
退化喀斯特森林不同自然恢复阶段土壤养分和酶活性特征*
2024-01-10 01:52蒙胧晨曦陈路瑶吴宇航陈应群邵雪蓉张广奇陈丹梅刘庆福隋明浈丁访军臧丽鹏
内蒙古大学学报(自然科学版) 2023年6期
李 雍,蒙胧晨曦,陈路瑶,吴宇航,陈应群,邵雪蓉,张广奇,,陈丹梅,,刘庆福,,隋明浈,,丁访军,臧丽鹏,
(1.贵州大学林学院森林生态研究中心,贵阳 550025;2.贵州荔波喀斯特森林生态系统国家定位观测研究站,贵州 黔南布依族苗族自治州 558400)
土壤作为植物生长发育的场所,为植物的定殖、发育提供必需的营养物质,而土壤养分作为土壤最基本的特征[1],其分布格局直接制约植物的生长分布[2]。但由于气候、地形及植被类型的差异造成不同区域限制植物生长的因子差异较大,例如,研究表明,中亚热带地区植被生长受土壤磷含量的限制,而荒漠草原植物生长受土壤氮含量的限制[3]。目前,普遍观点是我国北方植被分布的主要限制性土壤养分是氮含量[4],而在南方地区磷元素成为植被分布的限制性因素[5]。正确识别限制性土壤养分是生态学研究的基础。
无处不在的干扰会造成生态系统的逆向演替[6],常表现为生态系统功能衰退、土壤肥力下降、生物多样性减少[7],当干扰终止后,植被会沿既定方向恢复至顶级群落[8]。而植被恢复过程中,生物群落与非生物环境相互影响,一方面,群落的更替会改变土壤的理化性质;另一方面,土壤养分含量的变化也会影响植物定殖、发育,进而改变群落结构[9],现有研究表明,在植被恢复进程中,土壤养分含量具有下降趋势,进而引起物种组成差异及生态策略迁移[10-11]。因此,土壤养分含量变化也被作为评价植被恢复的重要指标。此外,土壤作为陆地生态系统的重要组成部分,其质量可以直接或间接地影响生态系统功能表达[12-13],而土壤酶活性是其功能表达的重要度量指标。研究表明,土壤养分含量的变化可以引起土壤酶活性的变化[14-16],恢复时间、林分结构等指标均对土壤酶活性具有显著影响[17-18]。然而,就目前研究来看,土壤养分与酶活性在生态系统恢复过程中的协同与权衡变化规律认识较少,尤其是在非地带性植被恢复过程中的变化规律未见报道。
我国西南地区是生态脆弱区之一[19],其喀斯特面积约54万km2[20]。喀斯特地貌上极易发生石漠化,严重制约了当地经济发展与生态修复,而石漠化最为直观的表征是植被的退化和大面积的基岩裸露[21],这种裸露的碳酸盐岩溶解富含钙元素,而钙极易与有机质和磷形成稳定的化合物[22],使得该地区土壤氮磷元素的供给能力下降。研究显示,喀斯特地区植被恢复早期,土壤氮元素供给水平较低,在恢复后期,土壤中缺乏磷元素较为明显[23]。伍方骥等[24]研究发现,通过植被恢复的方式既可以提升喀斯特地区土壤有机碳的含量,也能显著提升土壤全氮含量。但以往研究多关注土壤养分的变化,却忽略了土壤酶参与森林碳氮磷代谢的重要作用[25],针对不同自然恢复阶段的土壤酶活性的研究较少,且尚不清楚随着植被自然恢复进程的进行主导土壤养分变化的因子,也不能明确土壤养分与酶活性的相关性随着恢复进程的进行如何变化。因此,探讨喀斯特不同自然恢复阶段土壤养分与酶活性变化特征对该地区生态系统保护工作具有重要指导意义。本文以茂兰喀斯特常绿落叶阔叶混交林自然恢复序列为研究对象,通过测定不同自然恢复阶段的土壤养分及酶活性,利用方差分析及多重比较、主成分分析和冗余分析等方法,系统地探究土壤养分及土壤酶活性的分布特性、主导因素及不同恢复阶段内土壤养分与酶活性的关联特性,以期为喀斯特森林生态修复提供理论基础。
1 研究地概况与研究方法
1.1 研究地概况
研究地位于贵州省东南部的茂兰国家级自然保护区(25°09′—25°20′N,107°52′—108°05′E),保护区海拔最高处达1078 m,最低处430 m,总面积21285 hm2,其中核心区5827 hm2,缓冲区8910 hm2,实验区4588 hm2。该地区为中亚热带季风温润气候,春秋温暖、冬无严寒、夏无酷暑、雨量充沛。研究地年平均气温为15.3℃,气温年均差18.3℃,最冷月平均气温为5.2℃,最热月平均气温为23.5℃,≥10℃活动积温为4598.6℃,生长期237天,全年降水量1752.5 mm,集中分布在4—10月。研究区域成土母岩为碳酸盐岩,土壤类型以黑色石灰土为主。当地代表植被类型为亚热带常绿落叶阔叶混交林,主要树种有小花梾木(Cornus parviflora)、云贵鹅耳枥(Carpinus pubescens)、多脉榆(Ulmus castaneifolia)、香叶树(Lindem communis)、青冈(Quercus glauca)等。根据保护区参考资料查证,研究地范围内主要干扰方式为樵采,而依据恢复年限可大致分为3个阶段:1)灌木林阶段:樵采后自然恢复20~30年;2)次生林阶段:樵采后自然恢复50~60年;3)老龄林阶段:有记录以来未经过明显人为干扰。
1.2 样地建立及土壤样品采集
于2022年7 月在灌木林(Shrubbery,TG)、次生林(Secondary-growth forest,SG)、老龄林(Oldgrowth forest,OG)3种自然恢复阶段森林内,参照《全国生态状况调查评估技术规范—森林生态系统野外观测》(HJ1167-2021)方法,各建立10块30 m×30 m 固定样地,样地与样地间距离>50 m,每阶段主要优势物种见表1(按重要值排序)。在已建立的固定样地内,使用五点取样法在每个样方中心位置取表层土(0~20 cm 深度)各500 g,将5包土壤样品去除杂质后充分混匀,装袋后带回实验室,在0℃下冰箱中保存。待解冻土壤风干后,对土样进行研磨,一部分过0.25 mm 和2.00 mm 筛以供土壤理化性质测定,一部分过1 mm 筛用于酶活性指标的测定。样地情况与采样方法等见图1。
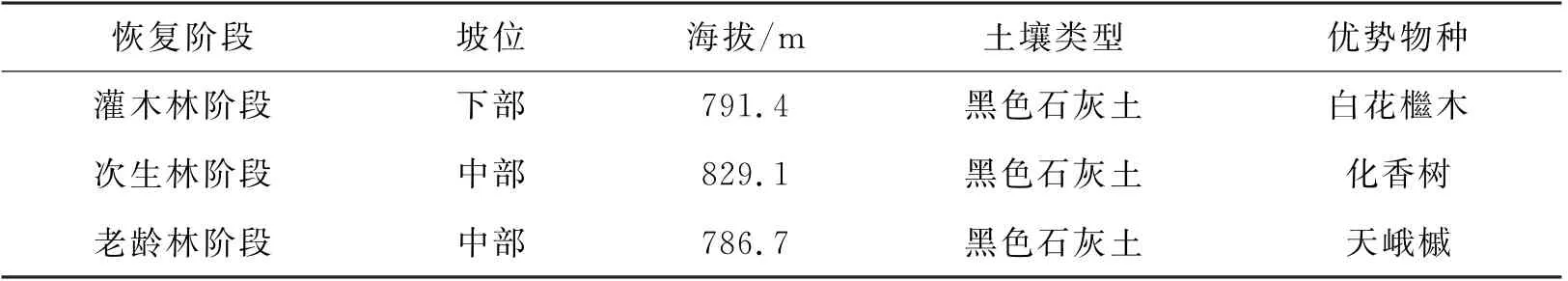
表1 样地基本特征Table 1 The basic characteristic of the plots
1.3 土壤养分含量及酶活性测定
土壤p H 值使用电位法测定,土壤碱解氮(AN)采用碱解—扩散法测定,速效钾(AK)含量用乙酸铵浸提—火焰光度法测定,全钾(TK)使用NaOH 熔融—火焰光度法测定,全磷(TP)测定使用氢氧化钠碱溶—钼锑抗比色法,有机磷(OP)使用灼烧酸浸提—钼锑抗比色法,上述养分测定方法见鲍士旦的《土壤农化分析》一书[26];铵态氮()、硝态氮()的测定参照“环保标准(HJ 634-2012)氯化钾溶液提取—分光光度法”;脲酶(S-UE)、纤维素酶(S-SC)、蔗糖酶(S-CL)、中性磷酸酶(S-NP)、中性蛋白酶(S-NPT)、脱氢酶(S-DHA)等酶活性使用索莱宝试剂盒测定[27]。
1.4 数据处理
使用R4.2.3分析软件进行数据处理和图表处理,采用“vegan”包进行单因素方差分析及多重比较(P<0.05)对比分析喀斯特森林不同恢复阶段内土壤养分含量及酶活性变化趋势;使用“Facto-MineR”和“factoextra”等包进行主成分分析,分析土壤养分含量和酶活性对不同恢复阶段的主导性因子;使用“vegan”和“ggrepel”等包进行冗余分析,分析不同恢复阶段土壤养分与酶活性间的相关性及其排序;所有图均使用“ggplot2”包进行处理。
2 结果与分析
2.1 退化喀斯特森林自然恢复进程中土壤养分特征分析
对不同自然恢复阶段的土壤养分含量进行单因素方差分析,结果如图2所示。随着自然恢复的进行,土壤全磷含量在3个阶段存在显著差异,土壤全钾、有机磷在前两个阶段差异显著,土壤全钾含量随着恢复进行先下降再上升,土壤有机磷的含量则与之相反,而土壤全磷含量随着恢复进行呈现上升的趋势,且差异显著;碱解氮、p H 值、铵态氮、硝态氮等养分含量在灌木林阶段与后面两个阶段存在显著差异,且土壤碱解氮、p H 值、硝态氮含量随恢复进行逐渐上升,而铵态氮含量随恢复阶段呈现下降趋势;速效钾在3个自然恢复阶段均无显著性差异。

图2 退化喀斯特森林自然恢复阶段土壤养分特征方差分析结果Fig.2 Analysis of variance of soil nutrient characteristics during the natural recovery stage of degraded Karst forests
2.2 退化喀斯特森林自然恢复进程中土壤酶活性特征分析
对不同阶段土壤酶活性特征进行单因素方差分析,结果见图3,随着自然恢复进程的发生,土壤蔗糖酶、脱氢酶、纤维素酶并没有显著性的差异;土壤中性蛋白酶、脲酶、中性磷酸酶存在显著性差异,且脲酶、中性磷酸酶随着恢复阶段的进行活性逐渐上升,而中性蛋白酶活性到达次生林阶段后趋于稳定。
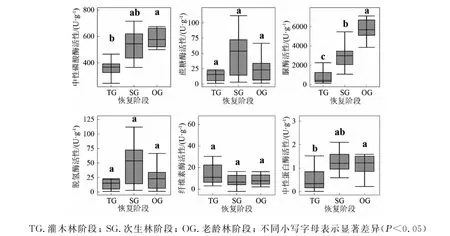
图3 退化喀斯特森林自然恢复阶段土壤酶活性特征方差分析结果Fig.3 Variance analysis of soil enzyme activity characteristics during the natural recovery stage of degraded Karst forests
2.3 退化喀斯特森林不同自然恢复阶段土壤养分及酶活性主导因子分析
2.3.1 退化喀斯特森林不同自然恢复阶段土壤养分及酶活性主成分分析
对不同恢复阶段土壤养分特征进行主成分(PCA)分析,结果如表2所示。3个阶段的主成分分析均显示前3个主成分特征根大于1,且累计贡献率分别达到78.72%、80.81%、69.88%,所以前3个主成分轴能够表征土壤养分的总体情况。灌木林阶段(TG)第一主成分轴土壤养分主要决定性因子为土壤硝态氮、全磷,第二主成分轴土壤养分主要决定因子为p H 值和有机磷,第三主成分轴土壤养分主要决定因子为土壤速效钾和碱解氮;而次生林阶段(SG)第一主成分轴中土壤养分的主要决定因子为速效钾和全磷,第二主成分轴的主要决定因子为铵态氮和p H 值,第三主成分轴主要决定性因子为硝态氮;老龄林阶段(OG)第一主成分轴土壤决定性因子为速效钾和碱解氮,第二主成分轴的主要决定性因子为碱解氮和全磷,第三主成分轴主要决定性因子为有机磷。

表2 退化喀斯特森林自然恢复阶段土壤养分主成分分析Table 2 Principal component analysis of soil nutrient dynamics during the natural recovery stage of degraded Karst forests
对不同恢复阶段土壤酶活性特征进行主成分分析,结果如表3所示。灌木林阶段(TG)前两个主成分特征根大于1,累积贡献率达到74.57%,能够有效反映土壤酶活性的基本情况,其中,第一主成分轴的主要决定性因子为脱氢酶和中性磷酸酶,第二主成分轴的主要决定因子为蔗糖酶和中性蛋白酶。次生林阶段(SG)3个主成分轴的特征根均大于1,累积贡献率为86.67%,所以选取前3个主成分轴表示该阶段土壤酶活性总体情况,其中第一主成分轴土壤酶活性的主要决定因子为脲酶和蔗糖酶,第二主成分轴的主要决定性因子为脱氢酶和中性蛋白酶,第3主成分轴的主要决定因子为中性磷酸酶。老龄林阶段(OG),仅有前两个主成分轴的特征根大于1,其累积贡献率达到70.04%,故前两个主成分轴能较好地表征该阶段土壤酶活性总体情况,该阶段第一主成分轴土壤酶活性的主导因子为蔗糖酶和脱氢酶,第二主成分的主导因子为脲酶和中性磷酸酶。
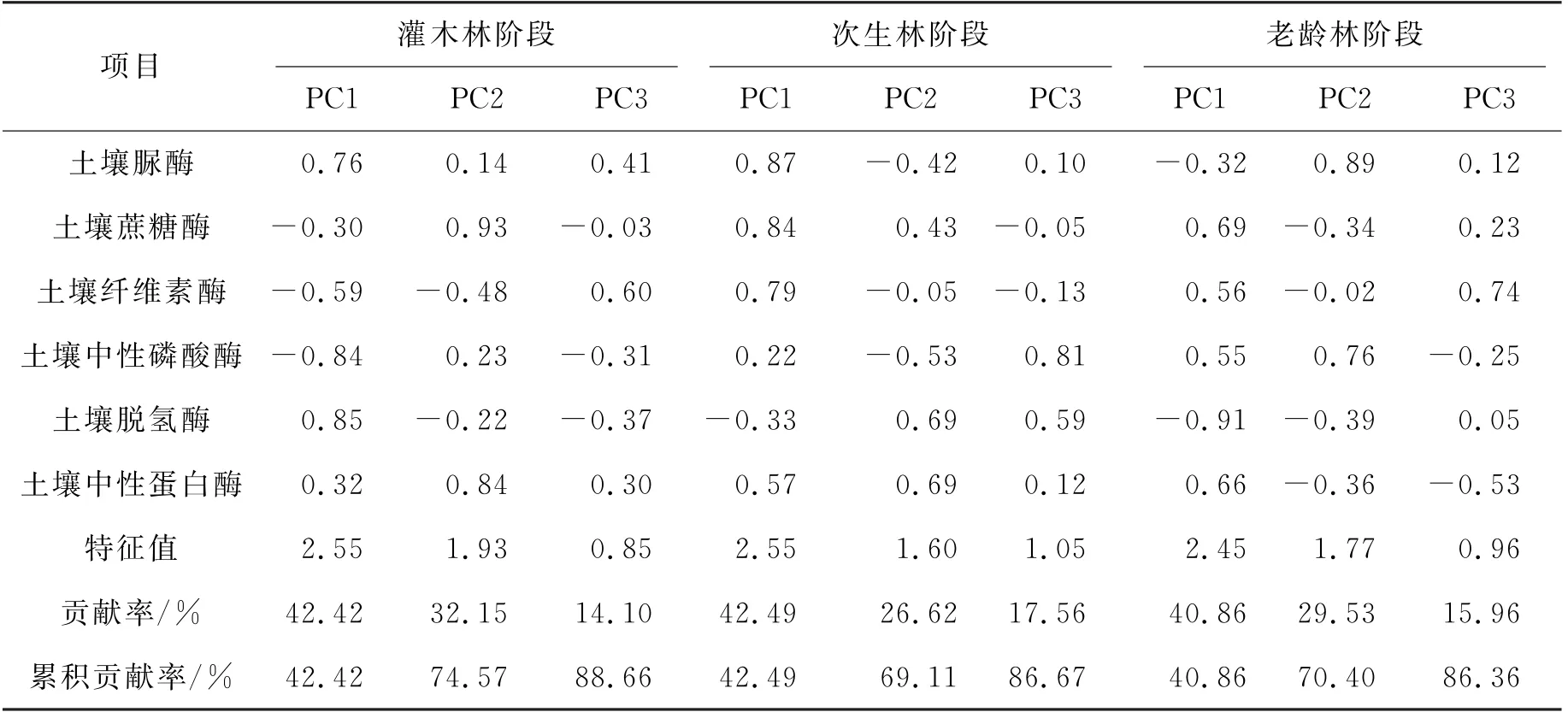
表3 退化喀斯特森林自然恢复阶段土壤酶活性主成分分析Table 3 Principal component analysis of soil enzyme activity during the natural recovery stage of degraded Karst forests
2.3.2 退化喀斯特森林不同自然恢复阶段土壤养分及酶活性RDA 分析
根据RDA 分析(冗余分析)原理,在进行分析之前对土壤养分与酶活性数据进行去趋势对应分析(DCA),结果显示3个阶段土壤养分—酶活性中DCA 梯度最长的排序轴长度均小于3,因此选择线性模型进行RDA 排序,结果见图4。其中,灌木林阶段RDA 第一、第二轴分别解释了土壤养分—酶活性数据中的41.40%和29.47%的方差(图4A),采用Monte Carlo Permutation Test对影响土壤酶活性的8个土壤养分因子进行排序,其与酶活性的相关性依次为全磷>硝态氮>p H>速效钾>碱解氮>全钾>有机磷>铵态氮;次生林阶段RDA 第一轴解释了土壤养分—酶活性数据的41.17%的方差,第二轴解释了26.78%的方差(图4B),土壤养分—酶活性相关排序为铵态氮>速效钾>全磷>全钾≈p H>有机磷>碱解氮>硝态氮;不同的是,老龄林阶段RDA 第一主轴解释了35.70%的方差,第二主轴解释了土壤养分—酶活性数据28.42%的方差(图4C),土壤养分—酶活性相关排序为p H>铵态氮>速效钾>硝态氮>碱解氮>全钾>全磷>有机磷。总的来说,不同恢复阶段内土壤养分与酶活性具有较强的相关性且土壤养分能较好地解释土壤酶活性。
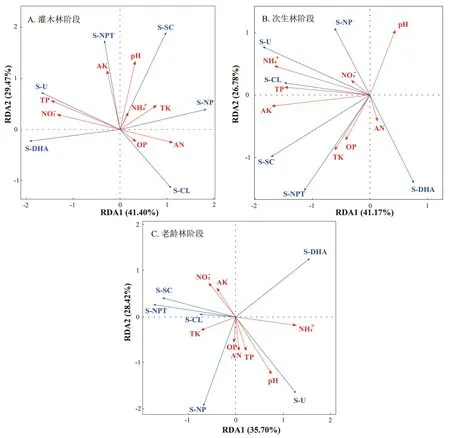
图4 退化喀斯特森林自然恢复阶段土壤养分及酶活性RDA 分析Fig.4 RDA analysis of soil nutrients and enzyme activity during the natural recovery stage of degraded Karst forests
3 讨论
3.1 退化喀斯特森林自然恢复进程中土壤养分含量变化特征
本研究中,土壤p H 值随着恢复进行不断上升,土壤整体呈现弱酸性至中性,这与森林中枯落物降解产生有机酸随着降水进入土壤有关[28];氮元素是植物生长发育不可或缺的重要营养元素,其中,土壤碱解氮可以反映土壤供氮能力[29];土壤中铵态氮可直接与光合作用的产物有机酸结合,形成氨基酸,进而形成其他含氮有机物,是土壤中重要的养分指标[30];土壤中硝态氮需要在土壤中还原成铵态氮后才能被植物吸收利用,所以土壤硝态氮也是衡量土壤养分的重要指标[30]。随着恢复进行,土壤铵态氮含量呈降低趋势,硝态氮和碱解氮含量从灌木林阶段到次生林阶段具有显著差异(P<0.05)。铵态氮是土壤中的重要氮元素来源,所以铵态氮随着恢复阶段的下降可能是影响老龄林阶段植被恢复的重要因素。土壤钾含量可以表征植物对钾的吸收与利用情况[31],在本研究中,土壤中速效钾整体变化差异不显著,全钾在前两个恢复阶段存在显著差异性,全钾是演替初期影响植被恢复的一个重要指标,演替后期由于植被恢复情况较好,地下枯落物层较厚,影响基岩风化从而释放钾元素,所以在本研究中全钾在恢复中后期差异不显著[32-33]。土壤磷是植物生长的重要营养元素之一[34],土壤有机磷可以表征植物对磷元素的吸收量,本研究中,土壤有机磷随着恢复具有先上升后下降的趋势,土壤全磷含量随着恢复进行显著上升,对照《第二次全国土壤普查土壤养分分级标准》,总体上,研究地全磷含量处在六级(<200 mg·kg—1)急缺水平[35],尽管植物从土壤中获取的全磷随恢复阶段呈现上升趋势,但是总体来说,磷元素的缺乏仍然是该地区植被生长的主要限制性因素。后续研究应该关注这种低磷限制对植被恢复的重要影响作用,总体来说,土壤中氮、磷、钾元素含量在不同恢复阶段存在显著性差异,这是由于自然恢复阶段的进行,地上植被类型发生了改变,地上枯落物的量也存在着不同,直接导致不同恢复阶段枯落物释放的氮、磷、钾元素在不同恢复阶段森林中存在差别[36],这说明喀斯特自然恢复阶段显著影响了土壤中氮磷钾元素的变化。土壤养分的主成分分析显示,灌木林阶段全磷、硝态氮、速效钾、有机磷、p H 值是土壤养分的主要决定因子;次生林阶段速效钾、全磷、硝态氮、铵态氮、p H 值是土壤养分的主要决定因子;老龄林阶段速效钾、碱解氮、全磷、铵态氮、有机磷是土壤养分的主要决定因子。可见,3个自然恢复阶段土壤养分主要决定因子存在差异,后续关于喀斯特自然恢复的研究应着重关注每一阶段指标变化的原因。
3.2 退化喀斯特森林自然恢复进程中土壤酶活性变化特征
土壤酶参与了森林生态系统中重要的生化过程,对土壤养分的释放和循环具有重要作用[37-38],其受到地表耕作方式、植物根系、土壤养分、水热条件等环境因子的影响[39-42]。本研究中,脲酶随着恢复的活性变化存在显著性差异,说明喀斯特森林采取自然恢复手段能显著提高该地区土壤脲酶的活性水平[43];土壤脲酶、中性蛋白酶、脱氢酶在恢复前期与恢复后期存在显著性差异,其他3种土壤酶活性在不同恢复阶段间均无显著性差异,说明土壤脲酶、中性蛋白酶、脱氢酶受不同恢复阶段影响较大,这是由于随着恢复阶段的进行,土壤养分、地上植被类型、植物根系、小气候等产生差异性改变,间接影响土壤酶活性,这与兰雪等人的研究相似[44]。本研究中纤维素酶活性随着恢复进行活性存在下降的趋势,可能的原因是:随着恢复进行,林分中的树种较多具有革质叶片,且具有发达的角质层,含有较多的难降解的木质素和鞣质等物质,不利于凋落物的分解[45-46]。可见,喀斯特自然恢复显著改变了土壤脲酶、中性蛋白酶、脱氢酶的活性。主导因子分析可知,在恢复进行到灌木林阶段,土壤脱氢酶、中性磷酸酶、蔗糖酶和中性蛋白酶是该阶段的土壤酶活性的主要决定因子;次生林阶段主要决定因子是土壤脲酶、蔗糖酶、脱氢酶、中性磷酸酶和中性蛋白酶;土壤脱氢酶、蔗糖酶、脲酶和中性磷酸酶是老龄林阶段土壤酶活性的主要决定因子。由于恢复阶段的不同,凋落物层的酶活性和酶种类也会随之发生改变,从而导致不同恢复阶段土壤酶活性的主要决定因子存在差异,这与前人的研究相同[47]。主成分分析结果表明,土壤脱氢酶、蔗糖酶是表征研究区土壤酶活性的重要指标,在不同恢复阶段中土壤蔗糖酶与脱氢酶均具有较好的解释度,说明不同阶段土壤中有某些特异性微生物群对分解恢复过程中凋落物的氮、磷等利用性较高[48]。土壤脲酶在3个恢复阶段均存在显著性差异,土壤蔗糖酶、纤维素酶、脱氢酶不受恢复阶段的影响,土壤中性蛋白酶、中性磷酸酶受恢复初期影响较大。说明喀斯特不同自然恢复阶段对于部分酶活性的影响较大,后续研究应重点分析这种差异变化的主导因素。
3.3 退化喀斯特森林3个自然恢复进程中土壤养分含量与酶活性关系
土壤酶能改善土壤质量,同时土壤理化性质也能给土壤酶提供酶促反应的环境和底物,进而对酶活性产生影响。土壤蔗糖酶是评定土壤肥力的良好指标[49],它能促进腐殖质、有机质等物质的转化,进而促进养分的释放以及循环,在本文研究的3个自然恢复阶段中,灌木林阶段的土壤蔗糖酶活性受到土壤p H 值、有机磷影响较大;随着恢复的进行,当先锋乔木在样地内占据主导地位,由于光照、水分条件、凋落物等环境条件的改变,此时次生林以及老龄林中土壤蔗糖酶不再受土壤p H 值和有机磷的影响,与全钾、速效钾、铵态氮呈强正相关。研究发现,土壤蔗糖酶与总氮、总磷含量具有极显著的相关性[50],本研究中土壤蔗糖酶整体上受土壤有机磷、全钾、p H 值、铵态氮等的影响较大,与前人研究相似。中性磷酸酶能促进凋落物分解形成植物所需的磷元素[51],但是,在本研究中,随着恢复阶段的变化,灌木林阶段土壤中性磷酸酶活性与碱解氮具有强正相关关系;而次生林阶段和老龄林阶段中性磷酸酶分别受土壤碱解氮、有机磷元素的影响。总体上,中性磷酸酶与土壤碱解氮、有机磷养分间具有较为密切的相关性,这可能是因为中性磷酸酶活性直接促进了土壤养分的释放和有效化,从而增加土壤中碱解氮、有机磷养分的占比[52-53]。在灌木林阶段,土壤纤维素酶与有机磷具有强正相关关系,当恢复阶段到达次生林以及老龄林阶段,此时的纤维素酶活性受土壤磷元素、氮元素、钾元素作用较大。纤维素酶能降解凋落物中的纤维素生成纤维素二糖,能有效地促进土壤中养分的转化,在一定范围内纤维素酶能改善土壤养分的释放情况[54],这很好地解释了3个自然恢复阶段内纤维素酶与N、P、K 养分因子间较强的相关性。脲酶是一种含有镍元素的寡聚酶,能够专一地水解土壤中的尿素[55],从而促进氮元素的释放。研究结果显示,土壤脲酶在不同自然恢复阶段与氮元素具有较强相关性,这是由于脲酶与土壤有机质中的碳氮键发生作用,从而将尿素水解成土壤中的氨,从而提高土壤中氮元素的水平,这能很好地解释脲酶与含氮养分间具有的很强相关性。可见,脲酶、蔗糖酶、中性磷酸酶、纤维素酶在喀斯特不同自然恢复阶段土壤养分转化过程中发挥着重要作用。
4 结论
退化喀斯特森林不同自然恢复阶段显著影响了土壤养分及酶活性变化特征,随着恢复阶段的进行,土壤全磷、脲酶含量呈上升趋势,土壤铵态氮、全钾、碱解氮、土壤中性蛋白酶、中性磷酸酶等指标受自然恢复影响显著。在不同自然恢复阶段,土壤酶活性和养分的主要决定因子存在不同,土壤脲酶、蔗糖酶、中性磷酸酶、纤维素酶与氮、磷、钾养分间存在较强相关性。本文以退化喀斯特不同自然恢复阶段为研究对象,分析了土壤养分与酶活性在不同阶段的变化特征、主导因子以及各阶段内土壤养分与酶活性的相关性,为退化喀斯特森林自然恢复提供了一定的科学理论支撑。
猜你喜欢
华北水利水电大学学报(自然科学版)(2020年6期)2021-01-29
生物技术通报(2020年12期)2020-12-21
腐植酸(2020年1期)2020-11-29
农业环境科学学报(2020年10期)2020-11-11
阅读(低年级)(2020年8期)2020-11-06
乡村地理(2019年2期)2019-11-16
————水溶蚀岩石的奇观
家教世界(2019年4期)2019-02-26
浙江理工大学学报(自然科学版)(2018年6期)2018-11-07
湖泊科学(2015年1期)2015-05-06
文化月刊·下旬刊(2014年6期)2014-08-28
