
雷公山森林群落结构组成及其随海拔梯度的变化
2024-01-05 08:40张菊花张旭杨光能刘延惠余永富余德会
贵州林业科技 2023年4期
张菊花,张旭,杨光能,刘延惠*,余永富,余德会
(1.贵州省林业科学研究院,贵州 贵阳 550005;2.贵州雷公山国家级自然保护区管理局,贵州 雷山 557100;3.贵州雷公山森林生态系统国家定位观测研究站,贵州 雷山 557100)
自然森林群落是在长期的历史演变过程中发展而成的植物复合体[1-2]。不同的森林群落在结构上存在很大差异,这种差异主要源于组成物种的不同生态、生物学特性及它们的构成方式[3]。从尺度来看,在大尺度上,森林群落结构受气候的影响;在山地区域尺度上,海拔是森林群落结构的重要决定因素[4-5],主要是由海拔变化引起的水分、温度和光照强度等生态因子改变的综合作用的结果[6]。随着海拔的上升,水热、土壤等环境发生变化[7],进而会影响森林群落结构组成。
植物的种类组成和结构是一个群落最基本的要素,是了解其组成、变化和发展趋势的基础[8]。此外植物群落结构和组成的相似性或差异性不仅反映了群落当地生境的差异性或多样性[9],而且在一定程度上反映了群落的复杂性[10]。因此,分析植物森林群落的结构组成,对揭示群落所在生境的状况以及群落的稳定性与演替规律具有极为重要的意义。
不同森林群落间的交错区表现的程度不同。有些群落间具有明显的过渡特征变化[11-13];有些群落间的界限并不是很明显,而是由不同群落间物种逐渐更替变化[14-15]。有研究表明植物类型受海拔梯度的影响存在一定的变化趋势,这种变化趋势在某种意义上可以反映出物种对气候变化的响应[16-17]。学者将用DCCA排序法研究植物群落与6个环境因子(海拔、坡度、土层厚度、枯枝落叶层厚度、坡向和人类干扰)之间的关系,得出海拔高度和人类干扰强度是制约植物群落结构及分布格局的决定因子[18]。但不同地区的森林群落受环境因子的影响有差异。雷公山自然保护区处于亚热带季风山地湿润气候,有着地理位置独特、森林类型多样,森林植被垂直带谱明显、生物多样性丰富等特点,属生物地理区较好代表之一。其森林群落的组成和结构随海拔梯度的不同而有着相对明显的差异,以往的调查工作虽对此做过一些考察,但因调查工作有限,无法系统地说明群落组成结构沿梯度变化的详细情况[19]。因此,本研究通过对雷公山自然保护区内不同海拔的森林群落进行相关研究,探寻森林群落组成结构及其对海拔梯度变化的响应,对掌握不同海拔下植物响应规律具有重要的参考意义,可以进一步为该地区资源保护研究提供一定的科学依据。
1 研究地区与研究方法
1.1 研究区概况
研究区位于贵州省黔东南苗族侗族自治州雷公山国家级自然保护区(26°15′~26°22′ N,108°09′~108°22′ E),保护区总面积4.73万hm2,是清水江和都柳江水系主要支流的发源地,是长江和珠江流域的分水岭。雷公山山体高大,气候类型多,相对湿度大,阴雨天气多。保护区最高海拔达2178.8 m,最低海拔650 m,相对高差在1500 m以上,年平均温度随海拔升高而降低,年平均温度在14~16 ℃,山腰11.7 ℃,山顶9.2 ℃;年降雨量在1300~1600 mm之间;降水多集中在春季和夏季,全年太阳总辐射值仅为3642.5~3726.3 MJ·m-2。
1.2 样地选择和设计
对雷公山自然保护区内不同植被群落、海拔、坡向、坡位等指标采取典型抽样法,沿雷公山海拔较低处(约795 m)至雷公山顶,以每200 m海拔高度为间隔,在阳坡或半阳坡面,选择典型地块,分别设置一块固定样地。样地大小为30 m×30 m,沿对角线设置5 m × 5 m 的灌木样方3个,1 m×1 m的草本样方3个。对标准样地内乔木树种进行每木检尺,记录各树种名称、胸径、树高、冠幅等指标,对灌木和草本记录名称、株数、盖度、平均高等,同时记录样地海拔高度、坡度、坡向、坡位等情况(表1)。
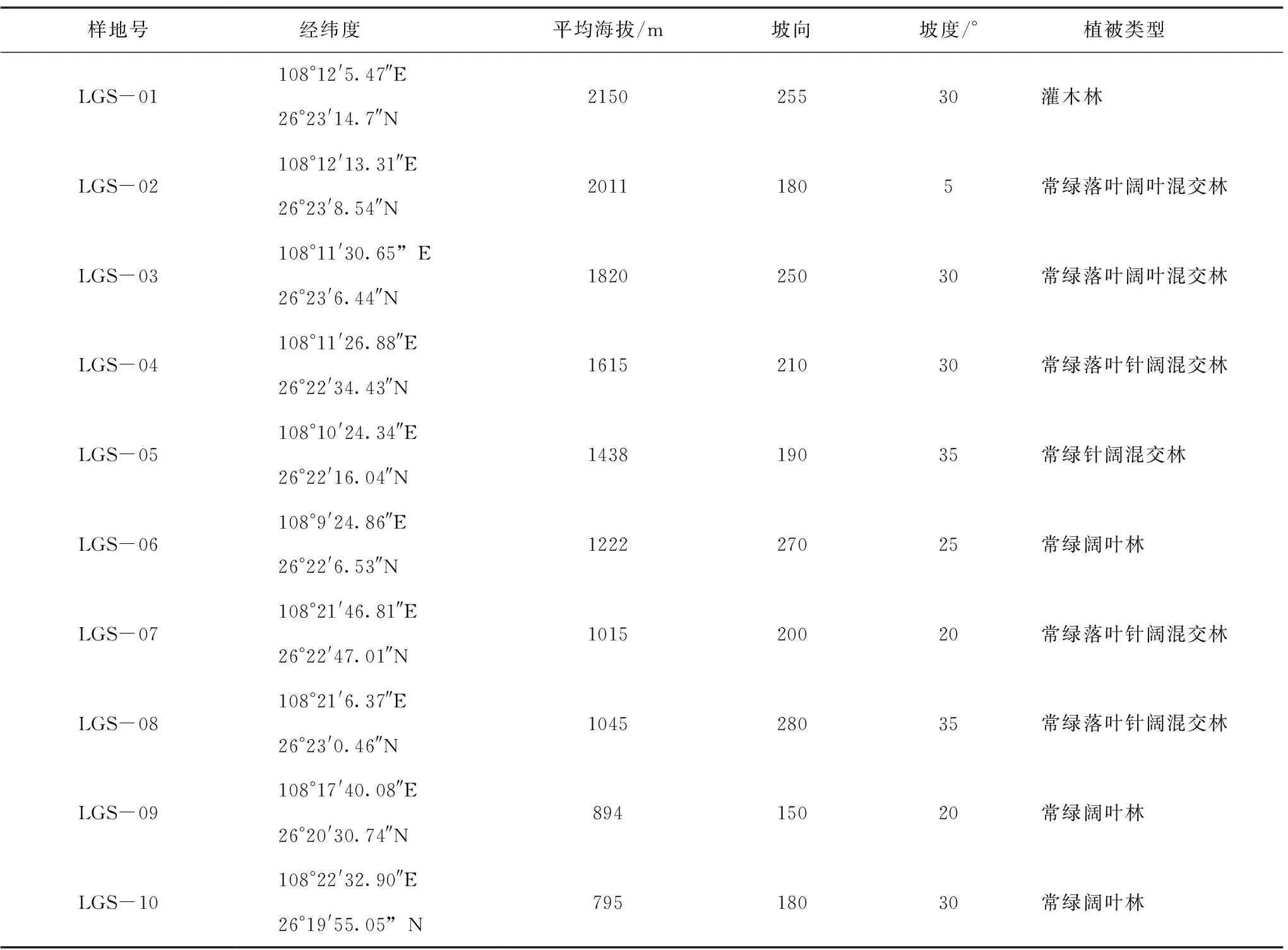
表1 样地信息
1.3 数据处理
对雷公山10个样方中出现的乔木物种,剔除掉频度<5%的偶见种后得46个种,形成10×46的原始数据矩阵。利用公式计算46个种的重要值,形成10×46的样方-重要值数据矩阵。环境数据包括4个环境因子(海拔、坡向、坡度、人为干扰强度),组成10×4的样方-环境因子矩阵用于植被数量分类和排序。海拔和坡度用实测的数据表示,坡向和人为干扰的数据量化转换,基于相关文献[20-21]中阴坡:0~45°;半阴坡:45°~135°;阳坡:135°~225°;半阳坡:225°~315°;阴坡:315°~360°。将坡向具体量化为:1表示半阴坡;2表示半阳坡;3表示正阳坡。人为干扰强度则分为3个等级:1表示较轻微的人类干扰;2表示中等程度的人类干扰;3表示比较强烈的人类干扰。
乔木重要值计算:IV=(相对密度+相对频度+相对胸高断面积)/3。
采用Excel 2010和Canoco 5.0[22]进行数据处理和分析。
2 结果与分析
2.1 森林群落的树种组成
植物群落种类组成是群落结构的基础,也是植物群落最主要的特征之一[23]。通过数量统计分析群落种类组成,进而判断群落类型。雷公山地带性植被,属我国中亚热带东部偏湿性常绿阔叶林,树种组成丰富多样,主要树种为阴香(Cinnamomumburmannii)、丝栗栲(Castanopsisfargesii)、青冈栎(Cyclobalanopsisglauca)、白栎(Quercusfabri)、贵定山柳(Salixpseudotangii)、杉木(Cunninghamialanceolata)、白檀(Symplocospaniculata)、云锦杜鹃(Rhododendronfortunei)、钝叶木姜子(Litseaveitchiana)、波叶红果树(Stranvaesiadavidiana)、青冈(Quercusglauca)、野茉莉(Styraxjaponicus)、清香木姜子(Litseamollis)、黄丹木姜子(Litseaelongata)、水青冈(Faguslucida)、云贵鹅耳枥(Carpinuspubescens)、多花山矾(Symplocosramosissima)、花楸(Sorbuspohuashanensis)、光皮桦(Betulaluminifera)、枫香(Liquidambarformosana)、青榨槭(Acerdavidii)、龙肾柿(Diospyroskaki)、檵木(Loropetalumchinense)、狭叶润楠(Machilusrehderi)。
由表1、2可知,海拔在2150 m的林地云锦杜鹃占比高达56%,表明云锦杜鹃是先锋树种;海拔在2011 m的林地内主要树种有白檀、云锦杜鹃、野茉莉等18种灌木和小乔木,其中白檀和云锦杜鹃分别占34%和25%;海拔在1820 m的林地内主要树种有白檀、青冈、贵定山柳等46种乔木,其中白檀和青冈占比达37%,其余占比都小于10%。随着海拔的降低,树种组成趋于多样,形成林分的树种杂乱,树种占比都小于10%,其主要树种所占比例都接近,说明各树种在该样地比较均匀,种间竞争激烈。对海拔梯度跟树种组成(树种种类)进行相关性分析,结果表明(表3):随着海拔降低形成天然林树种的种数逐渐增多,海拔高度与树种组成之间呈现显著负相关关系(p<0.01)。
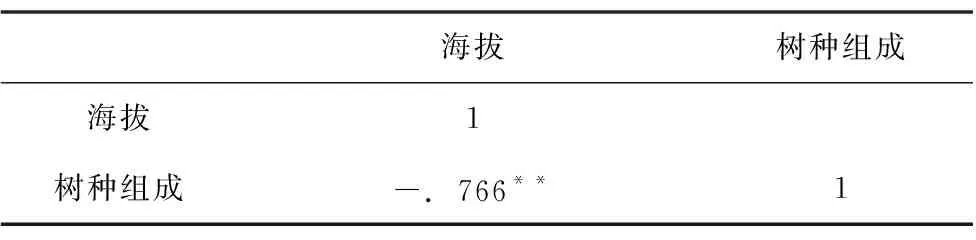
表3 各海拔高度与其树种组成的相关性
总体上,按海拔梯度分析各主要建群种,其垂直分布格局较清晰。各主要建群种由下而上分布次序为:阴香、丝栗栲、青冈栎、白栎、贵定山柳、杉木、白檀、云锦杜鹃。
2.2 森林群落结构的过渡趋势
经调查发现,雷公山植被类型垂直分布明显,在不同的海拔高度,森林群落并没有存在明显的转换过程,而是呈现为慢慢过渡的趋势。表4中低海拔以青冈栎、丝栗栲和黄丹木姜子为主,随着海拔的逐渐上升,在海拔为1015 m时,开始出现针叶树种杉木,海拔达到1438 m时,杉木转变为建群种,形成针阔叶林混交林;海拔为1615 m时,针叶树种减少直至消失;在海拔为1615~1820 m阶段,随着地势升高,气候、土壤均发生了变化,植被中常绿树种逐渐减少,转变为以青冈栎、长梗木莲等为主的常绿落叶阔叶混交林林型,因此,在2000 m左右的海拔跨度范围内,群落出现了针叶林向常绿阔叶林的转变过程;海拔为2011 m时,高大乔木减少并出现云锦杜鹃等小灌木,直至山顶云锦杜鹃转变为建群种,由大量的云锦杜鹃和箭竹形成矮小的灌木林。雷公山相对湿度随着海拔的增高而加大,海拔1100 m处年平均相对湿度为85%,海拔1600 m的林地相对湿度为88%,海拔2178.8 m的山顶相对湿度高达91%。因此,在海拔为2150 m的样地1处,主要是因为湿度大和山顶地形裸露因素,树干矮化,苔藓植物发育强,出现山顶苔藓多,从而形成矮树林。

表4 各海拔样地群落类型
雷公山属我国中亚热带东部偏湿性常绿阔叶林,但在调查的所有样地中,样地7和样地8存在较明显人为活动,人为活动频繁会对林分的结构有所影响,在该海拔的林地上出现了针叶树种和落叶阔叶种,植被类型的变化受人为干扰,森林群落结构发生变化。
2.3 环境因子与森林群落的相关性
为了更好地阐明森林群落结构与环境因子(海拔/坡度等)之间的关系,采用CCA排序分析,能直观地反映环境因子对森林物种组成的解释程度。从表5可知,第一轴和第二轴的解释量分别为90.87%、81.99%,说明所选的环境因子(海拔、坡向)可以解释海拔、坡向与森林物种组成之间差异的90.87%和81.99%。

表5 森林群落环境因子与CCA排序轴相关性
在CCA排序图中,各环境因子的线条长度表示环境因子与植物群落关系的大小,两条线条之间的夹角表示这两个环境因子之间相关性的大小。从图1可以看出海拔高度(ALT)、坡向(SA)、坡度(SD)是决定雷公山树种分布的主要环境因子。其中,海拔高度是决定森林群落形成的主导因子。由于海拔高度变化大,且不同海拔间的温度和湿度差异较大,对树种分布有决定性的影响。坡向和坡度的箭头长度仅次于海拔,说明坡向和坡度对植物的分布也具有较大影响,尤其是对树种组成的影响更为显著。相对于海拔、坡向和坡度而言,人为干扰(HD)的箭头最短,说明对树种分布的影响最小,由于本研究的对象都是自然森林群落,人为干扰很少,只有样地7和样地8有人为干扰,所以对植物群落稍微有些影响。
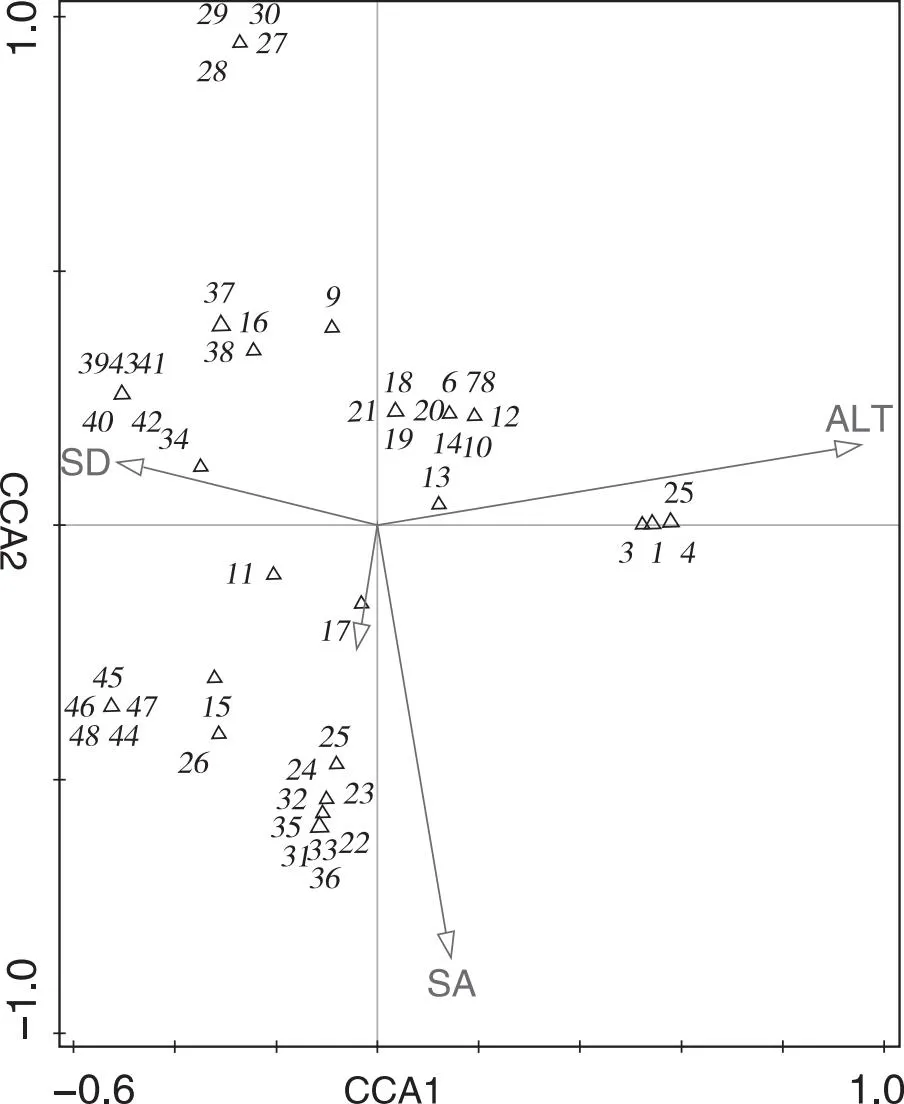
注:ALT:海拔;SA:坡度;SD:坡向;HD:人为干扰强度;1~48为所有样方中频度大于5%树种的重要值。
3 讨论
雷公山森林群落的建群种沿海拔梯度均呈规律性的变更,主要建群种由下而上分布次序是阴香、丝栗栲、青冈栎、白栎、贵定山柳、杉木、白檀、云锦杜鹃。先锋树种先由阔叶树种转变为针叶树种,后由针叶树种转变为阔叶树种,且树种的种类随海拔升高而逐渐减少。这一结论与山地垂直分布带物种丰富度沿海拔梯度呈现下降趋势[24]相同,但与长白山河岸带群落物种丰富呈现出高海拔低、中低海拔高[25]和中部较高的分布规律[26-27]不同。说明雷公山符合山地植物群落垂直梯度的变化规律。
雷公山自然保护区最高海拔为2179 m,最低海拔为650 m,相对高差在1500 m以上,各海拔梯度分布着不同的植被类型,由下自上依次分布着常绿阔叶林、常绿针阔叶混交林、常绿落叶阔叶混交林和山顶矮林。植被类型转换的过程中,阔叶树种的优势地位逐步被杉木针叶树种所取代,而后又恢复。低海拔地区为纯阔叶林,中海拔地区出现针叶树种,从而形成针叶树种为优势种的常绿针阔叶混交林,较高海拔地区优势种为阔叶树种的硬叶常绿阔叶林,高海拔山顶则形成了矮林。这进一步验证群落沿环境梯度呈连续性分布的观点[28]。此外,有研究表明人为干扰不仅改变了其沿海拔梯度的分布格局,而且加大了不同海拔区域群落之间的波动幅度[25]。本研究中样地7和样地8为常绿落叶针阔混交林,突然出现落叶阔叶和针叶树种,推断可能是由于人为因素的干扰。
相关研究表明,在山区,海拔作为山地植物群落分布格局的主要制约因子,通过太阳辐射和降水的空间再分配,使植被类型发生变化,进而决定植被的分布格局,从而影响群落类型的结构组成,最终决定山地植被的群落类型[29-30]。本研究中海拔是影响森林群落的决定性因素,与上述的研究结果相似。其次是坡向和坡度对群落分布影响,在该研究所有10个样地中,阳坡和半阳坡占80%,半阴坡占20%,一般来说,阳坡的水热条件比阴坡的较优渥,因此分布在阳坡的树种也是相对较多,从而改变森林群落的结构。坡度主要影响着环境湿度,坡度越陡,土壤水分容易流失,湿度就越小,土壤也较为贫瘠。坡度的变化能够创造出一些多样的小生境条件,由于物种生境条件的限制,使得它们会以斑块状的形式存在于特定的小生境内。从而坡向和坡度间接影响了物种组成和群落格局。
就环境因子而言,本研究对群落影响较大的环境因子有海拔、坡向、坡度,这与山地地区的研究结果一致[22]。森林群落结构不仅受自然环境因子的影响,还有人为干扰强度的制约,例如河岸带的森林群落受人为干扰的影响较大,改变了其沿海拔梯度的分布格局[31]。本文研究对象为雷公山国家自然保护区内的自然森林群落,因此人为干扰强度对森林群落的影响较低。为了更好地解释该地区森林群落结构的变化机制,后续可以对土壤因子、气象因子等其他环境因子进行相关研究。
猜你喜欢
现代园艺(2021年23期)2021-12-01
林业勘查设计(2020年1期)2021-01-18
新农业(2020年18期)2021-01-07
测绘学报(2019年11期)2019-11-20
农民致富之友(2018年23期)2018-12-19
吉林农业(2016年12期)2017-01-06
土壤与作物(2015年3期)2015-12-08
原生态民族文化学刊(2014年1期)2014-08-08
中央民族大学学报(自然科学版)(2014年2期)2014-06-09
河池学院学报(2014年5期)2014-02-27
