
老年性聋的研究和防治任重道远*
2024-01-29 08:30杨军陈建勇侯书乐陈鹏辉段茂利
听力学及言语疾病杂志 2024年1期
杨军 陈建勇# 侯书乐 陈鹏辉 段茂利
老年性聋又称年龄相关性听力损失(age-related hearing loss,ARHL),是因衰老的累积效应引起听觉系统的老化和退变所导致的听力下降。ARHL是老年人群中最常见的一种慢性感觉缺陷,是继心脏病和关节炎之后影响老年人的第三大常见健康问题。ARHL主要的听力学特征表现为双耳渐进性、对称性感音神经性听力损失;以轻度至中度的中-高频(2~8 kHz)听力下降为主;言语分辨能力下降,尤其是在嘈杂环境中言语感知和理解困难。因此,常导致沟通困难、社会孤立、认知能力下降,增加阿尔茨海默病的患病风险,明显降低老年人群的生活质量[1]。此外,ARHL对老年人群的精神、心理、生理也会造成一系列不利影响,增加社会负担。本文就近年来ARHL的相关基础与临床研究进行梳理和总结,旨在为ARHL的防治提供理论依据。
1 ARHL相关基础研究
毛细胞和螺旋神经节细胞的不可逆损伤以及血管纹的变性是ARHL的主要病理学改变。目前ARHL病理机制的研究主要围绕氧化应激、炎症反应、细胞凋亡和自噬细胞保护这四个方面[2]。研究认为随着年龄的增长抗氧化活性降低,进而加剧活性氧(reactive oxygen species, ROS)的产生[3]。过量的ROS引起线粒体DNA基因位点的缺失和/或突变,导致氧化磷酸化的失调,继而诱导能够启动凋亡途径的细胞损伤过程[4]。目前研究结果表明,NRF2-KEAP1(Kelch样ECH相关蛋白1-NF-E2相关因子2)信号通路参与了这一氧化应激过程。核因子-红细胞2相关因子2(NRF2)会随着年龄的增长而下调,导致NRF2调节的细胞保护和氧化还原稳定性受到阻碍,包括调节线粒体稳态、调节抗氧化酶表达、防止细胞凋亡、DNA修复和转录调节[5]。与年龄相关的NRF2活性降低还会影响许多其他生化过程,例如,在持续的氧化应激过程中,NRF2会降低5′腺苷单磷酸酯活化蛋白激酶(AMPK)的表达,进而抑制延长的自噬反应,从而防止细胞内容物随着时间的推移而过度破坏[6]。
上述氧化应激、细胞凋亡激活和线粒体功能障碍的免疫系统失调会产生一系列的慢性炎症反应。慢性炎症和氧化应激共同作用产生的ROS通过核因子κB(NF-B)的ROS激活而联系在一起,长期的氧化应激会引发慢性炎症和促炎细胞因子的产生,从而增加ROS的产生;另外在促炎细胞因子的促进下,进一步增加炎症反应,导致ROS的产生增加,进而启动导致慢性炎症损伤的正反馈效应[7]。
细胞凋亡已被证明是通过毛细胞死亡导致ARHL的主要因素。动物模型研究发现ARHL的发展主要与线粒体DNA缺失和重复、NOX过表达和解偶联蛋白2(UCP2)有关。与对照组相比,ARHL模型鼠的血管纹、螺旋神经节和Corti器中NOX3的表达以及切割的胱天蛋白酶3(CASP3)显著增加[8]。ARHL模型鼠的研究表明,腹侧耳蜗核中NOX2的表达和内耳结构中NOX3的表达显著增加。此外,研究还发现ROS作为p53上调因子,诱导持续的Bak激活,最终导致细胞凋亡(Someya等,2009)。类似地,在C57BL/6J ARHL小鼠模型中,Bak基因缺失阻止了毛细胞和螺旋神经节神经元细胞的凋亡(Someya等,2010)。
蛋白质合成发生在内质网(ER)内,ER评估合成的蛋白质是否存在异常组装,随后通过自噬溶酶体或泛醌蛋白酶系统进行破坏和回收,这个过程称为未折叠蛋白反应(UPR),是一种应激反应。慢性UPR激活和ER应激最终会抑制自噬,增加错误折叠和受损蛋白质的程度,并可能通过NF-B激活触发ROS的产生[9]。此外,FOXG1通过调节自噬在听觉退化过程中也发挥重要作用。FOXG1的抑制降低了自噬活性,并导致ROS的积累和随后耳蜗毛细胞的凋亡。通过阿司匹林给药增加FOXG1的表达,可进一步激活自噬,减少ROS的产生,进而抑制细胞凋亡,促进模拟衰老的HC和HC样OC-1细胞的存活[10]。
2 ARHL相关的基因遗传学研究
ARHL相关易感基因可能参与编码结构蛋白和稳态蛋白,这些蛋白通常是维持听觉细胞结构和功能到老年所需的。这些基因的亚型等位基因会使得对ARHL相关环境或生活方式等风险因素的抵抗力降低。另外活性氧物质代谢、抗氧化系统和线粒体功能相关的基因已被报道为ARHL易感性基因,但耳蜗感觉上皮中mTORC1基因缺失可以保护小鼠免受ARHL的侵害[11],耳蜗毛细胞中mTORC1活性的降低可能是预防ARHL的潜在策略。
尽管小鼠是研究哺乳动物ARHL发病机制的主要模式动物,但只有约四分之一的拟议人类ARHL基因与拟议小鼠ARHL基因存在重叠(Bowl,2015),仍需要一种更综合的方法来深入了解ARHL遗传学机制,如图1[12]所示。
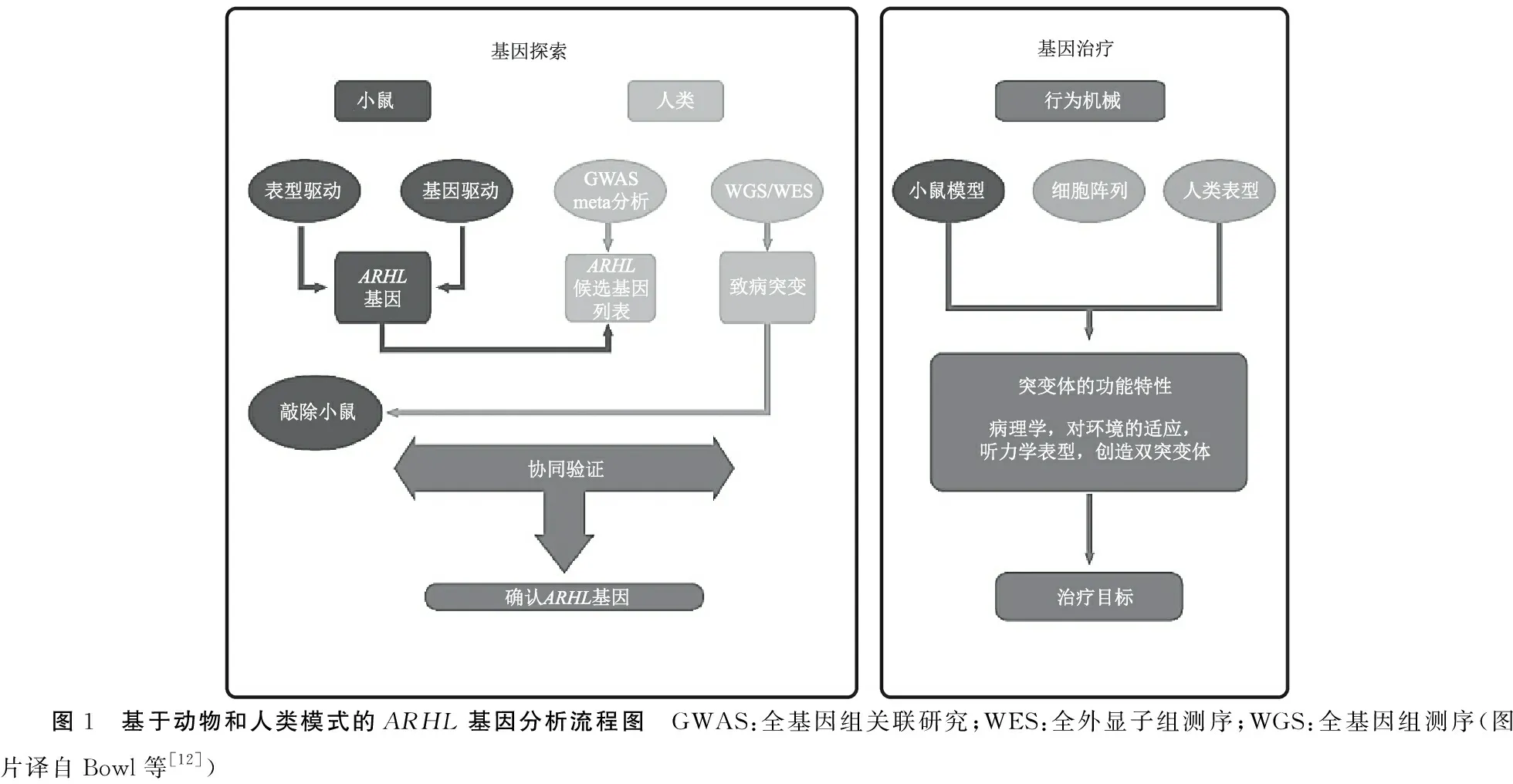
图1 基于动物和人类模式的ARHL基因分析流程图 GWAS:全基因组关联研究;WES:全外显子组测序;WGS:全基因组测序(图片译自Bowl等[12])
另外,在人类基因遗传学方面,ARHL的全基因组关联研究(genome-wide association study,GWAS)显示许多候选基因与ARHL有关,其中与ARHL密切相关的候选基因包括GRM7、CDH13、IQGAP2、DCLK1、PTPRD、GRM8、CMIP、ESRRG、ACVR1B、CCBE1、SIK3、PCDH20、SLC28A3、ISG20、TRIOBP、ILDR1、EYA4[13]。在这些候选基因中关联性最强的是GRM7基因。GRM7是一种编码谷氨酸代谢型受体7的基因,主要在毛细胞和螺旋神经节神经元中表达,当GRM7中的一种遗传变异发生时,会导致内毛细胞和听觉神经元之间突触连接处的谷氨酸释放增加,并逐渐累积到毒性水平,进而促进ARHL易感性。
另外与ARHL相关的一些少见基因,近年来也逐步受到关注和报道,包括已知基因KLHDC7B、MYO6、SYNJ2、TECTA、PUS7L、EYA4、ARHGEF28和新型基因PDCD6、PIK3R3、BAIAP2L2、CRIP3、KLHDC7B、MAST2、SLC22A7,单细胞测序显示这些基因在内耳组织细胞中广泛表达[14]。但目前这些罕见基因的发现都只是在欧洲白人的独立样本中获得复制,仍有必要进行更多的研究来阐明这些变体/基因是否在其他种族和地区人群的ARHL中发挥致病作用。
3 ARHL相关的临床研究
临床研究主要聚焦在ARHL相关的流行病学和风险因素,以及ARHL对大脑皮层和认知功能的影响。
3.1ARHL相关的流行病学研究 ARHL患病率随着年龄的增长而逐步增加,50岁以上人群约40%,70岁以上人群上升至约71%。与同龄女性相比,男性更容易遭受听力损失。与白人相比,黑人的听力损失风险降低,推测可能与黑色素水平的差异有关。
噪声暴露和耳毒性药物可能是加剧ARHL进展的外在因素(Yang等,2015)。吸烟和被动吸烟与听力损失的发生几率增加相关,与吸烟相比,被动吸烟的OR值更高(1.15∶1.28),提示被动吸烟导致听力损失的风险更高。除吸烟外,高体重指数(BMI)也与听力损失密切相关(Fransen等,2008)。另外,具有心血管危险因素(如高血压、糖尿病、吸烟、血清胆固醇升高)的个体听力损失的风险会更高。研究发现,在调整多种风险因素后,2型糖尿病与ARHL相关;与非糖尿病患者相比,糖尿病患者听力损失的患病率增加(Horikawa等,2013)。此外,ARHL也与社会经济地位(教育背景、职业和收入)存在一定关系,收入和教育水平较低的人的听力损失风险更高。但值得注意的是,多项研究发现适度饮酒与听力损失的几率降低相关,提示适度饮酒具有保护听力的作用(Gopinath等,2010),其深层的保护机制仍有待进一步深入研究。
值得注意的是,上述这些大样本人群的横断面调查研究结果仍存在一些局限性,因为整个队列随访研究过程中很难将个人风险因素的直接影响与对总体样本人群的间接影响有效区分,从而建立因果关系。然而,综合来看,这些数据表明,健康的生活方式,其中包括适度饮酒,可以起到一定的预防ARHL的保护作用。
3.2ARHL对中枢听觉皮层影响的相关研究 当外周听觉受损时,耳蜗将声信号转换为电信号的效能会降低,引起听觉信号减弱,进而影响大脑处理这些信息的方式,最终导致中枢听觉皮层的萎缩或重组[15]。在解剖学方面,主要表现为听觉皮层灰质体积减少[16]。有研究认为这种解剖变化与GABA水平降低导致的神经传递功能障碍有关(Gao等,2015)。灰质体积的整体减少是衰老的一种普遍性的神经特征,因此值得思考的一个问题是,ARHL引起的听觉传入剥夺是否会加剧脑萎缩的表现,以及是否会对皮层组织产生影响,这需要更深入的研究。
ARHL相关听皮层结构的变化,目前有以下两种理论假说:第一种,听觉障碍与听觉剥夺导致的脑容量下降之间存在直接的因果关系,也就是临床常讲到的听觉剥夺假说;第二种情况是,衰老本身就会导致听觉外周和中枢神经系统同时衰退[16]。
除了皮层的解剖结构变化外,ARHL患者在听觉处理的功能方面也表现出差异。功能性MRI(fMRI)研究显示,ARHL患者在颞叶,特别是右侧半球对粉红色噪声(即1/f噪声)的反应中表现出皮层功能激活增加。这种激活可能是由于与衰老相关的抑制作用减少,亦可能影响听阈升高的补偿机制(Chen等,2016)。
听觉信息的感知,尤其是理解,依赖于大脑网络之间的整合来理解和解释外周听觉刺激。ARHL患者参与听觉处理的大脑区域之间的功能连接存在重要差异,具体而言,ARHL患者视觉和听觉感觉皮层之间的连接,以及注意力和默认模式网络之间的连接显著减少(Puschmann等,2017)。说明在ARHL患者中负责对言语感知的整个皮层网络组织结构和功能发生了变化,进而对言语感知和理解产生阻碍。
3.3ARHL对中枢非听觉皮层影响的相关研究 ARHL不仅会影响听觉脑区,也会影响非听觉脑区。听力损失患者的非听觉脑区功能可能被上调以支持对言语的感知和理解,这种资源重组将进一步导致其认知和神经功能下降。
第一个影响表现在注意力网络方面。形态学研究表明,ARHL患者的前扣带皮层(ACC)体积减少。ACC萎缩与负责频率分辨的耳蜗外毛细胞的放大器功能之间存在关联,这种萎缩会进一步导致明显的记忆障碍[17]。也有研究认为,ARHL中受损的听觉处理会导致聆听更费力,进而逐步耗尽听觉和非听觉认知功能中可用的有限资源容量。通过研究参与努力感知声音的神经网络的激活增加现象或许有助于观察到ARHL患者中这些区域的神经退化[18]。
第二个影响表现在视觉网络方面。在听觉信号处理过程中抑制大脑其他感觉区域活动的能力会降低。例如,在单音节词识别任务中,当背景噪声增加引起言语可懂度降低时,视觉皮层的激活会明显增加(Vaden等,2016)。另外,静息状态下fMRI显示,ARHL患者中听觉和视觉皮层之间的神经网络连接也增加(Power等,2013)。这些研究进一步突出了与ARHL相关的视觉和听觉区域的皮层重组现象。
第三个影响表现在运动网络方面。听力损失越严重的患者,越需要依赖大脑多个区域的整合来理解听觉信息,但ARHL患者中目前尚不清楚运动网络是如何用于言语感知的。有学者提出以下两种假说来解释:首先,运动补偿假说认为运动网络的激活补偿了ARHL患者受损的听觉处理功能(Du等,2016);其次,运动衰退假说表明,外周听觉受损导致对听觉皮层的输入减少,听觉处理的缺陷进一步减少了对关节运动皮层输入的正向反馈[19]。
这些研究说明与ARHL相关的感觉剥夺会影响大脑结构、功能和典型的神经资源分配,这些结构的变化和神经资源的再分配重塑会影响ARHL患者的认知过程和听觉处理之外的其它功能。
3.4ARHL导致认知功能损伤的相关临床研究 ARHL与认知功能下降和患痴呆症的风险增加显著相关,并且这种情况会随着听力损失的严重程度而逐渐增加(Lin等,2013)。目前关于ARHL与认知功能下降的关联性,主要涉及以下三个假说:
(1)共同原因假说认为,认知功能下降和ARHL共病表现可归因于常见的神经退行性病变,这一假设得到了老年人的感觉功能和认知领域平行变化的研究证据支持(Fischer等,2016)。此外ARHL会加剧衰老过程中认知能力下降的进程和速度,信息退化和感觉剥夺假说同样支持这一观点[20]。
(2)信息退化假说则推测外周听觉受损引起听觉输入信号减少,导致听觉中枢对有限的处理资源提出了更高的要求(Wingfield等,2016)。在环境噪声或听力损失导致言语分辨率下降的情况下,大脑皮层处理和理解这些听觉信号的“听觉努力”会增加。因此,有限的认知资源被从其他认知任务中转移到努力聆听上,从而导致认知资源的耗尽。这种资源的重新分配对认知功能会产生不利的影响,在理论上可能导致认知功能下降(Humes等,2013)。当聆听更困难时,需要额外的认知资源来应对这种聆听需求,这就意味着用于其他认知过程的资源会逐渐被耗尽。支持这一假说的有力证据来自对助听器干预效果的研究,助听器有助于恢复听觉,进而减少认知负荷。助听器干预6个月可以显著改善患者听力和记忆力。
(3)感觉剥夺假说与信息退化假说存在一些理论上的共同点,但感觉剥夺假说更加强调ARHL的长期感觉剥夺,即随着时间的推移,认知资源长期向听觉重新分配会导致认知能力下降。该假说强调,这种长期剥夺会导致代偿性皮层重组和神经改变,从而阻碍一般的认知和情绪过程,但有利于听觉感知[21]。研究人员对感觉剥夺假说进行了扩展,提出尽管感觉剥夺通过感觉输入不足直接影响认知,但它也可能通过社交减少或抑郁增加间接影响认知功能。该假说提出,与社交孤立和抑郁相关的社交互动减少可能介导听力损失和认知能力下降之间的因果关系[22]。
4 ARHL预防与治疗相关研究
通过改变环境或饮食方式来预防ARHL的发生值得探索,然而其潜在机制仍未完全阐明。研究显示,重水(D2O)可调节新陈代谢,抑制啮齿类动物体内ROS的产生,延缓衰老的进程(Shchepinov等,2007)。笔者研究团队通过构建ARHL的小鼠模型,探讨了D2O对ARHL的影响,发现在4至9周龄的正常小鼠饮食中添加10% D2O可降低ABR反应阈,与未处理的小鼠相比,其蜗管中的氧化应激水平显著降低;代谢通量分析发现这一过程主要与D2O延缓与衰老相关的代谢反应有关。此外,实验还证实,Nrf2/HO-1/谷胱甘肽轴在D2O治疗的小鼠中下调,通过补充D2O可以减缓新陈代谢速度和减少耳蜗中氧化应激的产生,从而阻碍ARHL进展[23]。这些发现为保护耳蜗免受氧化应激和调节代谢以预防ARHL开辟了新的途径。
药物治疗方面也有一些新的发现。2013年辉瑞公司首次采用PF-04958242(AMPA受体的正变构调节剂,一种离子型谷氨酸受体)进行了ARHL的药物临床试验。选取44例50岁以上ARHL患者,单次给药后1 h和5 h听力测试发现听阈没有改善(Bednar等,2015)。在Autifony Therapeutics公司赞助并于2016年完成的CLARITY1研究中,评估了电压门控钾通道调节器AUT 00063对ARHL患者听力的影响,结果,在经过4周的治疗后,与安慰剂组比较,差异无统计学意义(注册号clinicaltrials.gov/ct2/show/NCT02345031,结果未发表)。诺华制药公司的一项I期和II期临床研究,采用基因治疗的方法,将促毛细胞转录因子ATOH1转导至重度至极重度聋患者内耳中,试图通过促进毛细胞从支持细胞再生来修复受损的听力(注册号clinicaltrials.gov/ct2/show/NCT02132130,结果未发表)。虽然动物试验显示这种方法能够恢复听力,但在人体内耳中是否有效尚未明确。此外,Pragma Therapeutics在噪声性听力损失(NIHL)动物模型中发现GRM7的调节剂能够改善ARHL的听力,但目前还在进行临床前试验,尚未应用到ARHL的前瞻性队列研究中[12]。
考虑到药物治疗ARHL的局限性,以及ARHL影响认知功能的现实依据,近年来较多学者开展了助听干预对ARHL认知功能下降影响的研究。一些研究发现,使用助听器干预能够预防或降低ARHL相关认知能力下降的速度(Mamo等,2017;Maharani等,2018),但需要进一步研究将其作为预防策略。另外,有研究发现,重度-极重度的ARHL患者植入人工耳蜗后认知能力明显提高[24]。虽然助听干预后整体认知能力的改善主要表现在即时和延迟记忆领域、工作记忆、处理速度和持续注意力,以及认知灵活性、注意力、配对关联学习和简单反应时间任务方面,但截至目前还几乎没有非常有力的证据能表明长期使用助听器或人工耳蜗植入后听觉相关中枢神经元会发生变化。另外,助听干预对认知功能的改善情况是否受听觉剥夺的年限、认知能力下降程度和痴呆分期等因素的影响,也无定论。这些临床问题都亟待深入研究,以期为ARHL相关认知功能下降的干预提供重要的理论依据,进而制定有效的防治策略。
5 ARHL防治任重道远
目前ARHL的确切病理机制尚不清楚,因此在药物研发方面仍缺乏精准的生物靶点,这也是目前ARHL药物治疗面临的最大挑战。研发可以预防或改善轻至中度ARHL的药物或许是目前更具现实前景的一个方向。虽然几项针对成人听力损失的药物治疗临床预试验和前瞻性队列研究仍在进行中,并显示出一些有效性,但仍需要更多的研究数据给予支持。此外,干细胞植入或基因靶向治疗,已经在动物研究中得到验证,但在人体内耳组织中,如何克服与内耳给药相关的解剖方面的生理屏障仍是目前治疗的最大瓶颈[25]。
随着人口老龄化速度的加快,听力损失和认知能力下降对幸福感和健康的影响也越来越受到社会关注。通过确定听力损失和认知能力下降之间潜在的介导因素及发病机制,可以确定早期干预的潜在途径,以缓解与ARHL相关的认知能力下降及老年痴呆的发生(Loughrey等,2008)。未来对ARHL产生的神经效应以及皮层重组和认知能力下降之间的因果关系的深入研究,将对ARHL相关健康问题干预策略的制定具有非常重要的意义。
或许可以通过WES和WGS捕获所有可能存在的变异,以阐明ARHL的遗传易感性。可通过GWAS和家族测序研究鉴定出更多的ARHL相关基因或等位基因,并进行纵向表型研究。此外,通过基因编辑技术(如CRISPR/Cas9)揭示基因改变后ARHL耳蜗的病理变化,或许可以为ARHL防治发挥关键作用。
总之,目前虽然鉴定了一些与ARHL易感性相关的候选基因,但对ARHL相关遗传学和病理机制的理解仍非常有限,并存在诸多挑战。尽管WGS正逐步用于ARHL的基因筛选,但缺乏足够的研究资金来进行强有力的表型研究,这是ARHL研究进一步进展的主要障碍。未来,ARHL防治仍然任重道远。
猜你喜欢
中国听力语言康复科学杂志(2021年6期)2021-12-21
中国临床医学影像杂志(2021年10期)2021-11-22
中国现代医药杂志(2020年3期)2020-05-08
基础医学与临床(2020年6期)2020-02-12
中华耳科学杂志(2020年6期)2020-01-08
中国生物医学工程学报(2019年6期)2019-07-16
中华耳科学杂志(2018年6期)2018-01-16
中外医疗(2016年15期)2016-12-01
海南医学(2016年8期)2016-06-08
听力学及言语疾病杂志(2015年5期)2015-12-24
