
基于多光谱无人机的不同放牧策略对草地叶面积指数变化动态解析
2024-01-23 12:37刘嘉慧陈荣昊
热带生物学报 2024年1期
刘嘉慧,余 瑞,王 有,李 欣,陈荣昊
(1. 海南大学 生态与环境学院,海口 570228; 2. 中国地质调查局海口海洋地质调查中心,海口 571127)
叶面积指数(Leaf area index,LAI,记作ILA)是草地生态系统重要的状态变量[1-2],它又称叶面积系数,指单位草地面积上植物叶面积的总和,通常用于描述植被覆盖的密度和茂密程度,数值越大代表植被覆盖越密集[3]。它是反映草地生长状态的重要参数,其变化能够直接或者间接揭示牧草在不同放牧方式下的变化规律。LAI 也用来描述植被的生长状况和生态功能,草地的许多生物过程都受到叶面积指数的影响,草地叶面积指数能够直接反映草地生长状况和生产力水平,并且能够指示草地生长状况[4]。因此,叶面积指数常用于研究草地群落组成与生态系统之间的反馈机制过程[5],以及在小空间尺度上衡量冠层生物量和异质性。在多草种混合草地草原上,LAI 经常被用来监测碳平衡和评价草的质量和生产力[6-8]。近几十年来,无人机结合植被指数的经验模型法使其在草地遥感监测方面成为一项极具前途的技术[9]。经验模型法认为叶面积指数与植被指数之间存在函数关系,根据此种函数关系可以进行叶面积指数反演。二十世纪七十年代,Bunnik 利用遥感技术成功提取出叶面积指数,奠定了遥感影像反演LAI 的基础[10]。无人机在图像质量、传感效率和运营成本之间都可以实现适当的平衡。各级草地管理单位及研究人员建立了众多的草地状况监测站点,结合遥感或无人机技术,能够提供大范围且精确的草地状况基础数据[11]。基于主动多光谱冠层传感器,Li 等[12]提出一种利用无人机主动传感估算水稻LAI 的新方法,相较于辐射传输模型的不确定性及辐射信号携带的有限信息,无人机所制备的植被指数是根据光谱信号的目标信息计算出来的,更加能够准确地反映出短时间内草地受到放牧干扰的生长情况。
放牧方式直接或者间接干扰牧草生长,代表性的放牧方式有轮牧和连续放牧。适当的轮牧充分利用牧草的生物学特性,降低了牲畜的选择性采食行为与践踏对草地的干扰强度,使草地的生产力在整个生产过程能够达到最大化[13-14]。然而,过度轮牧会造成地表被过度践踏而使草地裸露并导致裸露地表破碎化,改变土壤的物理性状,从而抑制草地植物的生长并降低草地物种多样性,改变草地生态系统原有的结构和功能[15-16]。因此,过度集约利用草原是许多植物物种消失的主要原因。而中等强度轮牧可以有效地缓和由于牲畜啃食造成的草地叶片减少而引起的光合作用减少,缓解草地有机物的急骤减少[17],促进草地牧草的生长,草地叶面积指数反而因放牧升高,这表明在适当强度的轮牧干扰下,有利于维持草地叶面积处于较高状态,草地生物的多样性和丰富度得以维持甚至有所增加。轻度的连续放牧由于食物来源的限制,在某一特定区域有序活动,可以加快草地无效掉落物的分解,增加土壤氮和有机物的含量,减少土壤硝酸盐的可提取磷进而降低土壤污染[18],从而促进草地牧草生长。彭祺等[19]却认为连续放牧是一种无计划、粗放的经营方式,过度地连续放牧会导致草地生物多样性和稳定性降低,牲畜的过度踩踏效应会使土壤压缩从而导致草地生产力降低,质量下降,甚至引起草地不可逆的退化[20],但是连续放牧并不意味着家畜对个体的连续无休止采食,通过轻度或者中度连续放牧也能使草层保持较高的有机物积累水平,刺激并促进草地的生长,维持草地的稳定[21],适当地将食草动物引入草原能对植物群落的富集和多样性有积极影响,有助于保持和增加其植物多样性,使草地叶面积指数保持稳定。热带地区不同放牧方式与放牧强度下,叶面积指数的动态变化机制是怎样的,值得深入研究。
本研究按照不同放牧方式(轮牧和连续放牧)、不同放牧强度(重度、中度和轻度)的放牧策略进行放牧试验,利用多光谱无人机对放牧期间的草地LAI 按日尺度进行监测,获得每日放牧前和放牧后的多光谱影像,分析不同放牧策略对草地LAI 的影响,揭示不同放牧策略对草地叶面积指数影响的变化规律,在微观层面优选出抑制草地退化适应性管理策略。
1 材料与方法
1.1 研究区概况放牧实验区位于海南省文昌市锦山镇锦华村,草地主要由地毯草和狗牙根组成,该地区位于海南岛东北部,气候类型属热带海岛季风气候,每年11 月到次年4 月为干季,全年平均温度在22.5~25.6 ℃之间,年均降雨量在900~2 500 mm 之间[22],该地区温暖潮湿的气候和较高的降雨量,为畜牧业提供了充足的水源和气候条件。经实地走访调查和村委会提供的数据,发现村内家庭牧场普遍采取散养的放牧模式,散养牛的总数量为212 头。并且该村土地撂荒情况严重,位于锦华村西部的大片水田无人管理,出现水田和草地无序放牧的情况。
1.2 放牧实验设计按照不同放牧方式(轮牧和连续放牧)、不同放牧强度(重度、中度和轻度)的管理方式进行试验(表1)。放牧强度通过调节样地牛头数来控制,利用多光谱无人机对放牧期间的草地LAI 按日尺度进行监测。实验草地面积为0.39 hm2,分成3 个放牧样地:G1、G2 和G3,面积分别为0.11、0.11 和0.17 hm2,在3 个草地的周围再选取1 个样地作为不放牧草地NG,以便进行对照,将G1 作为轮牧的第一块样地,G2 作为轮牧的第二块样地,按照时间顺序依次进行5 d 的放牧,G3 作为连续放牧的样地,进行10 d 的放牧,放牧时间为每日8∶30—17∶30,其余时间将牛赶回牛 棚,同一个时间段保证放牧强度相同,放牧强度的设定参考Costa 等[23]对热带牧场放牧强度的分析和当地25 头·hm-2的生产实际,总共进行3 次放牧试验(图1)。

图1 放牧实验设计图

表1 放牧实验设计
1.3 数据获取
1.3.1 叶面积指数实测数据采用LAI2200-C 植被冠层分析仪观测LAI。LAI2200-C 是一种常用于测量植被叶面积指数的仪器,其工作原理是基于直接测量植被冠层中的光线透过程度,该仪器内部还装有全球定位系统(GPS)模组。在稳定的天空条件下,对研究区的草地按照对角线的方式选择两条线路做好标记,以1.5 m 的等距离测量,进行测量时使用270°视野遮盖帽,将传感器放在植被冠层上方,所得到的天空亮度记为A值,然后在冠层下方读取4 个B值。测量完成后,LAI2200-C 会自动将测量结果保存在其内部存储卡中,将仪器与计算机连接来读取、导出测量结果,利用LAI2200-C 方法计算ILA,计算公式为:
式中,ILA是叶片(所有遮光体)面积指数,是i环的平均接触频率,Wi是i环的权重因子。
平均接触频率是透射率和路径长度的函数,可以通过n个Above/Below 的观测值计算获得:
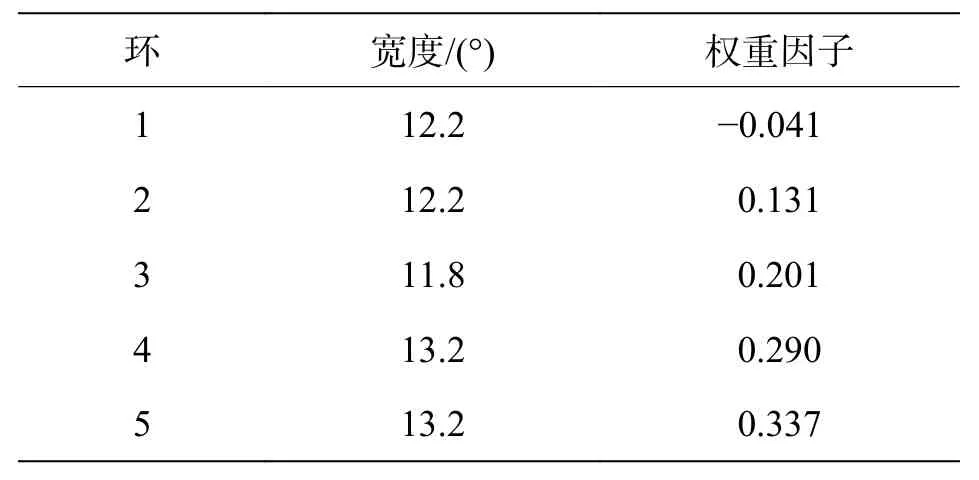
表2 环宽度和权重因子
1.3.2 无人机影像本研究利用无人机对不同放牧策略下的草地叶面积指数按日尺度进行监测。所用的DJI 精灵4 多光谱版无人机,可以拍摄5 个波段的图像:蓝光、绿光、红光、红边和近红外,提供高精度的定位信息。本试验利用无人机划定飞行区域,设置93.9 m 的飞行高度,飞行速度14.6 m·s-1,等时间间隔2.2 s 拍照,每日在早上牛入栏前和下午牛出栏后进行飞行,以保证获得草地5 cm分辨率的可见光成像(JPEG 格式)和多光谱成像(TIFF)。
1.3.3 畜牧行为采用目标取样法在轮牧草地和连续放牧草地中分别选择具有健康状况良好且具有代表性的一头牛,人工目视全天跟踪观测。图2 为牛在各种行为下的样本图像,利用连续记录法按每5 s 一个频次,采集在放牧时间内(8∶30—17∶30)放牧牛采食、站立、慢走、躺卧和反刍的运动行为时间。
1.4 数据处理
1.4.1 无人机影像制备NDVI无人机影像处理和制备NDVI 均基于大疆智图完成。将影像导入到大疆智图软件中,进行包括相机标定、畸变校正、图像匹配、飞行姿态校正等预处理操作,对原始影像进行正射,将图像像素的灰度值转换为地物反射率或光谱反射率,并根据像元的位置计算出地物的空间分布,以消除地形起伏等因素对影像的影响,获得正射影像。选取最常用的植被指数归一化植被指数(NDVI)进行LAI 反演,反演过程参照Hassan 等[24]的研究,归一化植被指数NDVI 的计算公式为:
其中:Aref(NIR)和Aref(Red)分别表示NIR 波段和Red 波段的反照率。
将Qinc和Qref分别代表波段上的入射光量和反射光量,太阳光(入射光)照射到地面后发生反射,多光谱相机采集地面反射光线的信号并产生影像,而多光谱传感器则采集入射光线的信号,从而得出下列公式:
Qinc(NIR)和Qref(NIR)分别表示在NIR 波段上的入射光量和反射光量,Qinc(Red)和Qref(Red)则分别表示在Red 波段上的反射光量和入射光量,Lcam(NIR)和Lcam(Red)分别表示从NIR 和Red 波段的多光谱图像中获得的图像信号值,而Ls(NIR)和Ls(Red)则分别表示从NIR 和Red 波段的多光谱光强传感器获得的感光信号值,ρNIR和ρRed表示调节NIR 波段和Red 波段的调节图像信号与多光谱光强传感器信号之间相互转化的参数。
在使用NIR 波段相机作为标准时,其他波段的相机和多光谱光强传感器的感度都需要进行校准,需要将其他波段的相机和多光谱光强传感器的感度调整到与NIR 波段的相机和多光谱光强传感器相匹配。ρx作为调节图像信号与多光谱光强传感器信号之间相互转化的参数,校准参数分别为pLcamX和pLSX。通过校准可以定义:
可以得到:
因此,使用无人机影像计算NDVI 的公式如下:
1.4.2 植被指数反演归一化植被指数(NDVI)作为最常用的LAI 反演指数,利用了冠层反射或辐射中的红光和近红外波段信息。本研究通过多光谱无人机数据生产NDVI,在3 个不同放牧区域内分别选取30 个监测点,使用植被冠层分析仪实地测量草地叶面积指数(LAI),分析监测点所对应NDVI 与所测量LAI 的相关关系,以此制备整个研究区的LAI 数据。利用归一化植被指数(NDVI)估算LAI 的通常步骤是先NDVI 与LAI 的经验关系,并使用植被冠层分析仪的实地观测数据进行拟合,再利用拟合好的模型进行估算,表达NDVILAI 之间的经验关系L=f(x)主要有以下几种形式[25 - 26]:
式中,X为归一化植被指数NDVI,参数A、B、C 和D 为经验值,不同植被类型参数有所不同。
采用Origin 统计软件对LAI 和NDVI 进行关系拟合。将同一时间通过LAI-2200C 所测出来的LAI 与基于无人机遥感影像所制备的NDVI 进行一一对应,根据常用的经验模型,将NDVI 和LAI 进行组合,逐步拟合回归,依次输入模型方程,在进行拟合回归时,要考虑到NDVI 的变化对于LAI 变化的显著性程度大小,对于与LAI 没有显著性关系的模型方程,直接排除该模型。通过实测的数据依次对上述方程进行拟合,获得最佳拟合方程和给定系数后,便可将公式应用到遥感影像上来进行LAI 空间分布的制图。
2 结果与分析
2.1 基于经验模型的草地LAI 反演基于经验模型反演的LAI 具有较高的精度。选择73 个样点进行经验模型关系拟合,部分样点NDVI 和LAI 值如表3 所示。从图3(a)可以看出,NDVI与LAI 具有较好的相关性,当NDVI 接近0.8 时,NDVI 值趋于缓和,该LAI 反演模型的相关系数R2为0.88,P值小于0.01,表明用该模型拟合NDVI 和LAI 的效果较好。图3(b)对实测LAI 和经验模型模拟出来的LAI 进行了验证,相关系数R2=0.74,两组数据之间的整体偏差较小,当LAI 靠近1.5 时,反演结果会与实测值出现偏差,但在其他取值范围偏差较小,并且从整体来看,两者具有大致相同的变化趋势,表明该经验模型反演的LAI 误差较小,反演效果较好。
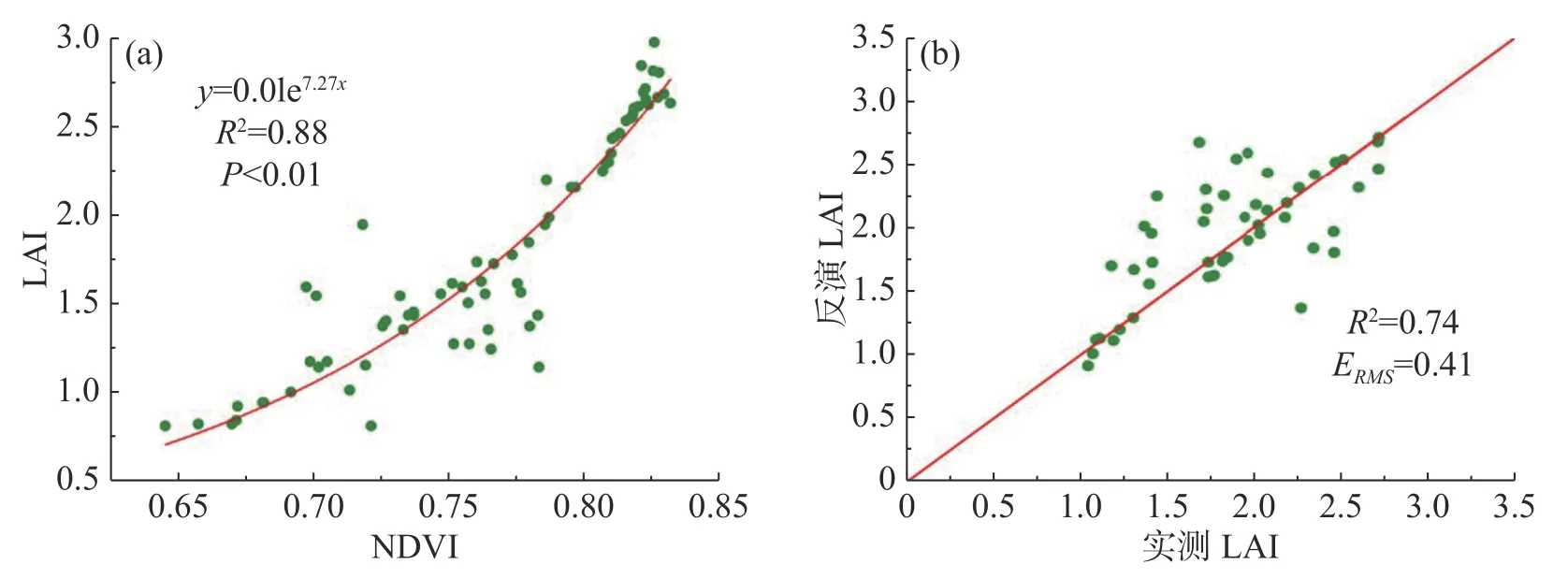
图3 基于经验模型的草地LAI 反演
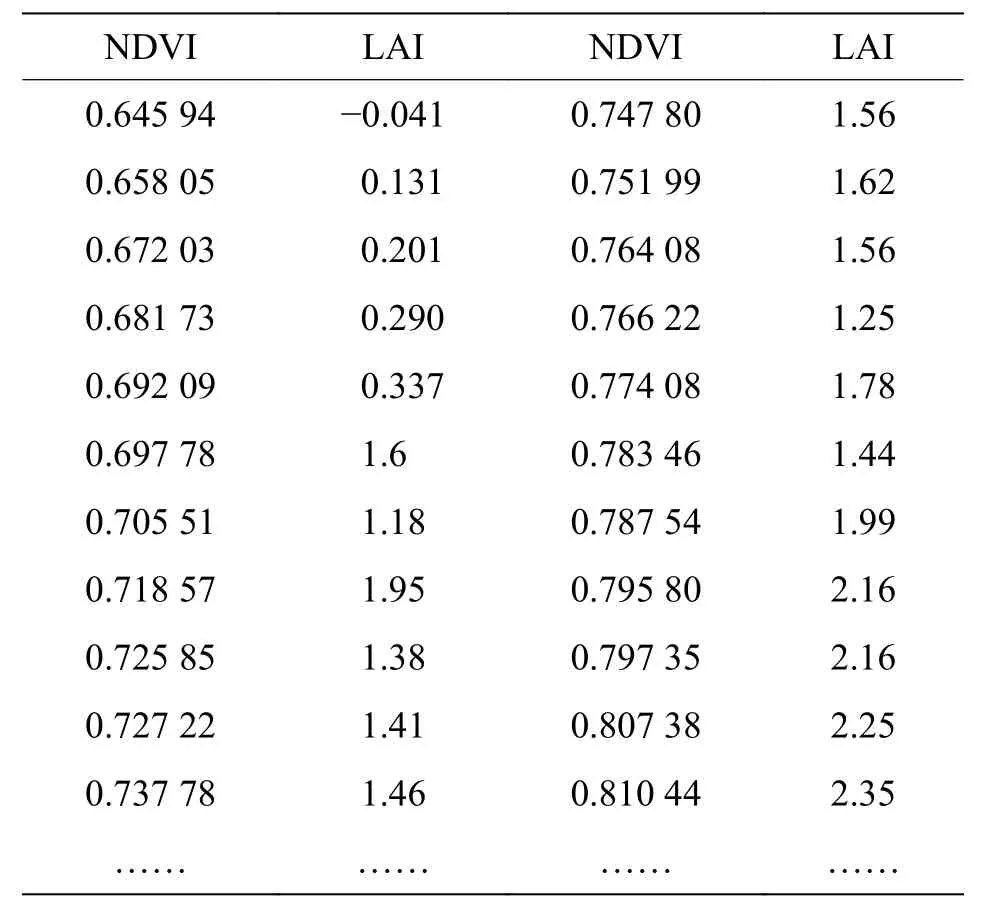
表3 部分样点归一化植被指数(NDVI)和叶面积指数(LAI)值
2.2 草地LAI 空间分布变化重度放牧下,轮牧和连续放牧的草地LAI 上升的区域占草地总面积的比例均比下降的区域占比小;中度放牧下,轮牧的草地LAI 上升的区域占草地总面积的比例比下降的区域占比大,但连续放牧的草地LAI 上升的区域占草地总面积的比例比下降的区域占比小;轻度放牧下,轮牧和连续放牧的草地LAI 上升的区域占草地总面积的比例均比下降的区域占比大。
不同放牧方式和强度下各草地的LAI 变化如图4 所示,从G1(a,b,c)和G2(a,b,c)可以看出,在轮牧方式下,重度放牧时G1 的LAI 显著上升的区域极少呈现零散分布,下降的区域占比高达94.40%,G2 的LAI 上升区域几乎为0,下降的区域占比高达99.19%;中度放牧时,G1 草地右下角处LAI 呈现显著上升状态,整体来看草地有53.37%的草地区域呈上升趋势,G2 则仅有47.91%的区域呈上升的趋势,上升区域主要集中在右上角的牛栏入口处,从平均水平上看,G1 和G2 两块草地平均上升的区域占比为50.65%;轻度放牧时,G1 各处LAI 有所上升,有高达64.21%的草地区域LAI 值呈现明显上升的趋势,G2 有57.83%的区域LAI 呈现上升趋势,上升区域分布较为均匀。从G3(a,b,c)可以看出,在连续放牧方式下,重度放牧时G3 的LAI 下降区域占比87.35%,比G1 和G2 的都要低,中度放牧时25.83%的区域LAI 呈现上升趋势,虽然下降的区域占比仍然较大,但是与重度放牧相比下降程度有所减轻,而在轻度放牧时,有60.37%的草地区域LAI 呈现了上升趋势。图中的NG(a,b,c)体现出,在没有放牧干扰的情况下,同期未放牧草地LAI 上升区域的占比始终比下降区域高,分别为67.96%、60.59%、53.83%,但可能是由于气候因素,上升区域占比逐渐减小。
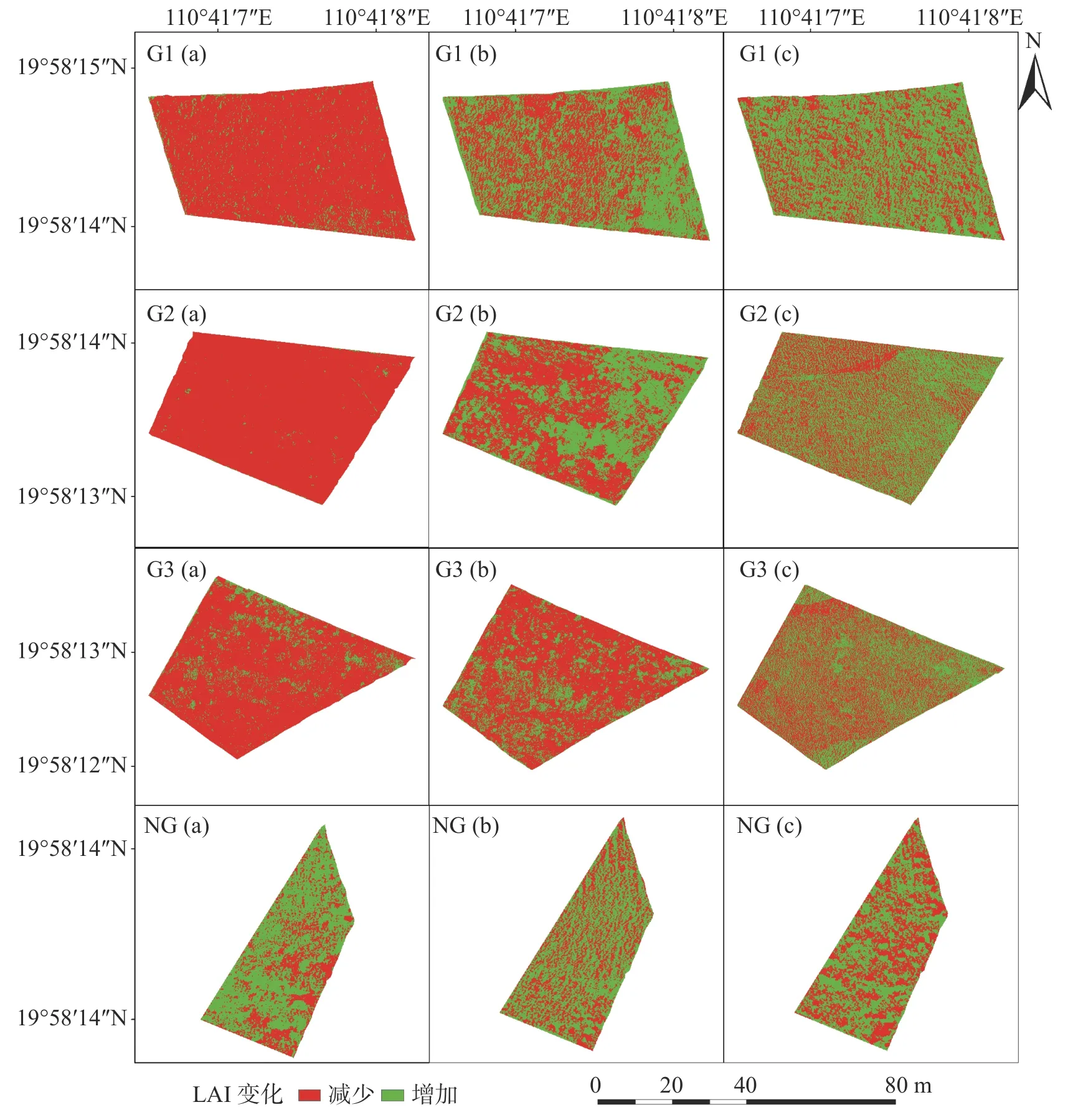
图4 不同放牧方式与强度下草地LAI 变化动态
由此可得,重度放牧下将牛集中在G1 和G2 分别进行集中的5 天放牧时,由于G1 和G2 面积较小,牛的可食用草量少,两个草地的LAI 变化降幅较大,虽然G3 的面积比G1 和G2 大,但是由于放牧头数过多和时间过长,导致3 块草地LAI 下降的区域占比都较大。中度放牧下轮牧有助于提高草地的LAI,而连续放牧由于畜牧长时间的啃食,导致即使是放牧强度降低,草地LAI 上升区域仍然较小。而轻度放牧下草地承载的畜牧量足够低时,无论是基于轮牧还是连续放牧制度,草地状况都呈现出相同的向好趋势,LAI 的上升区域占比也变得更大。
2.3 草地LAI 时间动态分析重度和中度放牧下,无论是轮牧还是连续放牧,草地LAI 均值都呈现下降的趋势,下降幅度较大,而在轻度放牧下,轮牧的草地LAI 均值呈现上升趋势,连续放牧的草地LAI 均值依旧下降,下降幅度较低。各草地的LAI 变化如图5 所示,重度放牧下G1 第一天的LAI 均值为2.01,第五天LAI 的均值为1.09,LAI 均值下降了约45.78%;中度放牧下G1 第一天的LAI 均值为2.01,而第五天的LAI 均值为1.89,相对于放牧前的LAI 均值下降了约5.97%,然而有53.37%的草地区域呈显著上升的趋势;轻度放牧下G1 第一天的LAI 均值为1.83,而第五天的平均LAI 均值仅为1.81,相对于放牧前下降了约1.09%,但是却有高达64.21%的草地区域LAI 值呈现明显上升的趋势;同期未放牧草地NG 的LAI 分别从2.00、1.99 和1.89 上升到了2.04、2.02 和1.97,上升幅度分别为1.97%、1.62%和3.66%。重度放牧下G2 第一天的LAI 均值为1.93,第五天LAI 的均值为1.12,LAI 均值下降了约41.97%;中度放牧下G2 第一天的LAI 均值为1.98,而第五天的LAI 均值为1.71,相对于放牧前的LAI 均值下降了约12.76%;轻度放牧下G2 放牧第一天的LAI 均值为1.71,而放牧第五天的LAI 均值为1.73,相对于放牧前上升了约1.17%;同期未放牧草地NG 的LAI 分别从2.04、2.05 和2.00 上升到了2.08、2.09 和2.01,上升幅度分别为2.01%、2.31%和1.01%。重度放牧下G3 第一天的LAI 均值为2.04,第十天的LAI 均值为1.03,LAI 下降了约49.51%;中度放牧下G3 第一天的LAI 均值为1.98,而第十天的LAI 均值为1.69,变化了0.24,相对于放牧前的LAI 均值下降了约14.65%;轻度放牧下G3 第一天的平均LAI 为1.71,而第十天的平均LAI 为1.70,相对于放牧前下降了约0.58%,但依旧有60.37%的草地区域LAI 呈现上升趋势;同期未放牧草地NG 的LAI 分别从2.00、1.99 和1.89 上升到了2.08、2.09和2.01,上升幅度分别为4.15%、5.12%和6.04%。

图5 不同放牧强度下草地LAI 均值变化
2.4 畜牧行为对草地LAI 的影响重度放牧下,无论是轮牧还是连续放牧都会增加牛的采食时间。根据连续记录法按每5 s 一个频次,采集牛采食、站立、慢走、躺卧和反刍的运动行为时间,图6 为采集的各个行为的比例,在G1 中,采食的时间呈现出先增加后减少再增加的趋势,G1 放牧的最后一天采食时间占比高达84.17%,较第一天增加了24.29%;躺卧的时间则呈现相反的趋势,最后一天躺卧占比仅有7.07%,较第一天减少了11.61%;站立的时间在放牧第一天和最后一天大致相同,分别为7.81%和8.57%,仅相差0.76%,但在其他时间极低;反刍的时间则在放牧最后一天达到最低,其变化趋势与采食时间完全相反,这可能是因为采食时间占用了反刍的时间;慢走时间占比始终最低,最后一天站立时间占比仅有0.19%,较第一天减少了1.62%。在G3 中,采食时间呈现出先减少后增加再减少的趋势,在放牧第四天采食时间占比达到最高值73.91%,但每日采食时间差额不大,变化较为平缓;躺卧时间虽呈现相反的趋势,但变化同样较小;站立时间在放牧后期占比较大,最高可达11.83%,较第一天增长了6.93%;反刍时间呈现出先减少后增加的趋势,在采食时间最低时反刍时间达到最高9.53%;慢走时间占比始终最低,最后一天的站立时间较第一天减少了2.51%。
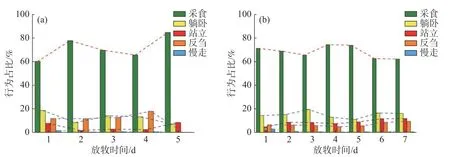
图6 重度放牧条件下牛的行为时间占比
以啃食时间占比与LAI 的比值来监测牛采食叶面积指数的量,发现在G1 和G3 中,随着放牧时间的变化,日均LAI 都越来越小,虽然G1 中啃食时间占比呈现了先增加后减少再增加的趋势,G3 中啃食时间占比则先减少后增加再减少,但是从图7 可以看出,两种放牧方式下草地的啃食时间占比与LAI 的比值都越来越大,表明牛采食同样的LAI 投入时间越来越长。综合来看,G1 平均每天采食的时间占比为71.20%,G3 为68.02%,无论是在轮牧草地还是在连续放牧草地,采食的时间占比始终最高,表明高强度放牧会显著增加畜牧采食时间。

图7 重度放牧条件下牛的啃食时间占比/LAI
当牛的采食时间增加时,LAI 减少量也随之增加,增加到一定程度就会到达草地的供给极限。图8 为牛每日采食占比与LAI 变化量的关系,LAI 变化量由每日放牧结束后的LAI 减去放牧开始前的LAI 计算获得。由图8 可知,在G1 中,当采食时间占比逐渐增加时,LAI 的变化量越大,减少量级越大,但当采食时间占比增加至70.88%左右时,LAI 变化量逐渐回弹,不再降低;在G3 中,最初的LAI 变化量也是随着采食时间占比的增大而增大,但当采食时间占比增加至73.42%左右时,LAI 变化量也逐渐呈现出上升的趋势。可见,无论是在轮牧还是连续放牧的制度下,当采食时间占比达到70.88%~73.42%左右时,已经到达了LAI 变化的极限。此时,草地叶面积指数下降到初始状态(第一天啃食前)的79.60%~79.90%,即使啃食时间占比增加,LAI 减少量也不再增加。
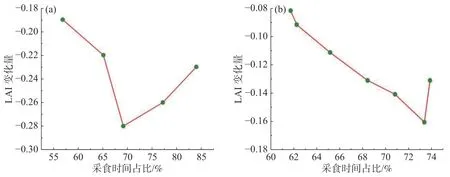
图8 牛的采食时间占比与LAI 变化量关系
3 讨 论
3.1 放牧强度对草地LAI 的影响通过放牧实验,本研究发现重度放牧对所有草地的LAI 上升不利,所有草地的LAI 上升的区域要远小于下降的区域;而在中度放牧强度下,轮牧制度的草地LAI 上升的区域占草地总面积的比例比下降的比例大,而连续放牧地块的草地LAI 上升的区域占草地总面积的比例比下降的比例小;轻度放牧强度下,轮牧和连续放牧的草地LAI 上升的区域占草地总面积的比例均比下降的比例大。过度放牧一直以来都被认为是植被叶面积指数下降的主要负驱动力[27-28]。在过去的几十年里,中国西北广大草原的环境退化程度不断加剧,过度放牧是主要原因[29]。考虑到年际气候变化也会影响到牧场草地的生长,Hao 等[30]将放牧对LAI 的影响与年际气候变化的影响分开,同样证明了放牧对植被叶面积指数的负面影响(减少LAI)极大地抵消了为草地带来的正面影响(增加LAI)。而随着放牧强度的降低,草地冠层盖度等生物指标和丰富度增加,中度放牧被证明可以显著改善植被覆盖和促进物种恢复[31-32]。Deng 等[33]在草原牧场,发现中度放牧管理可用于实现植物功能群落的理想转变,合理的放牧强度对牧场生态系统的可持续性有积极影响,通过中等强度放牧对草地进行长期有效管理既可以抵消当地的退化,也有益于草地的功能恢复。高寒草地野外观测也发现,在水热条件适宜的情况下,轻、中度放牧压力比不放牧更有利于提高叶面积指数,即轻、中度放牧区的叶面积指数高于非放牧区的叶面积指数[34]。因此,轻、中度压力的放牧对草地的贡献是积极的,有利于提高草地LAI,促进草地生长。这可能是由于草地的生长速率与草地的现存草量密切相关,重度放牧下草地现存草量少,草地再生长所需的营养物质和能量得不到充足的供应,草地再生速率降低,而在轻度放牧下,草地现存草量多,草地的营养物质维持了植被的呼吸作用,消耗掉大量能量,只有在适度放牧下,才能将植被高度和覆盖度控制在一定范围内,可以使得光线更加均匀地分布到草地内部,提高光合作用的速率和草地生长效率,牧草得到及时更新。
3.2 放牧方式对草地LAI 的影响轮牧制度的草地在重度放牧下呈现出下降趋势,但在中度和轻度放牧下草地的LAI 上升区域面积占比与下降的区域相比较大。连续放牧制度下的草地LAI 在重度和中度放牧下呈现相同的下降趋势,只有在轻度放牧时LAI 上升区域占比高于下降区域占比。当放牧强度适中时,在严格的放牧管理下如轮牧制度,可以使LAI 保持相对稳定,草地的叶面积指数出现微增。这是因为植物能够维持当前体内有机物的持续流量,通过体内的有机储备在使得草地保持在最低LAI 标准以上和不受到生长条件的限制[35],Trindaded 等[36]已发现了叶面积与消化率和营养价值呈正相关,轮牧制度通过更换放牧草地来增加放牧间隔时间,促进叶片的营养生长和草茎的伸长,保证牛的啃食量。一般来说,连续放牧的草地会导致草地管理不当,Da 等[37]发现轮牧和连续放牧之间的牧草积累和草地LAI 存在明显的差异,连续放牧所带来的高强度放牧会使草地LAI 减少,这表明在连续放牧条件下,畜牧的过度踩踏无法为植物提供可以维持生长的有机储备。Sbrissia 等[38]的结果也表明,在连续放牧的条件下,由于地上所产生的同化物会使得牧草积累,此时草地叶片的质量就对植物的生长起着至关重要的作用,甚至会阻碍植物生长,叶面积指数就会变得越低。
3.3 畜牧采食时间对草地供给的影响在重度放牧下,无论是轮牧还是连续放牧都会增加牛的采食时间,LAI 减少量也随之增加,增加到一定程度就会到达草地的供给极限。当牛的采食时间占比达到70.88%~73.42%左右时,也就是草地能够单日供给的极限后,草地可食用牧草大幅减少,牲畜个体采食竞争加剧,整体采食时间占比提高,草地叶面积快速下降。以往的研究也证明了由于牲畜对草地的直接啃食,草地地上生物量有所下降[39-40],这可能是由于畜牧在采食过程中不断踩踏,导致土壤压实从而限制了草地根系的伸长和发育[41-42]。此外也有研究表明,牲畜的踩踏和践踏会通过压实增加表土的容重[43],表土结构的这些变化导致含水量下降[44],渗透因此减少,加上植被覆盖减少将导致蒸腾减少[45]。畜牧的采食时间过长会导致植被产量和生物量减少,氨化、硝化作用和土壤肥力下降[46]。过度放牧下畜牧的高强度啃食已被认为是中国西北部草地覆盖率和质量下降、生物多样性丧失及水土流失导致土地退化的主要原因之一[47]。因此,了解草地退化背后驱动力的影响对于应对全球变化适应具有重要的生态系统管理意义[48]。为了提高草原生态系统的服务功能,应减少放牧压力,特别是在不利气候和过度放牧双重负面影响加剧的地区,轮牧应该纳入管理计划,规定在整个放牧区域的放牧时间,调整牛的啃食时间,有助于评估该地区最近大规模生态恢复活动的有效性,未来的研究也可以侧重于通过控制牛的啃食时间来了解不同放牧制度下的草地LAI 动态的生物物理过程。
4 结 论
基于多光谱无人机反演的不同放牧策略下草地叶面积指数结果显示,在中度放牧时轮牧草地LAI 上升的区域占草地总面积的比例比下降的区域大,但连续放牧草地的LAI 上升区域占草地总面积的比例比下降的区域小,这表明中度放牧条件下的轮牧有利于草地LAI 的提升。而在重度放牧下,牛的采食时间占总行为比始终最高,无论是轮牧还是连续放牧都会增加牛的采食时间,LAI 减少量也随之增加,增加到一定程度就会到达草地的供给极限,即使采食时间增加,LAI 减少量也不再增加。当然,放牧行为对草地的生长可能存在着滞后效应,这种效应在未来将会成为进行长期草畜监测研究的重要问题。
猜你喜欢
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
今日农业(2020年19期)2020-12-14
电脑报(2020年12期)2020-06-30
新疆农业科学(2020年1期)2020-02-14
农民致富之友(2018年20期)2018-11-25
中国农业科学(2017年8期)2017-05-03
安徽医科大学学报(2016年12期)2017-01-15
甘肃畜牧兽医(2016年12期)2016-09-21
癌症进展(2016年9期)2016-08-22
中国麻业科学(2015年5期)2015-12-28
