
绿藻产氢的机制及研究进展
2024-02-27 02:05王婧怡贾蒲连付爱根
生物学杂志 2024年1期
王婧怡 , 赵 琪, 贾蒲连, 付爱根, 王 菲
(1. 西北大学生命科学学院, 西安 710069; 2. 西北大学榆林碳中和学院, 西安 710069)
随着全球人口增长和经济飞速发展,人类对资源的需求也日益增长。煤炭等化石燃料的大量使用导致温室气体浓度增加,引发全球气候变暖以及极端天气频发等环境问题。因此,开发可持续、无污染的新能源成为全球科学家重点研究的方向。
氢气具有高效、清洁和热值高等优点,被认为是最有价值的可再生能源。传统的制氢方式包括化石燃料的重新转化和电解水等,但这些方法并不能从根本上解决能源短缺和环境污染等问题。绿色生物制氢作为当今新能源领域的热门方式亟待发展,有助于早日实现“碳达峰、碳中和”的目标。
生物制氢是通过多种微生物(如微藻、光合细菌和一些异养细菌等)的不同代谢途径产生氢气。微藻繁殖速度快且易培养,并具有固定二氧化碳的能力,其通过太阳能利用水生产氢气,成本低且资源广泛,是绿色工厂生产氢能源的首选物种。光合产氢的微藻主要分为真核绿藻和原核蓝藻两大类,它们以太阳光为能量来源,以水为电子供体,通过体内氢化酶的催化作用产生氢气和其他生物燃料,并且光合产氢过程不会产生温室气体和其他有害物质,是目前最理想的生物制氢途径。相比蓝藻,绿藻光合产氢效率更高,目前多以绿藻作为生物产氢的研究和应用对象[1]。
1 绿藻光合产氢的作用机制
绿藻的光合作用发生在叶绿体的类囊体膜上,其光合系统包括光系统I(PSI)和光系统II(PSII),PSII利用太阳能裂解水产生氧气和电子,而PSI利用太阳能和电子生成还原型辅酶II(NADPH)。Gaffron等[2]首次报道斜生栅藻可以产生氢气,随后莱茵衣藻的产氢能力也得到了证实,并成为光合产氢模式藻种[3]。目前已知的绿藻产氢途径有3种:依赖于PSII的直接产氢、不依赖于PSII的间接产氢和暗反应发酵途径(图1)。前两种产氢途径通过绿藻光合电子传递链调节,后者主要与发酵代谢有关。

图1 绿藻产氢的3种途径(根据文献[4]修改绘制)Figure 1 The hydrogen production pathways of green algae(Modified from [4])
1.1 依赖PSII的直接产氢通路
绿藻利用类囊体膜表面的捕光色素吸收太阳光,PSII的反应中心将水分解为质子和O2,并释放电子,随后电子进入光合电子传递链,经过质体醌(PQ)、细胞色素b6f复合体(Cytb6f)、质体蓝素(PC)和PSI,再传递给铁氧还原蛋白(Fd),还原的Fd将电子分配给NADPH铁氧还蛋白还原酶FNR(主要电子受体)或氢化酶(次级电子受体),氢化酶接受Fd传递的电子后,使质子还原为氢气。在厌氧条件或捕光色素吸收过多能量时,绿藻会利用氢化酶消耗多余的电子,从而避免光抑制并保护光合电子传递链[5]。直接产氢途径的优势在于利用水作为原料,阳光作为驱动能源,缺点是光合作用同时也会产生氧气,从而抑制氢化酶的活性,使产氢时间较短。
1.2 不依赖PSII的间接产氢通路
不依赖PSII的间接产氢途径一般包括两个阶段:(1)在光合作用期间积累碳水化合物,光合作用产生的还原力先被储存在淀粉中;(2)在淀粉分解代谢的过程中产生氢气。首先,绿藻光合作用生成的ATP和NADPH进入卡尔文循环,被用于二氧化碳的固定进而生成淀粉[6],接着在厌氧条件下,淀粉分解产生的NADH在II型NADH脱氢酶的催化作用下被氧化,同时释放的电子通过PQ进入光合电子传递链,最后传递给氢化酶,产生氢气[7]。不依赖PSII的间接产氢通路将光合裂解水阶段与光合产氢阶段分隔开,能够有效防止氧气对氢化酶活性的影响。但与直接产氢途径相比,间接产氢通路的产氢量较低,且受到恒定光源和ATP需求的限制而无法应用到大规模生产中。
Melis等[8]在此基础上发展了“两步法”诱导产氢,先将莱茵衣藻在以乙酸盐为碳源的光合异养培养基(TAP培养基)中培养至对数生长期,随后将其转移至缺硫的培养环境中,硫元素的缺乏使衣藻细胞内PSII核心亚基D1蛋白合成减少,光合速率降低,但呼吸作用不受影响,最终在细胞内形成厌氧环境,解除了氧气对氢化酶活性的抑制作用,持续催化氢气产生[9]。
1.3 暗发酵产氢
除了上述两种情况,微藻还能够在黑暗条件下通过分解有机物产生氢气,这一途径称为暗发酵产氢。该途径无需光能和水,且能够持续产氢6 h以上,但产氢量远远低于光合产氢量[10],因为在暗发酵过程中复杂的有机物需要经过水解、酸化、乙酰化等多个阶段的转化,才能形成氢气和脂肪酸等其他中间代谢产物。微藻暗发酵产氢的电子主要来源于丙酮酸铁氧还蛋白酶(PFR1)催化丙酮酸的氧化,这一过程与Fd的还原偶联在一起,最终Fd将电子转移到氢化酶而产生氢气[11]。该途径一般会受到发酵产物积累的限制,特别是乙酸会损害细胞生长从而影响氢气的产生,因此,控制pH值是提高暗发酵产氢途径效率的关键[12]。此外,厌氧发酵前对底物进行预处理也可以增加氢气产量[13]。
2 绿藻氢化酶及产氢的限制因素
2.1 [FeFe]-氢化酶
藻类产氢的关键酶是氢化酶和固氮酶,大多数固氮酶存在于蓝藻中,其作用是促进N2转化为NH3同时释放出副产物氢气。氢化酶根据活性位点结构不同,可分为[NiFe]-氢化酶、[FeFe]-氢化酶和[Fe]-氢化酶。[NiFe]-氢化酶广泛存在于原核生物中,催化氢气的释放和吸收;[FeFe]-氢化酶存在于一些真核细胞和厌氧细菌中,只催化氢气释放[14];[Fe]-氢化酶与其他两种酶的功能不同,它是将氢气分解成H+和H-,仅存在于产甲烷菌中[15]。绿藻产氢的核心酶——[FeFe]-氢化酶具有独特的活性中心——H簇,其催化活性比[NiFe]-氢化酶高出100多倍[16]。[FeFe]-氢化酶对氧气极其敏感,只有在厌氧的环境下才具有催化活性,并且氧气对该酶的抑制作用是不可逆的[1]。莱茵衣藻有两种[FeFe]-氢化酶:HydA1和HydA2。Meuser等[17]研究发现,HydA1是主要的氢化酶,其对衣藻光合放氢量的贡献约占75%,HydA2约占25%。
2.2 产氢的限制因素
限制绿藻大规模工业化产氢的主要因素有:(1)氧气对氢化酶活性的抑制作用;(2)卡尔文循环与氢化酶竞争电子。此外,绿藻产氢是多个调节因子共同参与的复杂生理过程,如光照强度、pH值、温度、底物浓度和生物反应器的配置等均是影响产氢效率的因素。
2.2.1 氢化酶对氧气的敏感性
氢化酶对氧气非常敏感,在早期研究中,研究人员曾尝试利用含有血红蛋白或连二亚硫酸盐的吸氧剂和氧清除剂降低培养体系中氧气含量,然而此类技术并不能完全克服氧气对氢化酶的限制。
藻类的生长需要营养成分。任何微量营养素或常量营养素的缺乏都会直接影响藻类的生长状态。因此,研究人员利用营养缺陷的方法去影响细胞代谢,进而减少氧气对氢化酶的抑制。在光照条件下用缺硫TAP培养衣藻大约24 h后,光合作用放氧速率小于呼吸作用耗氧速率,使密闭的培养系统逐渐达到厌氧状态,从而有利于氢气的产生。在这期间,缺硫导致PSII中的D1蛋白合成受损,电子由淀粉等内源底物的分解代谢产生[18]。短时间缺硫可以影响光合放氧同时不影响细胞其他的生理代谢功能,但缺硫时间过长时,细胞内开始积累大量有毒物质[4],细胞的产氢速率会逐渐降低直至停止产氢。之后的一些实验发现,培养基缺少氮、磷和镁等元素也会引起其他代谢途径发生变化从而造成PSII逐渐失活,细胞呼吸耗尽氧气进而使氢化酶活性提高,然而缺硫产氢仍然是目前最有效且最实用的诱导衣藻光合产氢的方法[19]。
2.2.2 能量转化效率
绿藻的产氢效率受温度、pH值、光照强度、碳水化合物含量等因素的制约。在众多因素中,pH是直接影响绿藻产氢效率的重要参数。因为pH不仅会影响绿藻的细胞活力,还会影响氢化酶的活性。Kosourov等[20]对莱茵衣藻进行缺硫处理,初始pH值为7.7时产氢量最大,而pH值降至6.5或升高至8.2时,产氢量降低。此外,温度也是一个重要因素,微藻在15~35 ℃的温度范围内生长状态最佳[21]。藻的细胞密度也是影响绿藻产氢的关键条件,Lakatos等[22]研究发现,在初始藻细胞密度为3.96×108个藻细胞/mL时,累积的总产氢量达(88.98±2.18) mL/L,随着细胞密度增加,氢气产量迅速下降。光照强度也是制约产氢效率的因素之一,绿藻吸收的光能不仅促进水分子的氧化、电子和质子的释放,还为电子向Fd的传输提供驱动力,因此,通常光照强度越低,氢气的产量也相对越低。有研究发现短暂的高光照射可以通过抑制光合放氧来增加氢气的产生速率[23]。除此之外,在厌氧培养阶段提高光强,叶绿素浓度增加,释放的电子数量也随之增加,这些电子能够与质子结合形成更多的氢分子[8]。同时,为了提高光子转换效率,还可以设计合适的光反应器来增加光的摄入量。碳水化合物含量也是一个限制产氢的重要因素。Kosourov等[24]发现莱茵衣藻在缺硫处理后,细胞淀粉含量在放氧阶段大量积累,而在产氢阶段明显下降。Posewitz等[25]提出淀粉代谢在莱茵衣藻产氢过程中占据着非常重要的地位。淀粉代谢既参与依赖PSII的直接途径,也参与不依赖PSII的间接途径。前者淀粉降解为呼吸作用提供还原力,支撑自身的厌氧代谢活动;后者淀粉降解产生的还原当量(NADH)被氧化后进入光合电子传递链,为氢化酶提供电子。Kosourov等[26]监测绿藻在光合自养条件下的淀粉含量变化,发现其与产氢量呈正相关。这也为提高绿藻产氢效率提供了一个新方向。
3 提高绿藻产氢的策略
受多种条件限制,目前绿藻光合产氢的效率远远达不到工业化应用的要求。为了提高产氢效率,研究人员们利用各种手段对绿藻进行改造和优化,本节重点概述7种提高产氢效率的方法(图2)。
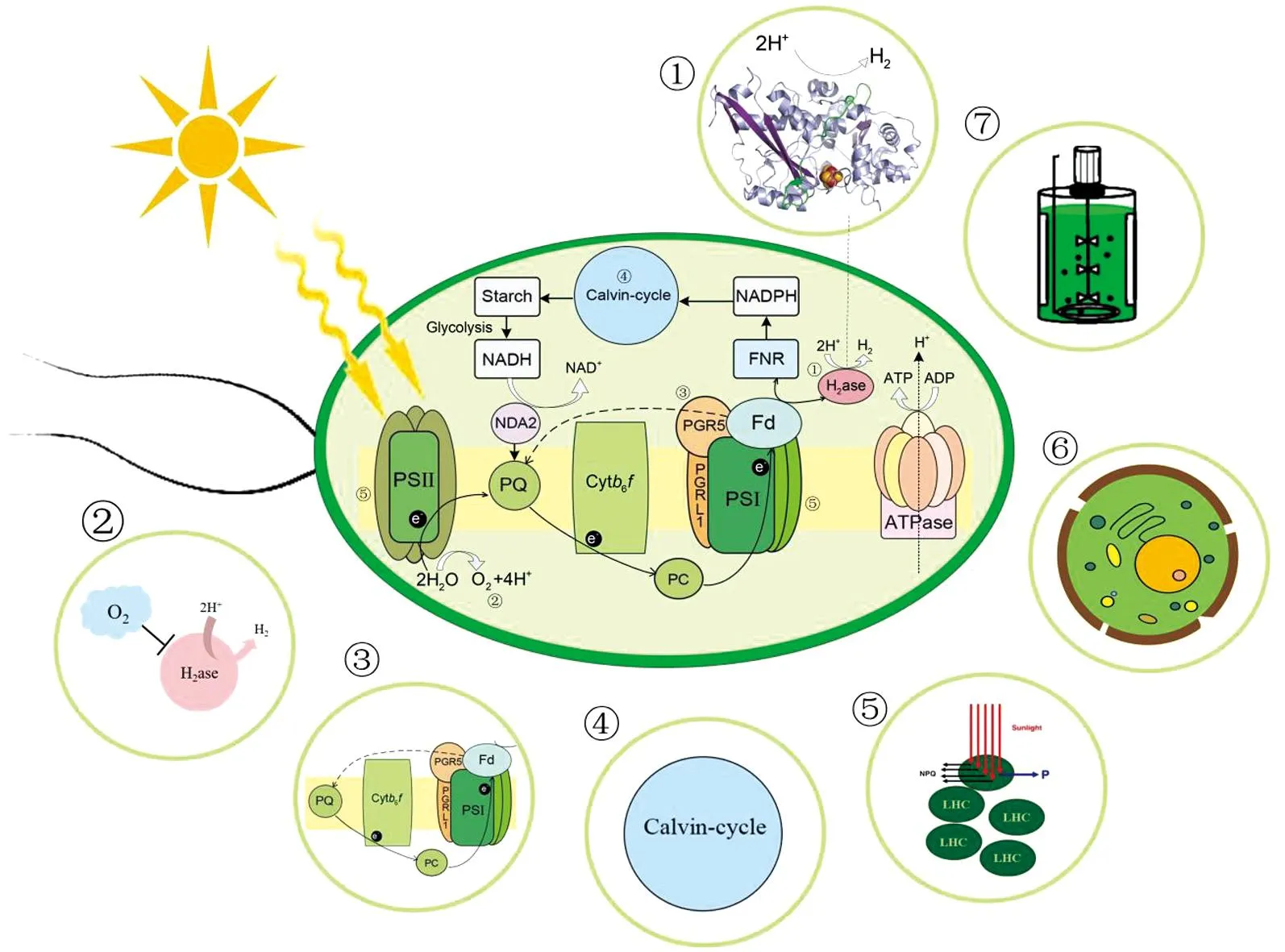
①氢化酶改造;②降低体系中氧气的含量;③减弱环式电子传递;④降低卡尔文循环;⑤缩小捕光天线;⑥底物预处理;⑦光生物反应器设计。图2 提高绿藻产氢效率的策略Figure 2 The strategies to improve hydrogen production efficiency of green algae
3.1 氢化酶改造
研究人员通过对氢化酶进行遗传改造来提高氢化酶对氧的耐受力和对电子的竞争力。Eilenberg等[27]将电子供体铁氧还蛋白Fd与HydA进行融合并转入莱茵衣藻中,发现该融合酶改变了细胞内特定的代谢途径,使更多的电子流向氢化酶并表现出更高的耐氧性。Noone等[28]将梭状芽胞杆菌的[FeFe]-氢化酶基因CaI的密码子进行优化,融合到[FeFe]-氢化酶表达缺陷的莱茵衣藻PsaD启动子上(该启动子调控光系统I的PsaD亚单位表达),CaI在D66ΔHYD中成功表达并延长了产氢时间,这为改造获得更耐氧气的氢化酶提供了新的思路。Redding等[29]设计出PSI亚基与氢化酶融合形成的嵌合蛋白,该蛋白具有优势竞争电子的能力,能够从PSI的PsaC亚基FB簇截取电子,使更多的电子流向产氢途径。Yang等[30]对来自小球藻的HydA(CsHydAc)结构进行遗传改造,通过修饰气体隧道周围的氨基酸残基A105I、V265W、G113I或V273I,以限制氧气进入酶活性部位,当培养至第6天,突变藻株(DT-AIVW-27)产氢量迅速增加,到第11天其最大产氢量已达到70 μL/mL,为野生型的30倍。
3.2 降低体系中氧气的含量
一些研究人员致力于通过降低细胞环境中的氧气浓度而解除氧气对衣藻产氢的制约,提高光合氢的产生效率。
藻类和细菌共培养降低氧气水平。在自然界中,一些细菌与藻类形成互惠关系,绿藻光合作用产生的氧气及碳水化合物可以支持兼性厌氧菌的生长,而细菌消耗氧气使绿藻氢化酶活性提高,同时细菌又可以为藻类的生长提供必需的生长因子和CO2。实验证明,藻类和细菌共培养比单独培养的产氢量多出约50%~60%[33]。Yu等[34]将莱茵衣藻与根瘤菌共培养,发现在不同氮源条件下,莱茵衣藻的呼吸速率、叶绿素含量、淀粉积累量以及产氢量均有不同程度提高。此外在菌藻共培养的最佳比例下,向体系中间歇添加氮化物,可以实现持续近55 d产氢,最大产氢量达到226.98 μmol/mg Chl,是对照组的5.2倍。Javed等[35]通过共培养小球藻和活性污泥中的细菌可使得其产氢量为纯细菌菌株的7倍。
绿藻聚集体的光合产氢新方法可以替代缺硫培养和氧气清除剂的使用。绿藻细胞聚集体的形成会使里层细胞的光合作用因缺乏光照而受到抑制,但其呼吸作用可正常进行,氧气被逐渐耗尽,最终在聚集体内部形成一个有利于产氢的厌氧环境[36]。绿藻聚集体产氢在常规的培养基中进行,不需要其他处理,因此,适合低成本大规模应用。研究人员证明了化学絮凝能够分离和浓缩绿藻细胞形成聚集体,并发现可以通过商业化学絮凝剂——阳离子醚化淀粉,在原始液体培养中直接形成聚集体,从而实现11 d的持续产氢[37]。
3.3 减弱环式电子传递
莱茵衣藻的光合电子传递链可分为线性电子传递(LEF)和环式电子传递(CEF)。在正常生长状态下电子传递以LEF为主,当LEF产生的ATP少于固定CO2所需的ATP时,光合作用将依赖于围绕PSI的环式电子传递。藻类中存在两种不同的环式电子传递途径:(1)(NADH脱氢酶)NDH途径;(2)PGR5/PGRL1介导途径。在厌氧条件下,PGR5/PGRL1介导的环式电子传递能力远高于NDH介导的环式电子途径[38]。产氢过程中,卡尔文循环水平较低,环式电子传递占主导,环式电子传递链不但与氢化酶竞争电子,而且在此过程中产生的跨膜质子梯度也可能不利于氢气的产生。因此,减弱环式电子传递在增强莱茵衣藻光合产氢效率方面有重要意义。Chen等[39]鉴定出缺失PGR5基因的突变株hpm91,该突变株在缺硫条件下能连续25 d产氢,产氢量可达野生型的30倍。Elman等[40]发现缺失PGR5基因的突变体可以在1 L的光合混养条件下连续产氢12 d,并探究出该突变体是通过增强叶绿体和线粒体之间的联系从而实现在简单的大规模培养体系中持续产氢。
3.4 降低卡尔文循环
除了环式电子传递,线性电子传递的FNR和卡尔文循环也是与氢化酶竞争Fd处电子的主要因素。由于Fd对FNR具有很高的亲和力,光合作用产生的绝大多数电子直接从Fd传递给FNR产生NADPH,用于CO2的固定。Elman等[40]在莱茵衣藻混养体系中加入磷酸核酮糖激酶的抑制剂——羟基乙醛,从而使卡尔文循环受到抑制,产氢速率大幅提高。为了将更多的电子引导到氢化酶靶点,Günzel等[41]分析了莱茵衣藻两种氢化酶HydA1和HydA2与不同植物型铁氧还蛋白异构体的相互作用图谱,发现铁氧还蛋白Fdx7对HydA1的亲和力高于对FNR的亲和力,这一发现为定制电子供体提供了新的方向。此外还有研究人员利用RNAi的方法得到FNR水平降低的衣藻突变株,发现FNR下调引起光合放氧速率下降、RuBisCO含量减少以及淀粉降解率提高的现象,这些现象可能导致FNR-RNAi藻株的零氧期提前或氢化酶的电子供应增加,从而实现产氢量的提高[42]。
3.5 缩小补光天线
光捕获是光合产氢的第一步,已有研究表明,捕光天线的存在会导致强光下微藻的能量过度吸收和浪费。减小天线尺寸可以提高光能转化效率,进而提高衣藻产氢效率。Ho等[43]证明PSI补光天线蛋白LHCA2的缺失会影响PGR5介导的光合电子传递,使更多电子流向氢化酶,从而促进光驱动氢气的产生。pgr5/lhca2双突变体的产氢量是pgr5突变体的两倍。Ban等[44]利用常压室温等离子体(ARTP)突变技术截断莱茵衣藻的叶绿素天线,使得突变体单位细胞体积的叶绿素含量降低,该突变体产氢量达到野生型的1.8~5.2倍,说明叶绿素天线的缩小提高了光合作用效率,从而促进了氢气的产生。Lu等[45]研究表明缩小补光天线会降低活性氧(Reactive oxygen species,ROS)的产生,有助于减少产氢过程中氧化损伤对莱茵衣藻的影响,从而提高衣藻的产氢效率。
3.6 底物的预处理
微藻的生物质含量极其丰富,为了促进碳水化合物的解聚和提高微藻的生物制氢能力,通常会进行适当的预处理以破坏细胞结构的完整性,释放储存的碳水化合物。底物的预处理技术,如化学法、热处理、机械法、酶处理法以及这些技术的组合,已被证明是提高氢气产量的有效方法。Liu等[46]先对底物质量为6 g的小球藻进行NaOH处理,再将其进行酶解,发现产氢效果要好于直接酶解,可以将产氢量从每克干重22.0 mL提高至49.1 mL。Ortigueira等[47]通过对斜生栅藻进行碾磨将总产氢量从每克干重32 mL提高到116 mL。未来针对联合预处理方法的探索将有望开发高效、能源密集型、成本效益高的生物质水解混合预处理方式[48]。
3.7 光生物反应器的优化
优化用于扩大培养的光生物反应器也是提高绿藻产氢效率的手段之一。光生物反应器(photobioreactors,PBR)是指能够培养光合微生物以及具有光合能力的细胞或组织的装置[49]。根据微藻生长环境是否与外界直接接触,可分为开放式PBR和封闭式PBR两大类。开放式PBR成本低,容易操作。封闭式PBR可以有效控制培养条件,减少污染。然而,目前现有的光生物反应器都有设备投资和运行成本高且寿命相对较短的缺点,因此,开发经济可行的PBR系统是实现高效产氢的重要途径。Salguero-Rodríguez等[50]尝试在管式光生物反应器中使用一种新型静态混合器,该混合器能够以较低的能耗增加微藻细胞经历明暗循环的频率,进而实现高生产率。Cui等[51]设计出一种新型的切线双管式光生物反应器(TDTP),与同心双管式光生物反应器(CDTP)和传统管式光生物反应器(TTP)相比,微藻生物量产量分别提高了51.2%和124.8%,在微藻培养中具有良好的应用前景。Nagy等[52]建立了一个基于厌氧诱导的光自养产氢反应器,通过使用密集的薄层培养,使得产氢量增加至普通培养的3倍。
研究人员还将固定化技术与光生物反应器相结合用于产氢微藻的培养。固定化技术具有反应速度快、易于操作、易于收集细胞等优点,还可以保护细胞免受污染,进而延长衣藻的产氢时间[53-54]。
4 展望
绿藻光合产氢提供了利用水和太阳能这两种最丰富的资源去产生可再生资源的可能性,是一种前景与挑战并存的生物技术,在解决能源及环境危机问题中发挥着重要作用。在过去几十年里,国内外学者就如何提高微藻光合产氢效率这一问题展开了多角度的探讨,进行了大量的实验研究,提出并建立了丰富的产氢理论和实验方法,为绿藻产氢领域的未来发展奠定了坚实的基础。目前大部分的研究主要集中在提高氢化酶对氧的耐受性以及能量转换效率两个方面,前者通过基因工程的手段筛选出对氧耐受或光合放氧过程受到限制的高效产氢藻株;后者通过发掘光合作用高效吸能、传能和转能的分子调控机制,使更多的电子不断流向氢化酶。
尽管目前已经开创出许多提高绿藻产氢量的方法和手段,但距离理论预期值还有一定差距,绿藻产氢的水平仍不足以使氢气作为一种商业竞争的能源载体。未来实现绿藻光合产氢工业化应用的努力方向包括:
(1)发掘更多高产氢突变藻株。深入挖掘并研究微藻在厌氧条件下的代谢调控网络,利用分子生物学、合成生物学等交叉学科的技术手段快速筛选高效且稳定的产氢藻株。可以通过突变特定的参与光合作用的相关蛋白或氢化酶基因、改变其他代谢途径以及将不同高产氢的表型进行组合等方式提高绿藻光合产氢能力。另外可以通过高通量组学分析突变体藻株相较于野生型藻株的代谢变化,发现更多遗传改造的靶点。
(2)开发高效光生物反应器。实验室小规模培养过渡到工业大规模应用需要很多硬性要求和条件,并且后续氢气的收集、运输和储存都需要很大的成本。高效光生物反应器的开发是从基础研究转到应用研究的关键一步。目前开发的高效光生物反应器可以稳定并且精确控制产氢过程中的光照、温度等基础条件。未来高效光合反应器的设计将侧重于成本低、效能高、延长产氢时间以及氢气易收集等方面,早日形成绿藻工业化大规模培养体系。
绿藻产生的生物氢能否成为未来人类社会主要的能源供应,不仅取决于合成生物学和微生物基因工程的发展,还取决于其他高效制氢方法的成本效益,以及社会对氢能使用的接受程度。
猜你喜欢
安徽农业科学(2021年4期)2021-03-18
浙江农林大学学报(2019年4期)2019-07-24
上海师范大学学报·自然科学版(2018年6期)2019-01-14
深圳大学学报(理工版)(2018年5期)2018-09-26
中国真菌学杂志(2018年2期)2018-05-17
安徽农业科学(2018年18期)2018-05-14
无机盐工业(2017年5期)2017-05-25
郑州大学学报(理学版)(2017年1期)2017-04-07
化工管理(2017年25期)2017-03-05
中国真菌学杂志(2016年5期)2016-12-12
