
干旱胁迫对滨海沙地主要造林树种水力结构的影响
2024-01-20 02:46张马啸黄小艳邹志广李树斌周丽丽
西北林学院学报 2024年1期
张马啸,黄小艳,邹志广,李树斌,潘 辉,周丽丽
(1.福建农林大学 资源与环境学院,福建 福州 350002;2.福建农林大学 林学院,福建 福州 350002;3.闽江学院 经济与管理学院,福建 福州 350108;4.闽江学院 地理与海洋学院,福建 福州 350108)
随着全球气温不断升高,季节性干旱频发[1]。降水不均及雨热不同期会使树木面临不同程度的水分亏缺,引起树木木质部水势下降和张力增加,从而形成不同程度的空穴和栓塞化,最终影响树木的生长及生存[2]。叶片是树木与外界环境接触最直接的器官,研究表明叶片水势是反映树木水分亏缺最敏感的生理指标之一,可用来判断植物受干旱胁迫的程度和抗旱能力的高低。一般认为叶片水势越低,树木的吸水能力越强[3]。植物在不同的水力条件下会有不同的形态结构变化和水分运输供给策略,水力结构也是衡量植物水分适应的重要指标,其中导水率(Kh)和比导率(Ks)是水力结构的重要指标,茎段木质部的导水率越大表明导水能力越强,比导率越大表示该茎段的输水效率越高[4]。在干旱环境下,由于水分的亏缺导致树木不同器官的木质部因导水率下降而产生不同程度的空穴和栓塞,形成水力疏导障碍。为适应不同程度的干旱逆境,树木在长期进化过程中形成了不同的水分运输策略。
近50 a来福建省年平均气温上升了1 ℃左右,呈明显的增暖趋势[5],加之福建滨海土壤多为潮积或风积沙壤土,养分和水分状况较差,雨热不均使得滨海沙地树种经常面临干旱风险。长期以来,木麻黄作为沿海省份的防护林树种,单一化严重,林分结构简单,生态系统稳定性差[6]。目前关于滨海沙地树木的研究主要集中在引种造林、养分吸收及凋落物养分含量、碳氮磷化学计量特征、叶绿素荧光参数动态、生物量分配格局和防护效果等方面[7-11],对滨海沙地造林树种响应干旱逆境的研究还很匮乏。不同滨海沙地树种如何通过调整导水率等水力结构特征来适应干旱环境?不同树种不同器官的水力结构分配有何差异?目前还不清楚,而这些问题的回答对于揭示不同滨海沙地造林树种在干旱胁迫下的水分生理及水分运输机制具有重要的理论意义,对于选育适合滨海沙地生长的耐旱树种及合理配置也具有重要指导意义。
鉴于此,选择滨海沙地的主要造林树种木麻黄(Casuarinaequisetifolia)、尾巨桉(Eucalyptusurophylla×E.grandis)、马大相思(Acaciamangium×A.auriculiformis)和黑木相思(A.melanoxylon)为研究对象(1年生幼苗),通过设置不同干旱胁迫程度的处理,比较不同树种不同叶位叶片水势的差异,分析不同树种不同器官导水率及比导率的变化特征,探讨不同滨海沙地主要造林树种响应干旱胁迫的水力学机制,为滨海沙地树种的选育、管理及合理配置提供科学依据。
1 材料与方法
1.1 供试材料及试验设计
2021年9月,在福建农林大学林学实验中心强化育种温室大棚内,选取福建省东山县赤山林场培育的1年生木麻黄、尾巨桉、马大相思和黑木相思4种实生苗作为试验材料,经沙床缓苗后选取健康幼苗,移入统一规格的塑料花盆中(高23 cm、上口径25 cm、下口径17 cm)进行驯化,每盆培养基质5 kg(土砂质量比2∶1)。木麻黄、尾巨桉、马大相思和黑木相思平均苗高分别为1.51±0.10、0.67±0.12、0.79±0.25 m和0.99±0.18 m,平均地径依次为10.59±1.01、9.29±1.79、10.52±1.42 cm和7.29±1.26 cm。
缓苗3个月后,于12月1日进行不同程度的干旱胁迫处理。根据王凯等[12]的研究,设置适宜水分作为对照(W0),轻度胁迫(W1)、中度胁迫(W2)和重度胁迫(W3)4个梯度水分处理,其中W0、W1、W2、W3分别为最大田间持水量(50.07%)的80%、60%、50%和40%。每个处理设置5个重复,共80盆。试验前选取不同种类苗高、地径基本一致的苗木,并用土壤水分测定仪进行水分监测。当每盆中土壤均自然干旱到规定梯度时,正式开始试验,试验过程中于每日18:00补充当天失去的水分,使其土壤水分保持在相应的干旱梯度内。通过观察,随着重度胁迫时间的延长,木麻黄的中、下部叶逐渐脱色,由绿色变黄直至变白,底部叶开始凋落;尾巨桉和马大相思的底部叶片脱色并大量凋落;黑木相思的形态变化最慢,重度干旱胁迫10 d左右叶片才仅有微微卷曲且并无变色,20 d时各部位叶片均开始卷曲,较为短小的叶片开始凋落。因此,我们在干旱胁迫20 d后对苗木收获测定[13]。
1.2 指标测定
1.2.1 水势的测定 苗木收获后采集不同叶位(底部、中部和顶部)的叶片进行水势测定,测定仪器为WP4-T露点水势仪(decagon devices,USA),使用前先用0.5 mol·L-1的KCl溶液对水势仪进行校准[14],然后将叶片剪碎铺满样品杯中进行测定。
1.2.2 导水率及比导率的测定 采用栓塞测量系统(XYL'EM-Plus,法国)和王明浩等[15]的改良冲洗法测定样品导水率。收获苗木后,将其根部浸入超纯水中并用黑色塑料袋罩住迅速带至实验室,以防水分散失。分别截取长度约4 cm的根、茎和枝条,剥掉韧皮部后用无油生料胶带缠住木质部,防止漏水,将木质部吸水端套入连接仪器的塑料管中,在低压(0.01 MPa)下,用0.03 mol·L-1KCI溶液冲洗并测定其导水率,用万分之一天平称量2 min内通过该样段的水量。用直尺和游标卡尺测量茎段长度、横截面直径和心材直径,计算出茎段边裁横截面积。导水率(Kh,g·m·MPa-1·min-1)采用水流量与样本长度的乘积再除以压力来计算,比导率(Ks,g·m·MPa-1·min-1·cm-2)采用导水率除以样段边裁横截面积来计算[16]。
1.3 数据分析
采用SPSS 22.0软件对数据进行统计分析。对同一树种不同干旱胁迫程度、同一干旱胁迫程度不同树种之间的各项指标分别采用单因素(one-way ANOVA)方差分析法,对树种、叶位和干旱胁迫程度对叶片水势的影响采用三因素方差分析法,对树种、器官和干旱胁迫程度对导水率和比导率的影响也采用三因素方差分析法。表中数据为平均值±标准误。
2 结果与分析
2.1 干旱胁迫对不同树种不同叶位叶片水势的影响
由表1可知,树种、叶位和干旱胁迫程度及其三者的交互作用对叶片水势的影响均达极显著水平(P<0.01)。由表2可知,干旱胁迫下4个滨海树种各叶位的叶片水势均低于对照,但干旱胁迫程度对叶片水势的影响会因树种和叶位的不同而存在明显差异。黑木相思各叶位的水势均表现为随干旱胁迫程度的增加先减后加再减的趋势,各叶位水势在中度干旱胁迫较轻度干旱胁迫时都有小幅增加,分别为底部叶增长11.60%、中部叶增长25.21%、顶部叶增长18.47%;木麻黄中部叶和顶部叶的水势表现出与黑木相思一致的规律,都表现为在中度干旱胁迫时有所增长,但其底部叶的水势则表现为随干旱程度的加深持续下降;尾巨桉中部叶和顶部叶的水势亦表现为波动变化的特征,具体为中度胁迫时的水势比轻度胁迫分别低54.13%和48.22%,重度胁迫时的水势比中度胁迫分别高46.63%和53.93%,且中部叶和顶部叶在重度胁迫时的叶片水势均高于中度和轻度,底部叶在重度干旱时水势下降幅度最大,比中度干旱低1 065.58%;马大相思底部叶和中部叶的水势表现为波动变化,底部叶的水势在中度干旱时高于轻度干旱的15.90%,重度干旱时低于中度干旱的2 421.82%,中部叶水势在中度干旱时与轻度相比下降了4.10%,重度干旱时与中度相比增加了6.71%,顶部叶的水势表现为由轻度到中度再到重度逐渐增高。

表1 树种、叶位和干旱程度对4种树木叶片水势的三因素方差分析Table 1 Three-factor variance analysis of tree species,leaf position and drought degree on water potential of four trees
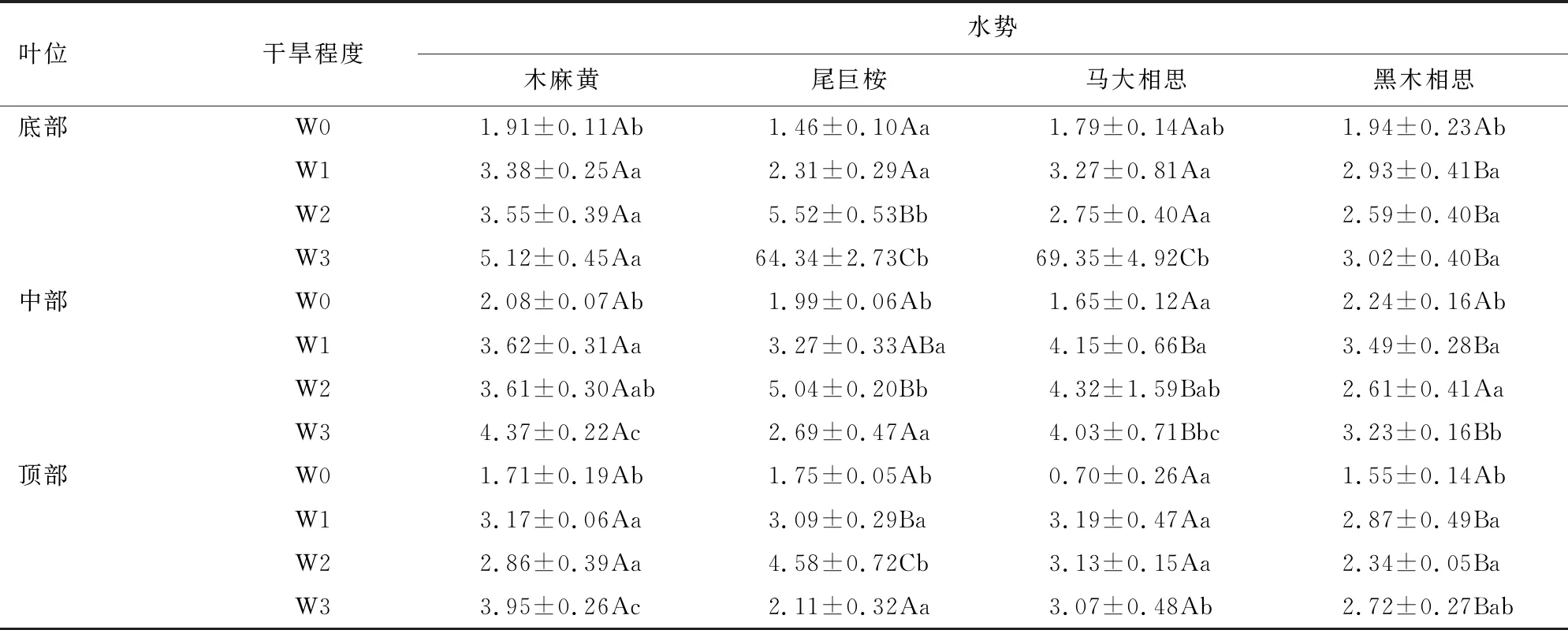
表2 干旱胁迫对不同树种、不同叶位叶片水势的影响Table 2 Effects of drought stress on leaf water potential of different tree species and different leaf positions
同一干旱胁迫程度不同树种之间的叶片水势存在明显差异,对照条件下,马大相思的顶部叶显著高于其他3个树种(P<0.05);中度干旱胁迫时,尾巨桉的底部和顶部叶水势显著低于其他3个树种;重度干旱胁迫时,尾巨桉和马大相思的底部叶水势显著低于其他2个树种。
2.2 干旱胁迫对不同树种不同器官导水率的影响
由表3可知,导水率受树种、器官和干旱程度及其三者交互作用的影响均达极显著水平(P<0.01)。由表4可知,干旱胁迫下不同器官导水率的变化不同,木麻黄根的导水率在不同程度的干旱胁迫时均高于对照,轻度、中度和重度分别高于对照3.77、1.48倍和1.23倍,在轻度胁迫下导水率最高,其他3个树种在遭受干旱胁迫时根的导水率均低于对照。干旱胁迫条件下4个树种的茎导水率均表现为随干旱胁迫程度先升后降的趋势,其中木麻黄和黑木相思茎的导水在干旱胁迫时均高于对照,在重度胁迫条件时这2个树种茎的导水率仍高于对照1.47倍和6.10倍;木麻黄枝的导水率均低于对照,黑木相思枝的导水率均高于对照,且表现出升-降-升的规律,重度胁迫时黑木相思枝的导水率比中度胁迫提高了57.53%。干旱胁迫对各器官导水率的影响因树种不同而差异明显,尾巨桉和马大相思根的导水率表现为随干旱胁迫程度的增加而下降,木麻黄根的导水率表现为轻度干旱胁迫时升高,中度和重度干旱胁迫时持续下降,而黑木相思根的导水率则表现出在轻度和中度干旱时下降,重度干旱胁迫升高;4个树种茎的导水率均表现为随干旱胁迫程度的增加呈升-降-降的趋势;枝的导水率亦表现为轻度干旱胁迫时升高、中度和重度连续下降的特点,但木麻黄枝的导水率则表现为连续下降的趋势。
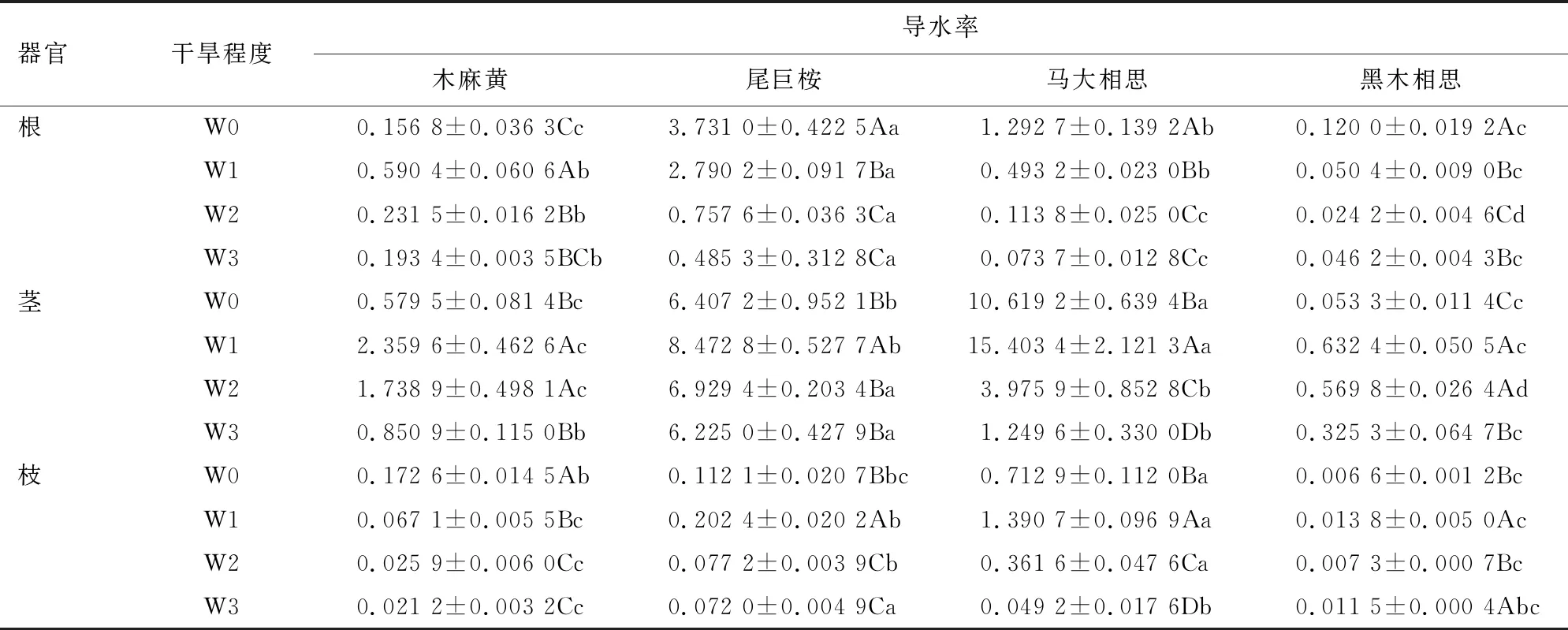
表4 干旱胁迫对不同树种不同器官导水率的影响Table 4 Effects of drought stress on hydraulic conductivity of different organs of different tree species g·m·MPa-1·min-1
同一干旱胁迫程度不同树种之间的导水率也存在显著差异。在对照条件下,尾巨桉根的导水率、马大相思茎和枝的导水率显著高于其他3个树种;轻度干旱胁迫与对照条件的规律相同;中度干旱胁迫和重度干旱胁迫时,尾巨桉根和茎的导水率显著高于其他3个树种。
2.3 干旱胁迫对不同树种不同器官比导率的影响
比导率受树种、器官和干旱程度及其三者交互作用的影响均达极显著水平(P<0.01)。由表5可知,干旱胁迫条件下不同器官比导率的变化不同,马大相思和黑木相思根比导率在干旱条件下均低于对照,木麻黄和尾巨桉根比导率在轻度干旱胁迫时分别高于对照578.81%和10.85%;4个树种的茎比导率在干旱胁迫条件下均表现为先增后降,在轻度干旱胁迫时分别比对照增加161.79%、6.70%、20.09%和1 553.12%,木麻黄和黑木相思茎的比导率在重度干旱胁迫下高于对照,4个树种中马大相思茎比导率重度胁迫下较对照降低79.16%;4个树种枝的比导率在轻度干旱胁迫下都高于对照,其中马大相思增加最多(61.62%),木麻黄和黑木相思枝比导率则表现为在重度胁迫时高于对照,分别高于对照2.45倍和1.72倍。
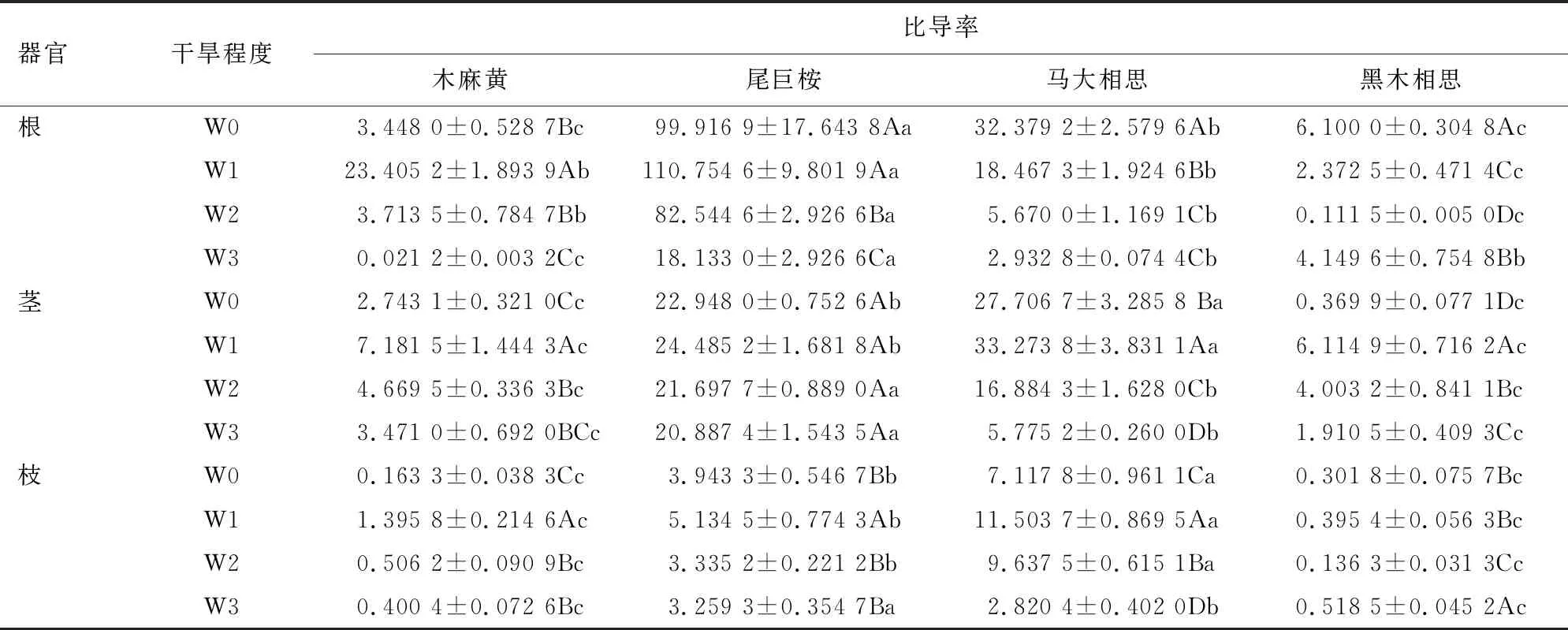
表5 干旱胁迫对不同树种不同器官比导率的影响Table 5 Effects of dought stress on conductivity of different organs of different tree species g·m·MPa-1·min-1·cm-2
干旱胁迫程度对各器官比导率的影响因树种不同而不同。轻度干旱胁迫条件下,尾巨桉根、马大相思茎和枝的比导率显著高于其他3个树种。在中度干旱胁迫和重度干旱胁迫条件下,尾巨桉根和茎的比导率最高;马大相思枝的比导率在中度干旱时最高,黑木相思枝的比导率则在中度干旱时最低,重度干旱时升高甚至高于对照。
3 讨论
叶片水势是反映植物体内水分亏缺程度的重要指标,叶片水势越高,说明植物供水条件越好。本研究发现,4个树种干旱胁迫条件下各叶位的水势均低于对照,轻度干旱胁迫时4个树种各叶位叶片水势均低于对照,中度胁迫时马大相思和黑木相思底部叶、木麻黄和黑木相思中部叶、木麻黄马大相思和黑木相思的顶部叶水势较轻度干旱时升高,重度干旱胁迫时尾巨桉和马大相思的中部和顶部叶水势较中度胁迫时升高,其余水势在干旱条件下均呈下降趋势,这与庞云龙等[17]的研究结果一致,说明干旱胁迫促使植物通过降低自身叶片的水势来调节体内水势梯度,从而保证水分从根部向地上部分运输。然而,不同叶位叶片水势对干旱胁迫的响应不同,本研究中随干旱胁迫程度的增加4个树种的叶片均呈现不同程度的卷曲、折叠,以减少水分辐射和水分丧失,尤其到重度干旱胁迫时,尾巨桉和马大相思底部叶片水势显著降低至-64.34 MPa和-69.34 MPa,底部叶片开始大量枯死并脱落,这与大部分研究结果一致[18-19],说明尾巨桉和马大相思在重度干旱胁迫时,会采取主动牺牲底部的劣势叶片、以保证中部和顶部叶片水分供应的生长策略。本研究发现,黑木相思在遭受中度干旱胁迫时各叶位水势均高于其他3个树种,尤其底部叶和顶部叶水势在整个干旱胁迫过程中并没有显著差异,表现出“高水势延迟脱水”的特性,其较好的抗旱性可能与黑木相思的叶片较细较长较小、蒸腾速率弱有关[20]。木麻黄中部和顶部叶和黑木相思各部位的叶片水势表现为中度胁迫高于轻度胁迫,尾巨桉和马大相思中部和顶部的叶片水势却表现为重度胁迫高于中度胁迫,这与李继文等[21]的研究结果一致,可能存在2种原因,一方面植物在轻度干旱胁迫时通过降低叶片水势来促进根系对水分的吸收,而当植物适应干旱后,中度干旱胁迫时叶片水势会有所回升;另一方面,可能由于干旱导致叶片水势降低较多,叶片与大气之间出现较大水势差驱动叶片在程度更深的干旱胁迫中吸取大气中的水分造成的[22]。
导水率是指通过离体茎段的水流量与引起水流动的压力梯度的比值,导水率越大,说明其导水能力越强。本研究中,4个树种茎、木麻黄根和尾巨桉枝、马大相思枝、黑木相思枝的导水率在轻度干旱胁迫时均高于对照;中度干旱胁迫时4个树种各器官的导水率均低于轻度;重度胁迫下除黑木相思根和枝的导水率外,其余均继续降低。根系是植物直接感知土壤干旱胁迫的器官,响应土壤水分变化也最敏感[23],在干旱胁迫条件下,木麻黄、尾巨桉和马大相思根的导水率表现为随干旱胁迫的增加而降低,这与North等[24]的研究结果一致。水分不足导致根系内、外皮层细胞木栓化,从而引起细胞质膜上水通道蛋白发生变化而降低导水率。黑木相思在遭受重度干旱逆境时根的导水率高于中度胁迫,结合其叶片水势在干旱胁迫下的变化规律,说明黑木相思在遭受严重干旱胁迫时会采取增加根部导水率、降低叶片水势的方式度过逆境期。木麻黄根的导水率在轻度干旱胁迫时明显增加,可能由于对照的土壤水分过于充足,使根系缺氧而影响水通道蛋白的活性[25],从而使对照根导水率明显低于轻度干旱胁迫。尾巨桉、马大相思和黑木相思在轻度干旱胁迫时根的导水率低于对照,而茎和枝的导水率却高于对照,表明这3个阔叶树种在遭受轻度干旱胁迫时通过降低根导水率、增加茎和枝的导水率的方式来满足植物对水分的需求。
比导率是指通过单位茎段的导水率与边裁横截面积的比值,比导率越高,单位有效面积的输水能力越强,表明输水效率越高[26]。除马大相思和黑木相思的根外,4个树种各器官在轻度干旱时的比导率均高于对照,说明面对轻度干旱胁迫时苗木首先通过增大其木质部输水效率来保证苗木生长的水量平衡。与徐茜等[27]报道的干旱胁迫下胡杨比导率总体增加的结论一致,胡杨幼株生长需水量大,干旱胁迫下,木质部增大其比导率来保持“根系吸水-根部输水-叶片蒸腾”的水量平衡。本研究中不同树种各器官(遭受重度干旱胁迫下的黑木相思的根和枝除外)随干旱胁迫程度的增加,比导率均呈下降趋势,可能与干旱胁迫导致蒸腾速率下降有关[28]。4个树种(除马大相思和黑木相思的根外)各器官在遭受干旱胁迫时,首先采取增大木质部的输水效率以保证地上枝叶对水分的需求,然而随干旱胁迫程度的增加,各器官的比导率却在下降,这可能会导致不同器官发生栓塞现象。可见,不同树种的叶片水势和各器官的导水率、比导率对干旱胁迫的响应不同,树种之间对干旱逆境的敏感性和响应策略有明显差异。
4 结论
4种1年生滨海沙地树种对干旱逆境的敏感程度不同,应对不同程度的干旱胁迫所采取的适应策略也存在差异。轻度干旱胁迫下,马大相思和黑木相思通过降低根的导水率和比导率、提高茎和枝的导水率来适应干旱逆境,木麻黄则采取提高根和茎的导水率、降低枝的导水率来适应干旱;中度和重度干旱胁迫下,木麻黄、尾巨桉和马大相思的茎和枝的导水率和比导率均呈下降趋势,且尾巨桉和马大相思会选择牺牲底部叶片、提高中上部叶水势,而黑木相思则采取提高根和枝的导水率和比导率、延迟叶片水势下降以适应中度干旱逆境。本研究结果有助于揭示不同滨海沙地树种响应干旱逆境的水分运输策略,对滨海树种的选育、造林配置及生态功能的优化提升具有重要意义。今后应加强叶片形态特征、气孔导度以及木质部解剖结构与耐旱性的研究,从更深层次揭示不同滨海树种对干旱胁迫的适应机制。
猜你喜欢
西北农业学报(2023年11期)2023-10-31
农业工程学报(2022年7期)2022-07-09
水利技术监督(2017年3期)2017-06-09
文学少年(小学版)(2017年4期)2017-04-18
幸福·健康版(2016年4期)2016-05-04
水利科技与经济(2016年3期)2016-04-09
水利科技与经济(2016年4期)2016-04-09
水利科技与经济(2016年2期)2016-04-09
环球时报(2016-03-16)2016-03-16
福建林业(2015年2期)2015-04-09
