
我国梅树上病毒及类病毒的检测和鉴定
2024-02-06 14:39张煜泓徐惠媛战斌慧卢美光李世访张志想
植物保护 2024年1期
张煜泓 徐惠媛 战斌慧 卢美光 李世访 张志想




摘要
目前,我國梅树上的病毒种类及发生情况仍不完全清楚。本研究从北京、武汉、南京和无锡的梅园中采集了64份疑似感染病毒的叶片样品,通过RTPCR和斑点杂交,对7种病毒和2种类病毒进行了检测。共检测到6种病毒和1种类病毒。其中,李属坏死环斑病毒(prunus necrotic ringspot virus,PNRSV)和桃潜隐花叶类病毒(peach latent mosaic viroid, PLMVd)为我国梅树上的首次检出。PNRSV、亚洲李属病毒2号(Asian prunus virus 2, APV2)、桃叶痘伴随病毒(peach leaf pittingassociated virus, PLPaV)的检出率高于30%。综合考虑病毒的分布及检出率,PLPaV、APV2、PNRSV和李树皮坏死茎痘伴随病毒(plum bark necrosis stem pittingassociated virus,PBNSPaV)是武汉、南京和无锡梅树上的主要病毒。此外,通过克隆和测序,获得了PLMVd和梅树病毒A(mume virus A, MuVA)的基因组,PLPaV的RNA1组分和PNRSV外壳蛋白(CP)基因序列。序列比较分析显示,我国PLMVd梅分离物和PNRSV梅分离物与我国桃分离物亲缘关系最近,表明PLMVd和PNRSV可能在梅和桃树间交互侵染;我国MuVA梅分离物序列与日本梅分离物序列的相似性高达98.56%;PLPaV梅分离物与我国桃分离物之间序列变异较大。上述结果不仅进一步明确了我国梅树上的病毒及类病毒种类和分布情况,而且有助于深入了解它们的流行与传播。
关键词
梅树; 核果类果树; 病毒; 类病毒; 检测
中图分类号: S 436.619
文献标识码: A
DOI: 10.16688/j.zwbh.2022690
Detection and identification of viruses and viroids in Prunus mume Siebold & Zucc in China
ZHANG Yuhong, XU Huiyuan, ZHAN Binhui, LU Meiguang, LI Shifang, ZHANG Zhixiang*
(State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection,
Chinese Academy of Agricultural Sciences, Beijing 100193, China)
Abstract
The occurrence and distribution of viruses and viroids infecting Prunus mume Siebold & Zucc. in China remain partially understood. In this study, we collected 64 suspected leaf samples infected by virus from P.mume gardens in Beijing, Wuhan, Nanjing and Wuxi, and performed detection of seven viruses and two viroids using RTPCR and dotblot hybridization. Six virus species and one viroid species were detected from these samples. Among them, prunus necrotic ringspot virus (PNRSV) and peach latent mosaic viroid (PLMVd) were first identified in P.mume in China. PNRSV, Asian prunus virus 2 (APV2), and peach leaf pittingassociated virus (PLPaV) had a high infection rate of more than 30%. Based on geographic distribution and detection rates, PLPaV,APV2,PNRSV and plum bark necrosis stem pittingassociated virus (PBNSPaV) should be the main viruses in P.mume in Wuhan, Nanjing, and Wuxi. Furthermore, the nucleotide sequences of PLMVd and mume virus A (MuVA) genomes, RNA1 of PLPaV, and coat protein (CP) of PNRSV were obtained by cloning and sequencing. The results of sequence alignment showed that Chinese isolates of PLMVd and PNRSV were individually closest to their corresponding Chinese peach isolates, suggesting possible transmission between P.mume and peach trees. Moreover, Chinese P.mume isolate of MuVA had the highest similarity with Japanese isolate. Chinese isolate of PLPaV markedly differed from Chinese peach isolate. These findings further clarify the occurrence and distribution of viruses and viroids in P.mume in China, and are helpful for understanding the epidemic and transmission of these viruses and viroids.
Key words
Prunus mume; stone fruit tree; virus; viroid; detection
梅Prunus mume Siebold & Zucc.原产于我国南方,具有三千多年的栽培历史。川、黔、滇、藏交界处的横断山区和云贵高原一带不仅是梅的自然分布中心,也是梅的变异中心[1]。按用途梅可分为果梅与花梅(梅花)。果梅不仅营养丰富,而且可入药;花梅具有很高的观赏价值[1]。
梅是桃、李、杏等核果类果树的近缘种,在演变过程中,桃、李、杏的种质不同程度地掺入其间[1]。这些近缘的核果类果树可被一些相同病毒侵染[2],常见的如亚洲李属病毒(Asian prunus virus, APV)[3]、李痘病毒(plum pox virus, PPV)[4]、李树皮坏死茎痘伴随病毒(plum bark necrosis stem pittingassociated virus, PBNSPaV)[5]、李属坏死环斑病毒(prunus necrotic ringspot virus, PNRSV)[6]等。此外,这些病毒可在不同的核果类果树间交互侵染。因此,梅树病毒的研究对核果类果树病毒的防控及生态学研究具有重要意义。然而,同其他核果类果树相比,梅树病毒的研究较少。目前,已报道的侵染桃树的病毒(含类病毒)有30多种,而侵染梅树的仅有10余种[2]。
此前,利用RTPCR和高通量测序技术,从我国不同地区的梅树上检测到了APV1和APV2、PBNSPaV、PPV、苹果茎沟病毒(apple stem grooving virus, ASGV)、樱桃病毒A (cherry virus A, CVA)、梅病毒A (mume virus A, MuVA)和桃叶痘伴随病毒(peach leaf pittingassociated virus, PLPaV)[79]。但是,仍不确定我国梅树上是否存在国外已报道的其他病毒,如APV3、PNRSV、桃潛隐花叶类病毒(peach latent mosaic viroid, PLMVd)和啤酒花矮化类病毒(hop stunt viroid, HSVd)等。
在已报道的侵染我国梅树的病毒中,MuVA属乙型线状病毒科Betaflexiviridae发样病毒属Capillovirus,最早发现于日本梅树[10],新疆桃树上也有发生[11]。目前仅有日本梅分离物pm14(NC040568)和新疆桃分离物pp(MN412555)的基因组序列,而我国梅分离物的基因组序列仍未被测定。PLPaV属伴生豇豆病毒科Secoviridae蚕豆病毒属Fabavirus,最早发现于我国桃树[12],韩国桃树上也有发生[13]。PLPaV的基因组包含两条RNA(RNA1和RNA2)。我国桃分离物XJ6(KY867750)和韩国桃分离物BC(MK460243)的基因组序列已完成测定。梅是PLPaV的新寄主[8],但我国PLPaV梅分离物的基因组序列仍未被测定。
本研究对我国武汉、无锡、南京和北京梅树进行了7种病毒和2种类病毒检测,进一步丰富了侵染我国梅树的病毒种类。此外,分别测定了MuVA 和PLPaV的梅分离物的基因组序列,为这两种病毒的传播及生态学研究提供了基础数据。
1 材料与方法
1.1 材料
1.1.1 试供植物
从武汉、无锡、南京和北京的梅园中疑似感染病毒病的梅树上采集64份叶片样品,主要表现为叶片褪绿、花叶、脉明等症状[7, 9]。
1.1.2 主要试剂
PCR扩增所需的试剂购自北京全式金生物技术股份有限公司和生工生物工程(上海)股份有限公司;DNA凝胶回收试剂盒购自康宁生命科学有限公司;pMD18T和pTOPOBlunt载体购自北京冰达生物科技有限公司;SMARTer RACE 5′/3′ 试剂盒购自赛默飞世尔科技(中国)有限公司。
1.2 方法
1.2.1 疑似感病梅树样品的RNA提取及RTPCR
取新鲜疑似感染病毒的叶片0.1 g,采用CTAB法提取总RNA[14]。利用RTPCR技术检测7种侵染梅树的病毒和2种类病毒,引物信息见表1。
逆转录反应(RT):10 mmol/L dNTPs 2 μL,5×MMLV缓冲液4 μL,MMLV 逆转录酶0.5 μL,RNA酶抑制剂 (50 U/μL) 0.5 μL,Oligo (dT)1 μL,随机引物(N6)1 μL,RNA模板2 μL,灭菌去离子水补足至20 μL,混匀、短暂离心后,42℃温育60 min。
PCR:2×SanTaq PCR混合液10 μL,10 μmol/L上、下游引物各1 μL,cDNA模板1 μL,灭菌去离子水补足至20 μL。反应程序为:95℃预变性3 min;95℃变性30 s,退火温度下退火30 s,72℃延伸1 min,35个循环;72℃延伸10 min,12℃保存。
1.2.2 扩增产物的克隆及测序
使用凝胶回收试剂盒回收目的片段。将回收的目的片段与pMD18T或pTOPOBlunt载体连接后转入大肠杆菌DH5α感受态细胞。通过PCR鉴定阳性克隆,送至生工生物工程(上海)股份有限公司测序。
1.2.3 MuVA和PLPaV的基因組扩增
基于高通量测序从我国梅树样品中获得这两种病毒部分基因组序列[8],以及已报道的基因组设计引物(表2),利用RTPCR扩增获得除末端外的序列;利用RACE获得末端序列(具体步骤参考SMARTer RACE 5′/3′试剂盒说明书)。MuVA的3′末端无poly(A),扩增末端序列前,需在3′末端添加poly(A)。反应体系为:10×poly(A)聚合酶缓冲液5 μL,MnCl2 (1 mol/L) 5 μL,DTT (100 mmol/L) 5 μL,0.1% BSA 10 μL,ATP (10 mmol/L) 0.5 μL,RNA 10 μmol/L,poly(A)聚合酶2.5 μL,无RNA酶水补至50 μL。混匀后,37℃孵育30 min,反应结束后,通过乙醇沉淀的方法纯化产物。
1.2.4 斑点杂交
通过体外转录[14],合成地高辛标记的PLMVd负义链RNA,以此为探针检测梅树样品中的PLMVd。将提取的RNA直接滴加至杂交膜上,待膜干燥后,通过紫外线照射,交联固定RNA。然后进行杂交反应并采集图像,具体操作参考文献[14]。
2 结果与分析
2.1 梅树病毒的RTPCR检测
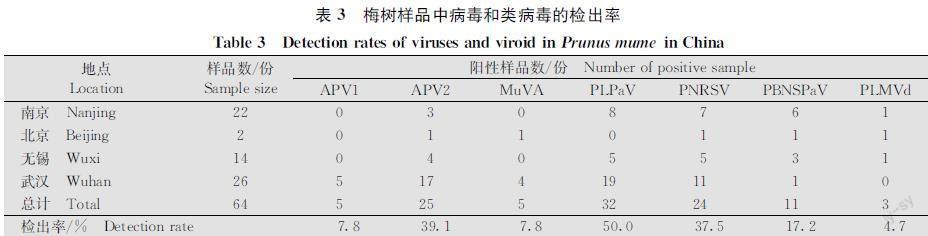
使用RTPCR对64份梅叶片样品进行7种病毒和2种类病毒的检测。除APV3和HSVd外,其余病毒均能检测到。其中,APV2、PLPaV和PNRSV的检出率较高,分别为39.1%、50.0%和37.5%;APV1、MuVA和PLMVd的检出率较低,分别为7.8%、7.8%和4.7%;PBNSPaV的检出率为17.2%(表3)。值得注意的是,南京、武汉、无锡的梅园中均检出APV2、PLPaV、PNRSV和PBNSPaV,且检出率较高。表明它们是这些地区梅树上发生的主要病毒。另外,10%的样品被2种及以上的病毒复合侵染。其中,武汉梅园的4个样品被MuVA、PLPaV、APV1和APV2复合侵染。
2.2 PLMVd的检出情况及序列分析
使用RTPCR从南京、北京和无锡的梅样品中检测到了PLMVd(图1a)。这是首次从我国梅树上检出PLMVd。斑点杂交检测进一步验证了该结果(图1b),说明我国梅树上的确存在PLMVd。
通过克隆和测序,从无锡、北京和南京的样品中各获得1条PLMVd基因组序列(OL854072~OL854074),长度均为338 nt,相互之间的核苷酸序列相似性为97.06%。与已报道的序列相比较,无锡、北京和南京的序列分别与我国桃分离物PLMVdT041(MH974835)、PLMVdT052(MH974838)和PLMVdG(EF591868)最接近,序列相似性分别为99.41%、99.70%和96.13%。表明我国梅树上的PLMVd与我国桃树上的PLMVd具有较近的亲缘关系。
2.3 PNRSV的检出情况及序列分析
通过RTPCR从我国梅树上检测到了PNRSV。这是首次从我国梅树上检出PNRSV。为进一步验证该结果,通过克隆和测序,从不同的梅叶片样品中获得了12条PNRSV外壳蛋白(CP)的基因序列,
它们之间的核苷酸序列相似性高达99.14%。与已报道的序列相比较,这12条序列与我国桃分离物Pcha(HQ833194)的核苷酸序列相似性最高,为99.56%~100%。表明我国梅树上的PNRSV与我国桃树上的PNRSV具有较近的亲缘关系。
2.4 我国梅树上MuVA分离物的基因组序列
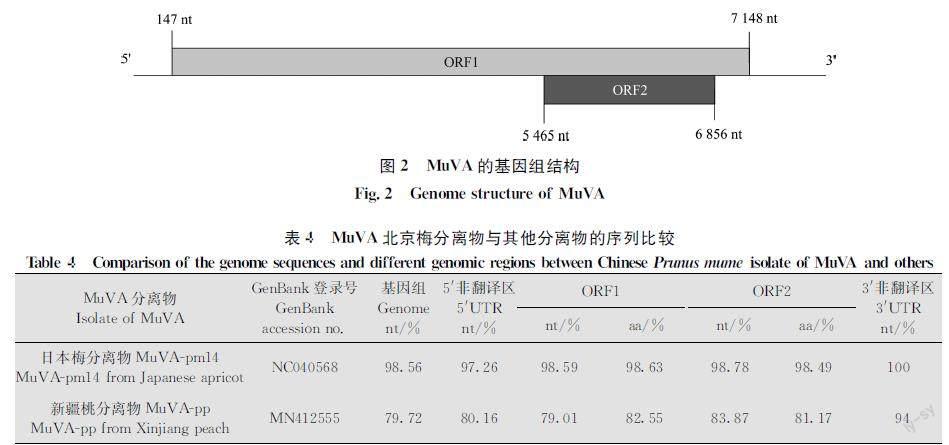
获得了我国梅树上MuVA分离物(MuVABJ)的基因组序列。全长7 644 nt,包含两个开放阅读框(open reading frame, ORF)(图2),ORF1(147-7 148 nt)编码多聚蛋白(polyprotein),含2 333个氨基酸,分子量约为265.4 kD;ORF2(5 465-6 856 nt)编码移动蛋白(putative movement protein,MP),含463个氨基酸,分子量约为51.3 kD(图2)。基因组结构与日本梅分离物MuVApm14(NC040568)[10]的一致。
将MuVABJ的基因组序列与日本梅分离物MuVApm14和我国新疆桃分离物MuVApp(MN412555)进行比较(表4),其与日本梅分离物
的核苷酸序列相似性高达98.56%,而与新疆桃分离物的核苷酸序列相似性仅为79.72%。ORF1和ORF2的核苷酸和氨基酸序列比较的结果与此相似。表明MuVA可能具有寄主特异性。
2.5 我国梅树上PLPaV分离物的基因组序列
经克隆和测序,获得了PLPaV武汉梅分离物(PLPaVJM3)的RNA1组分的序列,长度为6 085 nt,包含1个ORF(3-5 765 nt),编码一个多聚蛋白,由1 920个氨基酸组成,分子量约为217.3 kD。将其与已报道的两个分离物——我国桃分离物PLPaVXJ6(KY867750)和韩国桃分离物PLPaVBC(MK460243)的RNA1组分相比较,核苷酸序列相似性分别为86.62%和85.46%,氨基酸序列相似性分别为96.87%和96.08%。此外,还获得了PLPaVJM3的RNA2的部分序列,长度为848 nt。其与我国桃分离物PLPaVXJ6和韩国桃分离物PLPaVBC的 RNA2组分的核苷酸序列相似性分别为96.00%和84.14%。
3 结论与讨论
本研究通过RTPCR和斑点杂交检测,不仅从我国梅树上首次检测到了PNRSV和PLMVd,而且分析了6种病毒(APV1、APV2、MuVA、PBNSPaV、PLPaV、PNRSV)和1种类病毒(PLMVd)在北京、武汉、南京和无锡梅园的檢出情况,发现APV2、PLPaV、PNRSV和PBNSPaV在武汉、南京和无锡梅园中有发生,且检出率较高,应该是这些地区梅树上的主要病毒。此外,首次测定了我国梅树上的PLMVd、MuVA和PLPaV的基因组序列。这些结果不仅进一步明确了我国梅树上病毒的分布情况,而且为亲缘关系相近的核果类果树病毒的遗传及生态学研究提供了数据。
目前,已报道的侵染梅树的病毒有16种[2,89,1517]。此前,已经从我国梅树上检测到8种病毒(APV1、APV2、ASGV、CVA、MuVA、PBNSPaV、PLPaV和PPV)[79],本研究中又检测到PLMVd和PNRSV。由此可知我国梅树上至少存在9种病毒和1种类病毒。我国梅树上是否存在国外已报道的其余6种病毒及类病毒——苹果褪绿叶斑病毒(apple chlorotic leaf spot virus, ACLSV)、桃花叶病毒(peach mosaic virus, PcMV)[2]、油桃茎痘伴随病毒(nectarine stem pitting associated virus,NSPaV)、杏脉明伴随病毒(apricot vein clearing associated virus, AVCaV)[18]、APV3[3]和HSVd[16],仍有待检测和验证。
本研究发现APV2、PLPaV和PNRSV的检出率高于30%,且在武汉、无锡和南京的梅园中均有发生。此前的调查发现ASGV的检出率也高于30%,且在武汉、无锡、上海和南京的梅园均有发生[9];PPV的检出率更高且分布更广泛[7]。综上,我国梅树上的主要病毒有APV2、ASGV、PLPaV、PNRSV和PPV。
在侵染我国梅树的主要病毒中,ASGV是侵染我国苹果的主要病毒之一[19],PLPaV和PNRSV是侵染我国桃树的主要病毒[20]。推测梅树可能是这些病毒在我国核果类果树中传播和扩散的重要寄主。此外,本研究首次从我国梅树上检测到了PLMVd,其也是我国桃树上的主要类病毒[20],而且梅树分离物与我国桃树分离物的亲缘关系最近。表明PLMVd在我国梅树和桃树间可能存在交互侵染。因此,防控这些病毒或类病毒不能忽略梅树病毒的研究。
虽然本研究检测的梅树样品大多来自花梅,但仍有部分样品来自果梅,而且从中检测到了不同的病毒。因为病毒侵染会危害果梅的生产[6, 21],不仅会引起叶斑、坏死等症状,还会降低产量。即使目前国内尚未有果梅上的病毒病危害的报道,仍需加强病毒的检测和监测。
参考文献
[1] 褚孟嫄. 中国果树志梅卷[M]. 北京: 中国林业出版社, 1999.
[2] RUBIO M, MARTINEZGOMEZ P, MARAIS A, et al. Recent advances and prospects in Prunus virology [J]. Annals of Applied Biology, 2017, 171(2): 125138.
[3] MARAIS A, SVANELLADUMAS L, FOISSAC X, et al. Asian prunus viruses: New related members of the family Flexiviridae in Prunus germplasm of Asian origin [J]. Virus Research, 2006, 120(1/2): 176183.
[4] RODAMILANS B, VALLI A, GARCIA J A. Molecular plantplum pox virus interactions [J]. Molecular PlantMicrobe Interactions, 2020, 33(1): 617.
[5] NAKAUNE R, TAKEDA T, NUMAGUCHI K, et al. Ampeloviruses associated with incomplete flower syndrome and leafedge necrosis in Japanese apricot [J]. Journal of General Plant Pathology, 2018, 84(3): 202207.
[6] KURIHARA J, TOMARU K, OTSUBO T, et al. A new disease of Prunus mume (Japanese apricot) caused by multiple infection with cucumber mosaic cucumovirus and prunus necrotic ringspot related ilarvirus [J]. Acta Horticulturae, 1998, 472: 183193.
[7] ZHOU Ju, XING Fei, WANG Hongqing, et al. Occurrence, distribution, and genomic characteristics of plum pox virus isolates from common apricot (Prunus armeniaca) and Japanese apricot (Prunus mume) in China [J]. Plant Disease, 2021, 105(11): 34743480.
[8] ZHANG Yuhong, ZHOU Jun, ZHAN Binhui, et al. First report of peach leaf pittingassociated virus, plum bark necrosis stem pittingassociated virus, and mume virus A from mei (Prunus mume) in China [J]. Plant Disease, 2021, 105(8): 2259.
[9] 王程安, 周俊, 陳梦婷, 等. 我国部分地区梅树病毒病的病原初步检测[J]. 植物保护, 2018, 44(2): 170176.
[10]MARAIS A, FAURE C, THEIL S, et al. Molecular characterization of a novel species of Capillovirus from Japanese apricot (Prunus mume) [J/OL]. Viruses, 2018, 10(4): 144. DOI: 10.3390/v10040144.
[11]ZHENG Yinying, BU Fangdi, WU Caijie, et al. First report of mume virus A infection of Prunus persica in China [J]. Plant Disease, 2020, 104(10): 2741.
[12]HE Yan, CAI Li, ZHOU Lingling, et al. Deep sequencing reveals the first Fabavirus infecting peach [J/OL]. Scientific Reports, 2017, 7(1): 11329. DOI: 10.1038/s41598017117437.
[13]JO Y, CHOI H, BACK C K, et al. First report of peach leaf pittingassociated virus in peach trees exhibiting calico in Korea [J]. Plant Disease, 2019, 103(11): 29712972.
[14]张志想, 葛蓓孛, 潘嵩, 等. 菊花矮化类病毒的分子检测与序列分析[J]. 园艺学报, 2011, 38(12): 23492356.
[15]CANDRESSE T, FAURE C, THEIL S, et al. First report of nectarine stem pittingassociated virus infecting Prunus mume in Japan [J]. Plant Disease, 2017, 101(2): 393.
[16]JO Y, LIAN S, CHO J K, et al. First report of hop stunt viroid in Japanese apricot (Prunus mume) in Korea [J]. Plant Disease, 2017, 101(7): 1332.
[17]OSAKI H, YAMAGUCHI M, SATO Y, et al. Peach latent mosaic viroid isolated from stone fruits in Japan [J]. Annals of the Phytopathological Society of Japan, 1999, 65(1): 38.
[18]AL RWAHNIH M, SOLTANI N, SOLTERO B R, et al. First report of apricot vein clearingassociated virus infecting flowering apricot (Prunus mume) in the United States [J]. Plant Disease, 2021, 105(9): 2739.
[19]郝璐, 叶婷, 陈善义, 等. 我国北方部分苹果主产区病毒病的发生与检测[J]. 植物保护, 2015, 41(2): 158161.
[20]周俊. 桃及其野生近缘种中病毒组研究[D]. 北京: 中国农业科学院, 2020.
[21]NUMAGUCHI K, TAKEDA T, TSUCHIDA Y, et al. Largescale field survey reveals overall yield loss in Japanese apricot possibly caused by two ampeloviruses [J]. Journal of General Plant Pathology, 2019, 85(2): 116121.
(責任编辑:杨明丽)
猜你喜欢
当代水产(2022年1期)2022-04-26
小主人报(2022年5期)2022-04-01
椰城(2021年12期)2021-12-10
今日农业(2021年16期)2021-11-26
今日农业(2021年21期)2021-11-26
中国诗歌(2018年3期)2018-11-14
农家科技中旬版(2018年5期)2018-07-14
学生阅读世界(喜欢写作文)(2017年11期)2017-02-15
小学生导刊(2016年13期)2016-12-01
西南农业学报(2016年5期)2016-05-17
