
青藏高原灌丛土壤碳氮含量及其相关功能基因分布特征
2024-01-25 06:50李传虹刘四义韩丽丽姚保民申聪聪郝志鹏张丽梅
生态学报 2024年1期
李传虹,张 林,刘四义,韩丽丽,姚保民,申聪聪,郝志鹏,张丽梅,*
1 中国科学院生态环境研究中心,北京 100085 2 中国科学院大学,北京 100049 3 中国科学院青藏高原研究所,青藏高原地球系统与资源环境国家重点实验室,北京 100101
高寒灌丛是由耐寒的中生或旱生灌木为优势种而形成的一类植被,常分布在高山和高原林线之上向高寒植被的过渡带,是青藏高原常见的优势植被之一[1]。由于独特的地理、气候条件,青藏高原的灌丛类型复杂、种质资源丰富,为野生动物和家畜提供了栖居和放牧地,具有较强的生态服务价值[2]。然而其生态系统脆弱,对全球变暖的响应极为敏感[3]。近年来,气候变暖及其导致的土壤水分、养分有效性和土壤微生物群落等环境条件变化对青藏高原地区高寒灌丛植物生产力、生物量分配和演替过程产生了深刻影响,进而影响着区域生态系统碳氮循环和周转过程[4]。因此,探明青藏高原不同灌丛类型下土壤养分状况、碳氮循环特征及其影响因素对青藏高原灌丛合理利用与管理有重要的理论与实践价值。
微生物是土壤物质转化和元素生物地球化学循环的主要驱动力[5]。微生物驱动的矿化作用,即土壤中有机态化合物转化为无机态化合物的过程,是自然生态系统土壤碳氮循环的关键过程之一[6]。土壤碳氮矿化过程深刻影响着土壤碳、氮等元素的循环流动,土壤的养分供应、有机质的更新和地上生产力[7-8]。碳和氮矿化速率的微小改变能够显著影响土壤碳库和氮库大小以及大气二氧化碳浓度,最终影响碳氮循环-气候耦合系统的动态变化[9],对地上植被的演替和分布有极为重要的意义。大量研究表明,气候因子如温度和降水是影响土壤碳氮矿化速率的主要因素,温度升高和降雨量增加可以提高碳氮矿化速率从而提高土壤中养分的有效性和净生产力[10-11]。除气候因素外,土壤有机质含量、pH、水分等对碳氮矿化速率也有交互影响。有机质含量越高,矿化速率越高;同时,随土壤pH升高,土壤有机质的可溶性增加,为微生物生长提供了充足的生长基质,因此碳氮矿化速率通常与土壤pH呈显著正相关关系[12-14]。此外,土壤水分一方面可直接控制微生物的活性,另一方面可通过影响凋落物的分解和有机质的形成间接地影响碳氮矿化速率[15]。研究表明土壤氮矿化速率通常随土壤水分增加而增加,在土壤含水率达到田间持水量时最高,并随含水率进一步增加而降低[16],如亚热带林区氮矿化速率与土壤含水量呈显著负相关[17]。碳和氮的矿化速率也可能受土壤pH和含水量等因素的交互影响,如与湿润地区相比,干旱地区土壤pH相对较高,矿化能力也相对较强[18]。青藏高原灌丛面积分布广泛,气候和土壤环境复杂[19],但目前对灌丛分布地区土壤碳氮矿化作用强度、影响因素及其与灌丛分布特征的关系等的认识较少。探明青藏高原灌丛分布区土壤碳氮含量、矿化作用特征及其影响因素,对理解青藏高原气候变化背景下,灌丛分布区土壤质量变化及其对地上植被的影响具有重要意义。
此外,土壤碳、氮元素矿化过程复杂,参与反应的微生物类群众多,并与其他过程耦合发生,可能导致土壤CO2排放增加、温室气体产生和养分流失等问题[20-21]。但目前对碳氮循环相关功能微生物的研究多针对单一元素或单一过程开展,缺乏大尺度下自然生态系统土壤碳、氮循环关键过程功能基因分布特征的耦联分析,且在不同研究尺度和生态系统类型中开展的研究结果不同[22-23]。对参与土壤碳、氮循环过程相关的功能基因多样性和丰度的调查有助于更好地理解微生物介导的元素生物地球化学过程及其对全球变化的响应和反馈[24-26]。因此,本研究采用高通量定量PCR(quantitative microbial element cycling,QMEC)技术,对青藏高原喜马拉雅山-冈底斯山地区沿降水梯度采集的高寒灌丛土壤样品碳氮循环相关的功能基因多样性进行了分析,并结合土壤理化性质、气候、植被等信息,综合探讨了青藏高原不同灌丛分布区域土壤养分状况和参与碳氮矿化过程相关功能基因的分布特征、驱动因子及其与灌丛分布特征的关系,以期为认识气候变化背景下青藏高原土壤质量和灌丛生态系统功能变化趋势提供重要信息和依据。
1 材料与方法
1.1 研究区域概况与土壤样本的采集
土壤样品采集于2020年8月第二次青藏高原科学考察喜马拉雅山-冈底斯山沿线灌丛样带调查,沿线经革吉-吉隆-昆莎机场-拉昂错-嘛咪乡-门士乡-马攸木拉山-普兰-帕羊-狮泉河-扎达县-如角-珠峰-茶巴拉乡-多角乡-贡嘎县-南木乡-日喀则-扎囊县-拉孜-八宿-东达山-拉藏-南木林-折巴-拉乌山-业拉山(79.97°-98.68°E,28.31°-32.62°N)等地区,最远相距1800 km。植被以灌丛为主,包括5种优势灌木类型:变色锦鸡儿(CV)、金露梅(DF)、砂生槐(SM)、香柏(JP)、雪层杜鹃(RN)。根据同一灌丛的分布面积和间隔距离,共布设了28个采样点,每个采样点间隔至少9 km。使用手持式全球定位系统(GPS)记录采样点的经度、纬度等地理数据,并从中国科学院资源环境科学数据中心(http://www.resdc.cn/)提取年平均温度(MAT)、年平均降水量(MAP)和植被NDVI指数[27],采样点分布和对应灌木类型如图1所示。

图1 采样点的地理位置分布示意Fig.1 Geographic locations of sampling sites
每个采样点尽量设置在远离人类居住的地区,以本地分布的天然灌丛为主。每个采样点随机选取3个5 m×5 m样方,每个样方按梅花状进行5点取样混合成一个样品,取样深度0-15 cm,最终共采集到84份土壤样本。所有样品在去除根系和大的石块后放入无菌塑料袋中,通过便携式车载移动冰箱运到实验室。土壤样品通过2.0 mm筛去除细根和砾石后分成两部分:一部分储存在4 ℃下,用于土壤理化性质分析,另一部分储存在-80 ℃下,用于土壤总DNA提取。
1.2 土壤理化性质测定
土壤理化性质测定参考文献[28]:土壤pH(水土比2.5∶1)采用酸度计电位法测定,土壤含水量采取烘干称重法测定,土壤速效磷(Available Phosphorus,AP)采用Olsen法测定,土壤全碳(Total Carbon,TC)和全氮(Total Nitrogen,TN)含量采用元素分析仪(Vario EL III-Elementar,Germany)测定,土壤无机碳(Inorganic carbon,IC)含量采用总有机碳分析仪(SSM-5000A,Japan)测定,土壤有机碳(Soil organic carbon,SOC)为TC与IC的差值,碳氮比(C:N)为SOC与TN的比值。
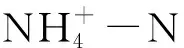
土壤CO2释放速率(SRR)的测定:称取10 g土壤样品放入培养瓶中,用保鲜膜封好瓶口后放置于20 ℃培养箱预培养24 h。培养完成后将培养瓶放入封闭动态系统中,使用LI-820 CO2分析仪进行CO2浓度实时动态测定,得到封闭动态系统中CO2浓度的增加速率a,利用线性回归方法计算土壤的CO2释放速率[31]。
1.3 土壤DNA提取与QMEC检测
称取0.5 g土壤样品,利用Dneasy PowerSoil Kit(QIAGEN GmbH,Germany),按步骤对土壤微生物总DNA进行提取。使用NanoDrop分光光度计(NanoDrop Technologies Inc.,Wilmington,DE,USA)测定DNA的浓度和纯度。DNA样品存储在-40 ℃冰箱中备用。
QMEC包含71个微生物CNPS引物和1个细菌类群引物,可以平行定量72个DNA样本,引物与基因参照文献所示[32]。主要操作过程如下,首先将检测合格后的DNA样品添加至384孔板作为样品板(Sample Sourceplate),同时将引物和qPCR所用的试剂添加至另一384孔板作为引物板(Assay Sourceplate)。采用高通量自动微量加设备SmartChip Multisample Nanodispenser(Takara Biomedical Technology)分别将样品板和引物板试剂添加至高通量qPCR芯片SmartChip MyDesign Chip(Takara Biomedical Technology,Clontech)的微孔中,在SmartChip Real-Time PCR System(WaferGen Biosystems USA)中进行qPCR反应及荧光信号检测,并自动生成扩增曲线和溶解曲线[5]。根据SmartChip Real-Time PCR System和Canco software软件给出的各基因在各样本中的Ct值,根据公式(1)计算得出各基因在各样本中的相对丰度信息。只有在三个技术重复均被检出的基因,才会将该基因判定为阳性,并计算其平均值作为该基因在对应样本中的相对定量。以F525/R907为引物通过普通定量PCR(Roche,LightCycler480Ⅱ)获得细菌16S rRNA基因的绝对丰度信息,16S rRNA相对丰度默认为1,根据公式(2)计算得到各样品中各基因的绝对定量信息[32]。
基因相对丰度= (31-Ct)/(10/3)
(1)
基因绝对丰度=基因相对丰度×16S rRNA绝对丰度/16S rRNA相对丰度
(2)
本文从71 个功能基因数据中选取了 18个碳降解功能基因和7个氮矿化功能基因做进一步深入分析。参考 Benbi和 Richter 等[33]学者的观点,本文将硝化作用并入氮矿化过程,因此将 AOA(氨氧化古菌)amoA、AOB(氨氧化细菌)amoA、amoB、hao、nxrA基因也纳入矿化基因进行分析。
1.4 数据分析
利用SPSS 26.0软件,基于Kruskal-Wallis检验对不同灌丛类型间碳、氮矿化基因的丰度和土壤理化性质进行非参数检验,以及分别将碳、氮基因丰度进行因子分析降维并标准化转化。使用Pearson相关性分析研究碳、氮矿化基因丰度之间以及与土壤基本理化性质及环境因子之间的相关性。此外,用R(v.4.2.1)软件的“vegan”进行Mantel test分析,以探明青藏高原喜马拉雅山-冈底斯山沿线高寒灌丛区碳、氮矿化过程相关微生物的空间分布格局的主要驱动因子。
2 结果与分析
2.1 不同灌丛类型土壤养分状况及其影响因子


表1 采样点基本信息及土壤理化性质Table 1 Geographical information of sampling sites and soil physiochemical properties

图2 土壤理化性质与气候和植被因子间的相关性分析Fig.2 The relationship between soil properties and climatic and vegetational factors圆圈的颜色表示相关性的大小;MAT:年平均温度 Mean annual temperature;MAP:年平均降雨量 Mean annual precipitation;TN:全氮 Total N;SOC:土壤有机碳 Soil organic C;C:N:碳氮比 SOC:TN;Mnet:氮净矿化速率 Net N mineralization rate;SRR:CO2释放速率 Soil respiration rate
2.2 碳、氮矿化过程功能基因的丰度分布特征
基于QMEC检测,我们对与碳矿化和分解相关的18个基因,以及与氮矿化作用相关的7个基因进行了统计分析。结果表明,除丰度较低的淀粉水解酶基因amyX、木质素降解酶基因pox和纤维素降解酶基因naglu,其余22个基因的丰度在五种灌丛类型差异显著(P<0.05,图3)。丰度总体表现为从青藏高原西部向东部降低的趋势,变色锦鸡儿、金露梅和砂生槐灌丛土壤基因丰度普遍高于香柏和雪层杜鹃灌丛,并以雪层杜鹃灌丛的丰度最低(图3)。但细菌16S rRNA基因的绝对丰度(4.5×109-5.1×109拷贝数/g干土)在不同灌丛类型土壤中无显著差异(P>0.05,图3)。

图3 青藏高原不同灌丛类型土壤碳氮矿化功基因丰度热图Fig.3 Heatmap of absolute abundances of C and N mineralization genes in soils separated according to shrub types on the Tibetan Plateau不同灌丛类型间同一基因的不同字母表示差异显著(P<0.05)
易降解碳矿化基因中,半纤维素降解酶基因abfA丰度最高(2.9×106-5.3×107拷贝数/g),其次为半纤维素降解酶基因manA(2.4×106-3.2×107拷贝数/g)和xylA(1.2×106-3.3×107拷贝数/g),淀粉水解基因amyX丰度在所有碳氮矿化基因中最低(0-6.8×103拷贝数/g)(图3)。难降解碳矿化基因中,木质素降解酶基因mnp丰度最高(6.6×105-1.1×107拷贝数/g),纤维素降解酶基因naglu丰度最低(0-3.1×104拷贝数/g)(图3)。尿素酶基因ureC的丰度在所有碳氮矿化基因中最高,为5.8×107-3.0×108拷贝数/g,谷氨酸脱氢酶基因gdh丰度也较高,为2.7×106-2.6×107拷贝数/g(图3)。硝化过程相关基因中,古菌的氨单加氧酶基因amoA和amoB丰度最高,分别为8.1×104-7.6×106拷贝数/g和4.5×105-8.6×106拷贝数/g,羟胺氧化还原酶基因hao丰度最低,为0-1.3×105拷贝数/g(图3)。
2.3 影响碳氮矿化基因丰度的关键因素
Mantel test分析结果显示土壤SOC、TN、pH、MAP、SWC对土壤易降解碳矿化基因、难降解碳矿化基因、氨氧化基因和硝化基因的丰度均有显著影响(P<0.05),且以SOC、TN、pH和土壤水分的解释量(R值)最高(表2)。相关性分析结果显示,C、N矿化基因丰度与MAP、土壤含水率、SOC和TN含量均为显著负相关(P<0.001),但与土壤pH均为显著正相关(P<0.05),与年平均温度(MAT)无显著相关性(P>0.05)(图4)。此外,除丰度较低的amyX基因外,其余C、N矿化相关的功能基因丰度之间均存在显著的正相关性(P<0.05)(图5)。
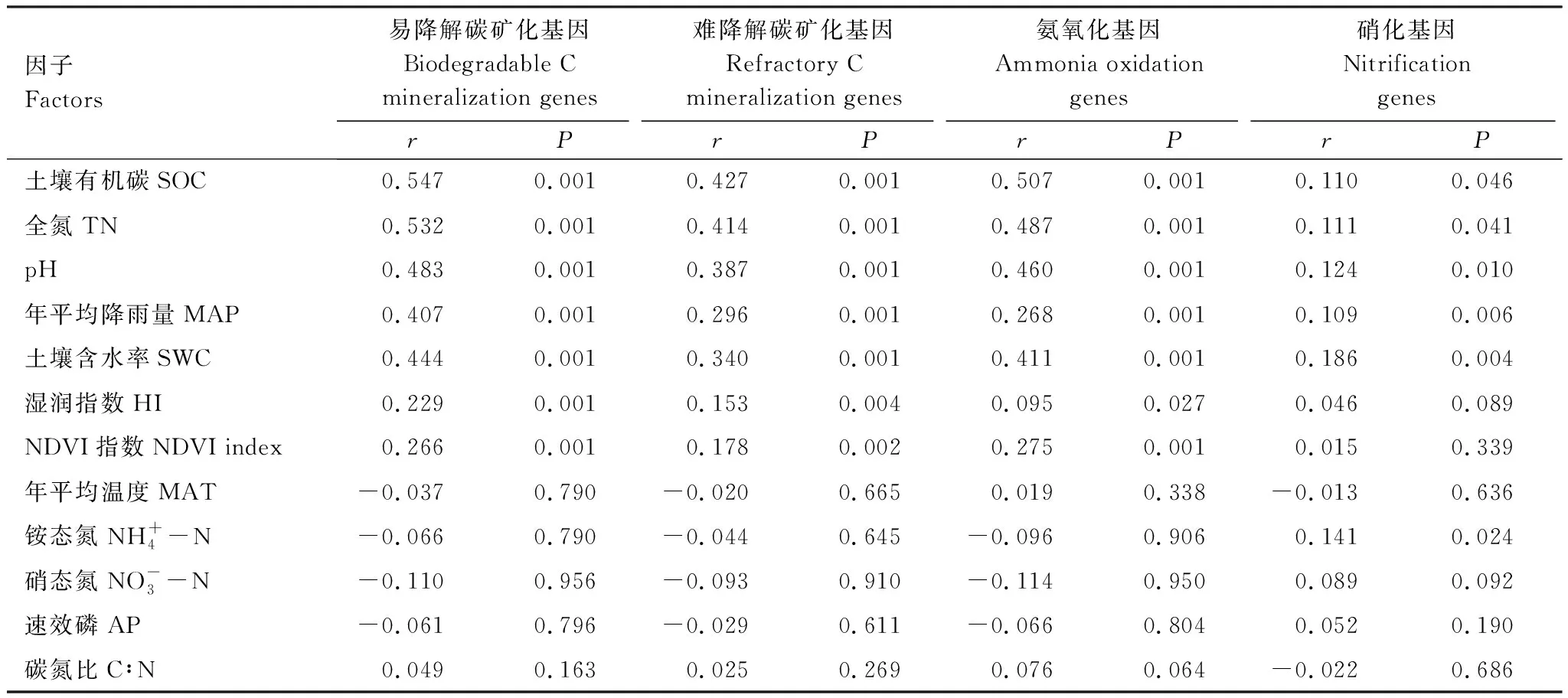
表2 碳和氮矿化基因丰度与环境因子的Mantel test分析Table 2 Mantel test analysis for the correlation between the abundance of C and N mineralization genes and environmental factors
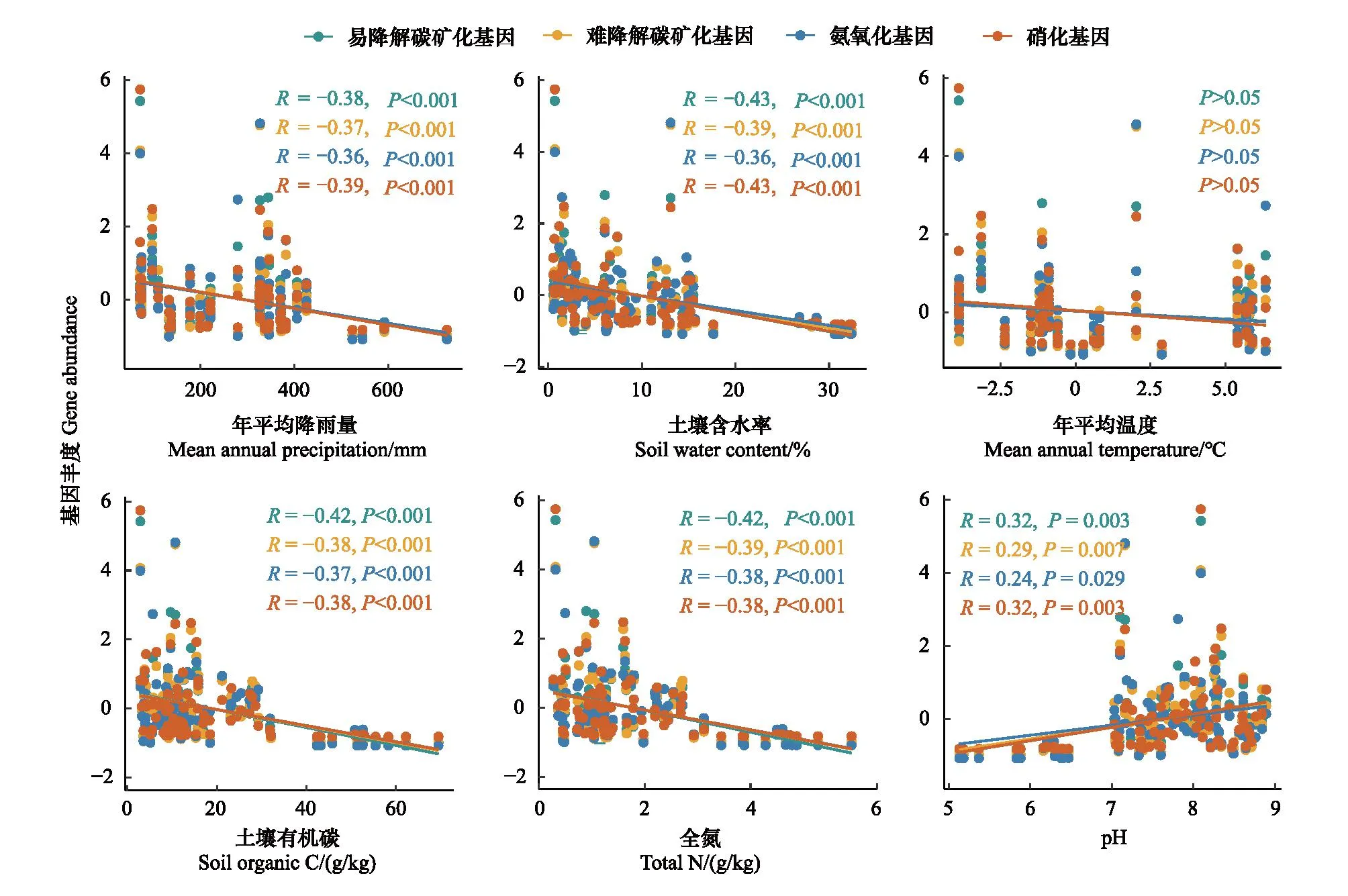
图4 环境因子与碳、氮矿化基因丰度间的相关性分析Fig.4 The correlation between environmental factors and the abundance of the C and N mineralization genes矿化基因丰度值经过了标准化转换

图5 土壤碳和氮矿化相关功能基因丰度间的耦合关系Fig.5 The coupling relationship among the abundance of C and N mineralization functional genes圆圈的颜色和大小表示相关性的大小
3 讨论
土壤中碳氮元素的含量及其周转对地上植被的生长发育和生态功能的发挥极为重要。不少研究表明灌丛、森林、草地等植被类型演替过程中土壤的性质也发生了显著变化[34]。如Hibbard等[35]发现由于灌木叶片中木质素和纤维素含量低于草本植物,亚热带草原灌丛化过程使0-10 cm土壤有机质和全氮含量显著提高8-23 g/m2,氮矿化速率增加3-5倍。本文研究的结果显示,不同类型灌丛土壤的有机碳和全氮含量、土壤CO2释放速率、净氮矿化速率和碳氮矿化基因的丰度有显著差异。其中,位于东南部的雪层杜鹃和香柏灌丛比位于中西部的变色锦鸡儿、金露梅和砂生槐灌丛土壤具有更高的有机碳、全氮含量和CO2释放速率、净氮矿化速率,但碳、氮矿化基因丰度更低。这意味着灌丛类型的演替也同样伴随着土壤元素循环、养分供应的改变。由此可见,相同植被类型下,灌丛种类演替对青藏高原生态系统功能的影响也值得关注。
温度和降水是影响土壤元素循环和养分供应的重要影响因素。本研究结果显示,土壤有机碳、全氮含量、CO2释放速率、净氮矿化速率与年平均降雨量、湿润指数、土壤含水量等水分指标显著正相关,而与年平均温度无显著相关性。这些结果说明水分条件的变化对土壤碳氮等元素含量和矿化过程有更重要的影响。已有研究表明,降水减少会通过降低微生物生物量和胞外酶活性来抑制凋落物的分解[36-39]。与本文的研究结果类似,苟小林等[40]也认为由于更充沛的水分条件,青藏高原半湿润地区的植物生物量和土壤碳氮磷等元素含量比半干旱地区显著更高。喜马拉雅山-冈底斯山自西向东降雨量明显增加,降水量增加一方面可直接影响土壤理化性质和微生物活性促进土壤养分的周转,另一方面可通过影响植物生长间接提高土壤养分含量[41]。这些结果表明,降水条件改变对青藏高原灌丛土壤碳氮循环的影响更大。目前青藏高原正面临着暖湿化的影响,降水增加引起的碳氮循环变化及其带来的生态环境效应值得进一步关注。
由于微生物的活动需要水分,所以在一定湿度范围内,土壤水分增加有利于矿化作用发生[10,42]。然而,本文研究却显示土壤碳氮矿化基因丰度与年平均降水量呈显著的负相关关系,但与土壤pH呈显著的正相关。导致这一现象的原因主要有两方面:一方面,pH可直接影响微生物的生理代谢,过低或过高的pH会对微生物产生生理限制,并影响微生物间相互作用进而影响其群落组成和丰度[43];另一方面,土壤pH作为一个综合变量,与一些土壤养分可利用性、有机碳特性和土壤水分状况等密切相关,土壤pH的升高会增加土壤有机质的可溶性,为微生物生长提供了基质从而间接影响了微生物的多样性和丰度[12-13]。大量研究表明,土壤pH是影响土壤细菌群落空间分布格局及多样性的主要驱动因素[44]。与我们的研究结果一致,Tsiknia等[45]发现在流域尺度土壤pH与氮循环功能基因丰度显著正相关。对我国大尺度区域不同生态系统类型土壤的研究发现,pH是影响氨氧化微生物分布和功能活性的主要因子,氨氧化细菌和古菌amoA基因的丰度与土壤pH值呈显著正相关[46-47]。乔沙沙[48]研究发现土壤pH通过影响土壤微生物群落结构,决定碳代谢相关微生物的丰度,进而通过影响碳代谢功能基因的丰度来影响土壤碳代谢过程。然而,我们的研究发现土壤细菌的丰度在不同植被类型下无显著差异,与土壤性质、MAP、植被NDVI指数等均无显著相关,这些结果表明功能基因对pH的响应比细菌更敏感。细菌的多样性远远高于参与C、N转化的功能微生物,因而具有更多生理代谢类型和更宽的pH适应范围。特别值得注意的是,本文结果显示土壤的pH与年平均降雨量、湿润指数和土壤含水率均为显著负相关,这说明气候因素,特别是降水改变对土壤pH有重要影响。如有研究发现水分平衡的变化会导致碱性土壤向酸性土壤逐渐过渡,这主要是由于当降雨量大于蒸发量时,土壤及其母质的淋溶作用十分强烈,土壤溶液的盐基离子易于随渗滤水向下移动,导致土壤中易溶性成分减少,溶液中H+取代土壤胶体表面交换态的金属离子被土壤所吸附,这些被取代的金属离子和酸根阴离子一起淋失,使土壤盐基饱和度下降、氢饱和度增加,引起土壤酸化[49]。因此,未来需要重视青藏高原降水改变引起土壤pH变化影响碳氮循环功能微生物代谢,进而对土壤碳、氮等元素的赋存及其周转过程产生影响。
除此之外,土壤养分也是影响功能基因分布的关键因子,大多研究表明基因丰度与土壤养分呈正相关关系[23,50-52]。如斯贵才等[53]研究发现,随着年平均降水量的下降,土壤有机碳和全氮含量显著下降,细菌、真菌和放线菌含量随着年降水量的下降而显著减少。然而,我们的研究发现与之相反,碳、氮矿化过程相关的25个功能基因的丰度均与土壤SOC和TN含量呈负相关,与无机氮含量不相关,这也表明降水条件所导致的土壤pH的差异对土壤碳氮循环功能基因丰度的影响可能远远超过了养分的影响。本研究中位于中部的砂生槐灌丛所处海拔最低,年均温和pH在所有灌丛类型中最高,降雨量适中,土壤有机碳和全氮含量最低,不及其他灌丛类型1/2,但其功能基因丰度比碳氮含量最高的雪层杜鹃灌丛高。这些结果表明较高的功能基因丰度以及适宜的温度、湿度和pH条件,可能加速了土壤中有机碳氮养分库的分解,不利于碳氮养分的累积。综合以上,这些结果表明青藏高原土壤碳氮养分库和矿化作用受土壤pH、气候、植被和功能微生物多因素的共同调控,这些结果为准确评估青藏高原土壤碳、氮库及其动态平衡提供了重要参考。
本研究中,土壤CO2释放速率、净氮矿化速率与碳氮矿化基因丰度之间没有显著的正相关关系。类似地,不少研究发现在DNA水平上功能基因丰度与碳氮循环转化速率无显著相关。如汪小兰等[54]发现紫色水稻土的硝化作用活性与AOA和AOB的基因拷贝数无显著相关性。肖海兵[55]研究发现侵蚀区土壤细菌丰度显著高于沉积区,但有机碳矿化速率显著低于沉积区;刘琪等[56]也发现微生物生物量较低的土壤在培养初期(前20 天)CO2排放速率是高微生物生物量土壤的1.1-6.1倍。此外,土壤中不同碳氮转化过程耦合发生,矿化和硝化作用的产物可作为其他反应过程的底物被消耗,如土壤有机氮矿化所释放的无机氮,也会进一步被其他土壤微生物同化利用,导致净矿化速率测定值低于实际的氮矿化速率。此外,在碱性土壤中有机碳分解矿化所释放的CO2还可以被碱性物质吸收,以碳酸盐的形式保存在土壤中,从而降低碱性土壤的CO2排放速率。因此,未来研究可利用碳氮稳定性同位素示踪技术进一步区分土壤碳氮转化具体过程(矿化、同化等)的转化速率,为高原土壤的碳氮管理提供理论指导。
土壤中碳、氮循环存在强烈的耦合关系,土壤微生物驱动的碳氮耦联作用机制也是土壤微生物生态学研究的热点之一[57]。本研究发现参与碳、氮矿化的基因类群之间存在显著的正相关关系,表明碳氮循环过程互相依赖,紧密联系。土壤中碳氮的周转受其计量比的控制[58],在氮素充足的条件下,土壤中较高的植物残体投入量及其较低的碳氮比导致微生物更倾向于选择“新鲜”碳源底物,进而减少了原有有机碳的矿化[59];反之,在氮素受限条件下,高碳氮比的植物残体可能会增加微生物对无机氮素的需求,刺激土壤原有有机质的分解[60]。本研究发现土壤C:N与CO2释放、净矿化速率呈显著正相关,这也进一步证明了上述推断。同样,土壤微生物自身也维持相对稳定的CNP比值,据估计,全球土壤微生物的CNP比值的平均值为60∶7∶1[61]。显然,青藏高原碳氮元素循环紧密耦合,调控和驱动生物地球化学过程,加剧了其过程的复杂性和不确定性[57]。
土壤中大多数微生物都参与土壤关键元素的生物地球化学过程,但这些微生物种类繁多,数量巨大,哪些微生物优先参与、如何参与仍不清楚[62]。本研究利用高通量定量PCR技术,对青藏高原不同灌丛类型下参与土壤C、N循环的多个功能基因丰度进行了表征,一定程度上也代表了参与C、N循环的功能微生物的多样性及其潜在功能活性,未来还要结合宏基因组学、宏转录组学和地球化学分析技术等对土壤碳、氮元素的耦合作用过程和调控机理开展深入研究。
4 结论
青藏高原喜马拉雅山-冈底斯山地区不同灌丛类型土壤的碳氮含量、碳氮矿化速率及其相关功能基因的丰度有显著差异,位于东南部的雪层杜鹃和香柏灌丛区比位于中西部的变色锦鸡儿、金露梅和砂生槐灌丛具有更高的土壤有机碳含量、全氮含量、CO2释放及净氮矿化速率,但碳、氮矿化基因丰度更低。土壤有机碳、全氮含量、CO2释放速率、净氮矿化速率与年平均降雨量显著正相关,表明降水条件的改变对土壤碳氮元素赋存及其矿化过程有重要的影响。土壤碳氮矿化基因丰度与土壤pH呈显著的正相关,同时pH与年平均降雨量、湿润指数和土壤含水率均为显著负相关,这意味着青藏高原暖湿化背景下,降水条件改变可能会通过影响土壤pH来调节碳氮循环相关功能微生物的丰度和活性,从而对灌丛地区的土壤养分和灌丛的分布产生重要影响。同时,碳、氮矿化相关功能基因丰度在各类群间呈现显著的正相关关系,进一步证实了土壤碳氮循环过程间紧密的耦合关系,这些结果为准确评估青藏高原土壤碳、氮库及其动态平衡提供了重要参考。
猜你喜欢
水土保持学报(2022年5期)2022-10-10
北京大学学报(自然科学版)(2022年4期)2022-08-18
西北农林科技大学学报(自然科学版)(2022年3期)2022-04-01
河北医学(2021年10期)2021-10-27
草业学报(2021年9期)2021-09-22
中国临床医学影像杂志(2019年6期)2019-08-27
草业科学(2019年5期)2019-06-04
农业环境科学学报(2017年2期)2017-03-20
天津师范大学学报(自然科学版)(2016年4期)2016-12-14
发明与创新(2015年25期)2015-02-27
