌丝体MDA含量越高。
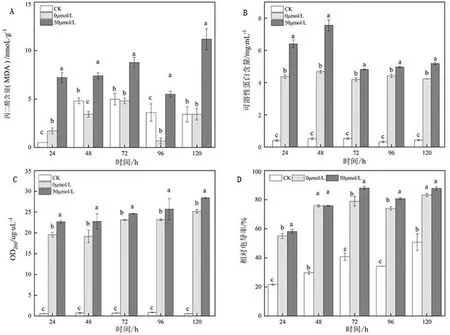
图5 MT诱导芽孢杆菌对炭疽菌的破坏作用
对照组菌体培养液中蛋白质含量极低;经MT诱导下的DZY 6715发酵液作用后,蛋白质含量迅速增加,后趋于稳定。在培养48 h时间段,50 μmol/L MT处理发酵液是对照组的13.5倍,对比未诱导菌株上升61.6%。推测MT诱导后的发酵液可能破坏了炭疽菌菌体细胞膜的完整性,导致培养液中蛋白质含量增加。
当微生物细胞膜遭到较大破坏时,核酸等大分子物质也会释放到细胞外。核酸在波长260 nm处有很强的紫外吸收,测定细胞外OD260有助于判断菌体细胞膜破坏程度。如图5所示,OD值是随着时间而升高,MT诱导处理与未诱导菌株菌液趋势一致,但50 μmol/L MT处理发酵液OD值对比未诱导菌株菌液存在显著差异,在120 h时间段中达到峰值28.43 ug/uL,说明MT诱导处理的DZY 6715增强炭疽菌核酸泄露。
在整个时段内,MT处理和CK的菌丝相对电导率总体上都呈现一个上升的趋势,且MT处理较对照组显著增加,在培养72 h时达到最大值88.09%,是对照组的1.12倍。说明与CK相比,MT处理下的炭疽菌丝的损伤程度增大,通透性增加,加速菌体细胞质渗漏到细胞外。
2.6 不同浓度MT诱导下芽孢杆菌DZY 6715在离体叶片上对炭疽菌的抑制作用
如图6、7所示,在不同浓度MT诱导下,72 h的芽孢杆菌DZY 6715在离体叶片上对炭疽病病原菌都有一定的抑制作用,且MT诱导DZY 6715处理病斑面积均小于对照组和纯培养,呈现病斑面积随MT浓度增加而增大的趋势。其中50 μmol/L MT处理发酵液有较好的抗病效果,病斑面积显著缩小,是对照组的15.3%,纯培养的28.4%。而不同浓度MT溶液对病斑面积并无明显影响,基本呈现病斑面积随MT浓度增加而减小的趋势,说明单独使用MT对油茶的病斑大小无显著影响。

图6 不同MT处理芽孢杆菌DZY6715对油茶叶片炭疽病的控制效果

图7 不同MT处理芽孢杆菌DZY 6715和单独MT对炭疽病的控制效果
2.7 发酵液处理叶片时间对炭疽病的控制效果
由图8可以看出,对比对照组来说,MT处理均对叶片病斑面积的扩增有明显的抑制效果,其中,第2 h和24 h处理组效果比较明显,较对照组缩小84.1%和88.46%。
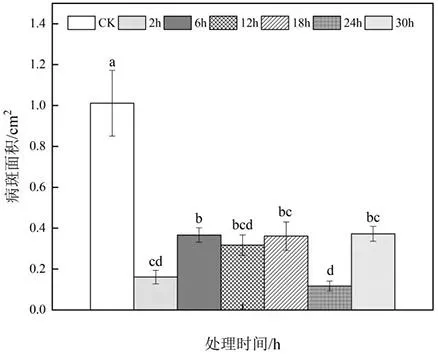
图8 发酵液处理叶片时间对炭疽病的控制效果Fig.8 Control effect of leaf time on anthracnose
3 讨论与结论
已有研究证明,天然高分子化合物可有效提高生物防治效果,化合物作为激发子诱导培养可显著提高生防菌的拮抗效力[26],促进生防菌生长繁殖和菌株活力等。本研究结果表明低浓度MT(50 μmol/L)诱导DZY 6715菌悬液细胞数量增加,且显著高于未诱导菌株,细胞浓度在72 h时达到峰值,说明MT作为生物激发子在低浓度条件下对菌株DZY 6715有一定促进生长作用,使细胞活力增强,种群密度上升。随着MT浓度的逐渐升高,其对芽孢杆菌DZY 6715的促进生长作用逐渐降低,且随着浓度的进一步升高会呈现抑制作用,与此结果类似,有研究报道当浓度高于1 000 μmol/L会显著抑制芽孢杆菌的等的生长[27]。推测MT对芽孢杆菌生长的影响可能遵循报酬递减规律,这与Yu等的结果相似[28]。而当拮抗菌株达到一定的种群密度,就可能会产生群体效应由单细胞形态转变为生物膜形态[29],帮助菌体更好地附着于植物表面,在激活植物防御反应、拮抗病原微生物等方面帮助植物抵御病害侵染。生物膜的形成可能是拮抗菌防治病害的新机理之一。MT诱导可以增强DZY 6715的生物膜形成能力,进而提升其对油茶炭疽菌的抑制作用。本研究结果表明,50 μmol/L MT诱导72 h的DZY 6715在平皿对峙试验中对炭疽菌的抑制率可达到76.33%,显著高于未诱导菌株。同时50 μmol/L MT诱导72 h 的DZY 6715在叶片上也能控制炭疽病病斑的扩展,这可能是50 μmol/L MT诱导72 h的DZY 6715细胞活力最强,能迅速占领空间,从而有效地与病原菌形成营养与空间上的竞争,抑制炭疽病菌的入侵和繁殖。此外,诱导菌株对油茶叶片的处理时间也显著影响其对炭疽病原菌的控制,处理24 h时病斑面积最小,这可能是由于50 μmol/L MT诱导72 h 的DZY 6715菌株生物膜形成能力在24 h时最强,且在叶片上处理24 h后,对病原菌产生了系统抗病性。这与何方涛[30]的研究结果类似,诱导激发培养后提高了拮抗菌对病原菌的控制效力,生防菌在抑菌过程中分泌一些物质如几丁质酶、葡聚糖酶,可破坏真菌细胞的细胞壁。林福呈等[31]研究发现,细菌可以进入真菌细胞壁繁殖并分泌溶菌物质。本研究中MT诱导可以增强DZY 6715几丁质酶、葡聚糖酶活性,破坏真菌细胞壁。细胞膜是真菌组织结构的一部分,能够维持正常的生理结构,有效调节内外环境的稳态,保障其正常生命活动[32]。已有研究表明,许多芽孢杆菌可以分泌次级代谢产物如surfactin、fengycin和difficidin等抗菌物质,这些物质可以破坏真菌细胞壁,通过改变真菌细胞膜的通透性来破坏其内外环境的稳态,从而起到抑菌作用。病原菌细胞膜受损,其内含物外渗,培养液电导率增加。MDA是膜脂过氧化的主要产物,能够衡量菌丝体的膜脂过氧化水平[32]。Yang等[33]研究发现戊二醛处理使病原菌细胞内积累大量活性氧,且MDA含量剧增,表明菌丝体发生了脂质过氧化反应,膜系统受损。而细胞膜被破坏,通透性增加,随后释放细胞内的大分子蛋白和核酸,菌体蛋白质和核酸外泄会严重影响正常的生命活动[34]。Wang[35]等研究也发现,在灰霉菌细胞膜被破坏后,会导致细胞内的大分子蛋白和核酸释放。本研究中,经50 μmol/L MT诱导DZY 6715处理的病原菌菌丝,其电导率和MDA显著高于未诱导菌株,OD260和可溶性蛋白的含量增加,说明MT诱导DZY 6715增强其对炭疽菌细胞结构的破坏,使病细胞膜的通透性增加,释放细胞内的核酸和蛋白。综上,MT诱导芽孢杆菌DZY 6715可以通过分泌相关活性酶如几丁质、葡聚糖酶,破坏病原菌的细胞壁和细胞膜,使菌体内含物外泄,进而达到溶菌目的。
实验发现,MT处理和未诱导菌株的DZY 6715菌悬液对油茶叶片的抗炭疽病效果也有不同,其中50 μmol/L MT平板对峙抑菌率为76.33%,表明诱导后的菌株与病原菌对峙培养后,能迅速占领空间,从而有效地与病原菌形成营养与空间上的竞争[35-36]。叶片离体实验表明,MT诱导后的芽孢杆菌DZY 6715处理油茶叶片,其病斑面积缩减,50 μmol/L MT处理表现最佳,这与何方涛[30]的研究结果类似,诱导培养后提高了拮抗菌对病原菌的控制效力。由50 μmol/L MT诱导的DZY 6715处理后,再放置不同时间后接种病原菌于叶片,均可诱导叶片产生抗病性,从而增强植株自身防御反应,且以放置24 h后病斑面积最小。这可能是因为DZY 6715菌株的生物膜形成能力在24 h时最强,可以在油茶叶片上进行定殖,从而提高拮抗能力,而单独施用MT对油茶叶片抗病并无明显作用。
综上所述,本研究结果表明低浓度的MT诱导可以促进芽孢杆菌DZY 6715生长,且50 μmol/L MT诱导后的菌株生防效果最好;MT作为诱导激发子低浓度时可以增强芽孢杆菌DZY 6715对暹罗炭疽菌的抑制效果,且50 μmol/L MT诱导72 h的DZY 6715菌株效果最好;诱导的DZY 6715菌株具有较强的生物膜形成能力,分泌几丁质和葡聚糖酶,破坏了病原菌菌丝的结构,改变细胞膜的通透性,使细胞内物质泄漏,从而使病原菌溶解死亡。本研究结果可为油茶高效生防菌剂开发利用提供理论参考,使其大规模应用并替代传统化学杀菌剂成为可能。然而,虽然明确了MT作为诱导激发子可以增强芽孢杆菌DZY 6715对油茶炭疽病病菌的抑制作用,但其抑菌机制和抗病机理尚不明确。后续宜对植物微观结构、生理生化及转录水平进一步深入研究。
猜你喜欢
今日农业(2022年15期)2022-09-20
现代临床医学(2021年5期)2021-11-02
昆明医科大学学报(2021年4期)2021-07-23
现代畜牧科技(2021年4期)2021-07-21
现代园艺(2017年21期)2018-01-03
现代园艺(2017年21期)2018-01-03
中国西部(2017年4期)2017-04-26
湖南农业(2017年1期)2017-03-20
营销界(2015年23期)2015-02-28
中华皮肤科杂志(2014年4期)2014-12-19
