
冀西北山地森林不同生境下白桦分枝结构特征
2024-02-28 14:01郑玉张化永邹恒超黄头生白卓青甄欣欣
科学技术与工程 2024年3期
郑玉, 张化永, 邹恒超, 黄头生, 白卓青, 甄欣欣
(华北电力大学工程生态学与非线性科学研究中心, 北京 102206)
分枝格局是描述树冠内部结构的重要指标,反映了树叶在树冠外围的分布情况。枝构件的三维形态和枝条的发散程度是分枝格局的重要内涵[1],直接影响了树木在空间环境的生存能力和拓展能力,以及树木光合作用、呼吸作用、营养运输等生理机制[2-3]。根据前人研究,一般采用枝条长度、枝条基径、分枝角度、枝径比、分枝率和分枝间距等指标度量分枝格局[4-5]。研究分枝格局能够揭晓树冠的动态变化、林木的生长对策[6-9]以及提供合理的林业管理方法[10]。
林木生长发育过程中形成的分枝格局是内部遗传基因和外部环境共同调控的,外部环境包括光照[11-12]、营养[13-14]、水分[15-16]、风[17-19]、空间[20]等因子,说明不同生境下多种因子的竞争关系制约着枝条的生长发育。白桦在不同的中生生境下花序构件数量随林分郁闭度的增大而减少[21],在优质的土壤环境中水分和氮含量充足对菖蒲枝条密度有正向影响[22]。近些年有许多学者对白桦分枝格局进行了广泛研究,主要探讨树冠分形维数[8]、分枝格局与种植密度的响应关系[23]、基因对分枝构型的调控[24]、树木生长过程中枝条的变化规律[25]以及枝条生物量和发芽的内部影响因素[26]等。陈东升等[27]、贾炜玮[28]在研究枝长、枝径生长关系时加入了树干胸径改进模型,说明胸径与分枝结构存在一定关系。然而,针对不同生境下种间种内竞争造成的白桦分枝格局差异研究还相对较少。本研究将植物构型理论应用于不同物种竞争生境下的白桦,旨在探讨白桦枝构件对环境的适应策略和进化机制,选择出适宜冀西北地区白桦生长的生境。
冀西北地区是生态敏感区,由于土地的不合理利用,生态退化比较严重[29]。白桦(BetulaplatyphyllaSuk.)是冀西北山地森林乡土树种,成熟植株平均树高达10 m左右,也是保障当地森林水源涵养的重要乔木之一。本研究对白桦纯林和不同混交林(混交林Ⅰ:白桦和落叶松;混交林Ⅱ:白桦和云杉;混交林Ⅲ:白桦、杏树、杨树和樟子松)下白桦分枝格局差异和枝长胸径间关系进行研究,以期为当地森林管理提供可靠的理论依据。
1 材料与方法
1.1 材料
研究区处于内蒙古高原与华北平原的汇接地带(从华北平原向内蒙古高原过渡的石质山地和丘陵盆地),位于京津冀西北区张家口永定河上游,地理坐标为40°57′N~41°00′ N、115°20′E~115°28′ E。研究区海拔在814~2 174 m,地势西北高,东南低。地处高寒地区,温带大陆性季风气候较明显,昼夜和季节温差大,土壤主要类型是棕壤、褐土和栗钙土,分布着二十世纪六七十年代栽植的人工林,以及遭受过度人为干扰的次生林。森林乔木资源丰富,其中次生林的优势树种主要为白桦(BetulaplatyphyllaSuk.)[30]。
2019年8—9月,在研究区域共设置了4个100 m×100 m的样地(表1),分别为白桦纯林、混交林Ⅰ(白桦81.5%,落叶松(Larixprincipis-rupprechtiiMayr)18.5%)、混交林Ⅱ(白桦34.4%,人工云杉(PiceaasperataMast.)64.1%)和混交林Ⅲ(白桦16.6%,杏树(PrunusarmeniacaL.)50.2%,杨树(PopulusdavidianaDode.)18.9%,樟子松(Pinussylvestrisvar.)8.7%)。在样地的对边中位线处建立两条相交的10 m×3 m的十字样带,选取生长良好、无病害的白桦各3株,重复测量,并对选定的植株进行编号。
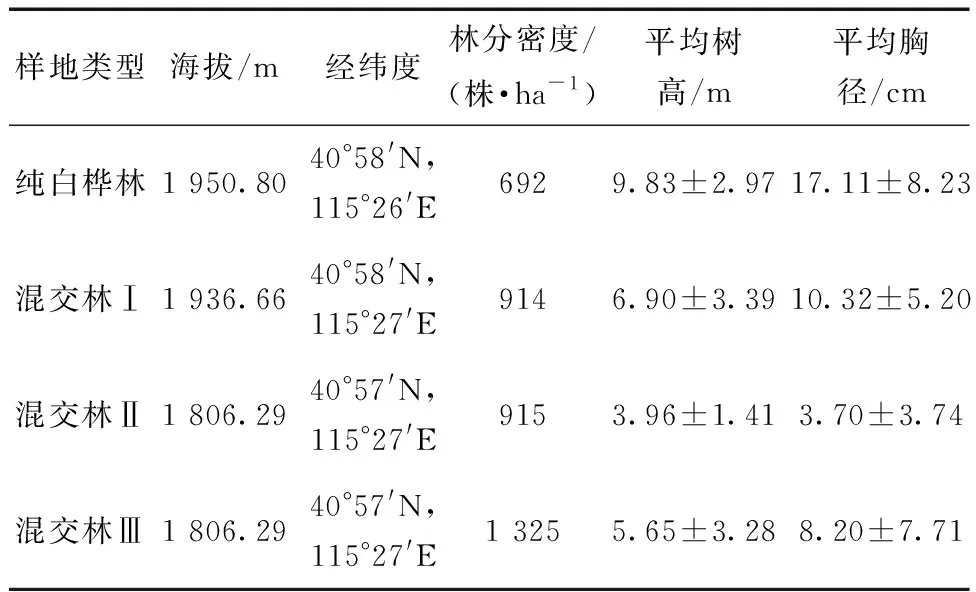
表1 样地基本参数
1.2 方法
1.2.1 枝级的确定方法
分枝级数的定义方法分为离心法和向心法。本研究选取与树冠发育一致的离心法,使相同功能的枝条处于相同的枝级,避免违背生物学原理[31]。由内向外、自下而上,依附主干生长的枝条作为1级枝,1级枝上的侧生枝为2级枝,2级枝上的侧生枝为3级枝,以此类推其他枝级,如图1所示。

图1 分枝解析图Fig.1 Branch diagram
1.2.2 指标的测定与计算
用游标卡尺测定第i级分枝与第i-1级分枝交汇处的D(基径);用钢卷尺(精度为0.1 cm)测定各级H(基部至顶端的总枝长);电子秤(精度为0.000 1 g)测定干重M(105 ℃温度下杀青,85 ℃温度下烘干至恒重);统计分枝数等指标。
其他分枝指标的计算方法如下。
枝长比(ratio of branch length,RBL):
RBLi∶i-1=Hi/Hi-1
(1)
式(1)中:Hi和Hi-1分别为第i和第i-1级枝条的长度。
枝径比(ratio of branch diameter,RBD):
RBDi∶i-1=Di/Di-1
(2)
式(2)中:Di和Di-1分别为第i和第i-1级枝条的基径。
逐级分枝率(stepwise bifurcation ratio,SBR):
SBRi∶i-1=Ni/Ni-1
(3)
式(3)中:Ni和Ni-1分别为第i和第i-1级枝条的分枝数。
分枝密度(branch fork density,BFD):
BFDi=Ni/Hi
(4)
式(4)中:Ni和Hi分别为第i级枝条的分枝数和枝长。
1.2.3 数据处理
试验数据采用Excel 2019进行初步整理,运用SPSS 23.0中的Kruskal-Wallis非参数检验对数据多重比较分析,使用Origin 2017绘图。样地乔木信息如表2所示。
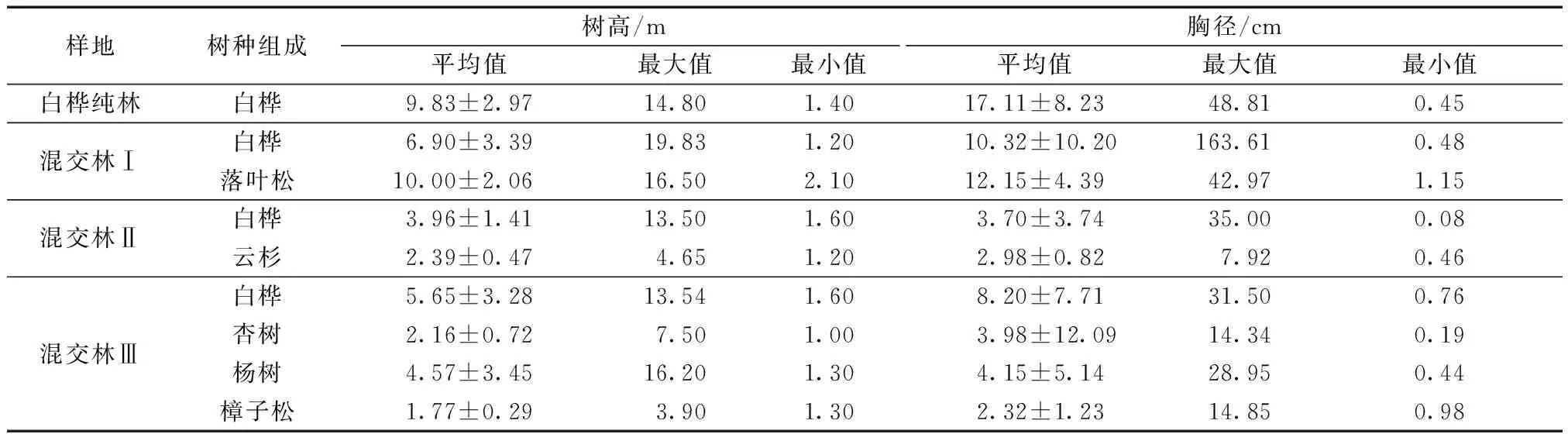
表2 样地乔木信息
2 结果与分析
2.1 各级分枝指标变异系数
如图2所示,不同分枝指标间变异系数存在一定差异。一级枝M、RBL、RBD和SBR的变异系数较大,分别为0.44、0.45、0.57和0.39,属于高度变异,说明一级枝条在各样点间不稳定;H、D和BFD的各级间变异系数相差不大,在0.09~0.16,属于弱变异。
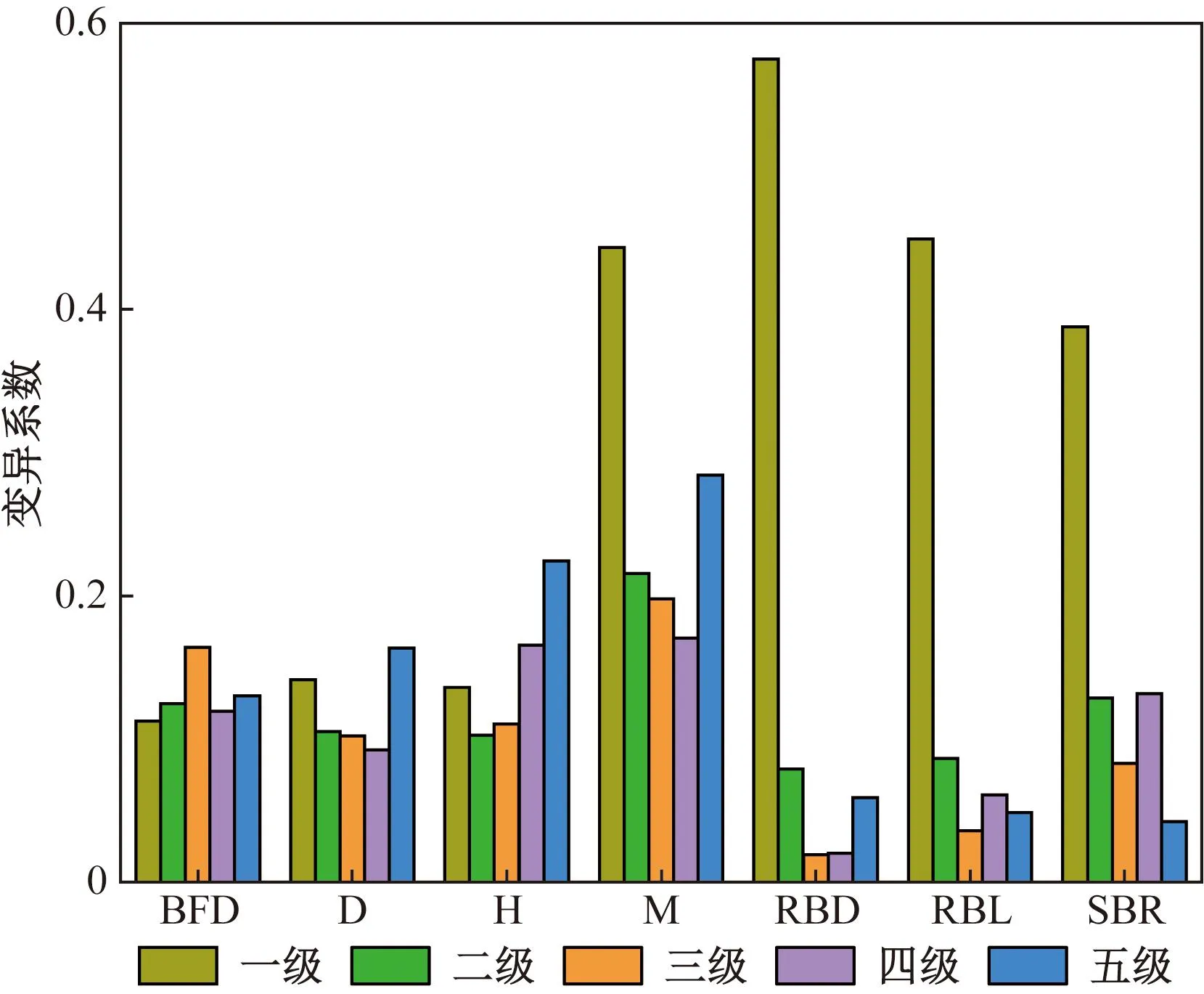
图2 分枝构型指标变异系数Fig.2 Variation coefficient of branching configuration index
2.2 分枝格局与树干胸径关系研究
分枝结构的差异是由多方面造成的,胸径(diameter at breast height,DBH)直接影响树的大小,进而影响树冠的内部结构——分枝结构,进一步分析枝长、枝径、枝生物量、枝长比、枝径比、逐级分枝率和分枝密度与胸径之间的关系。
如图3所示,各级枝长、枝径和生物量与白桦胸径均存在显著性相关,在0.426~0.880。枝级越大,分枝长度、基径和生物量越小。各级分枝长度、基径和生物量与胸径的线性图存在高度一致性,表明三者存在强相关性。各级枝条表现出相同的规律:指标大小随着白桦胸径的增大而增大。分枝长度受树干胸径的影响较大,尤其是一级枝条,树干胸径增加1个单位枝长平均增加5.04个单位;各级分枝基径随树干胸径增加1个单位分别增加0.037、0.014、0.011、0.006、0.005个单位,基径受树干胸径的影响逐级减小;各级分枝生物量随树干胸径增加1 mm分别增加0.414、0.025、0.007、0.003、0.002 g,受树干胸径的影响逐级减小。拟合枝长比、枝径比、逐级分枝率和分枝密度与胸径的关系发现它们均不存在显著性关系(p>0.05)。
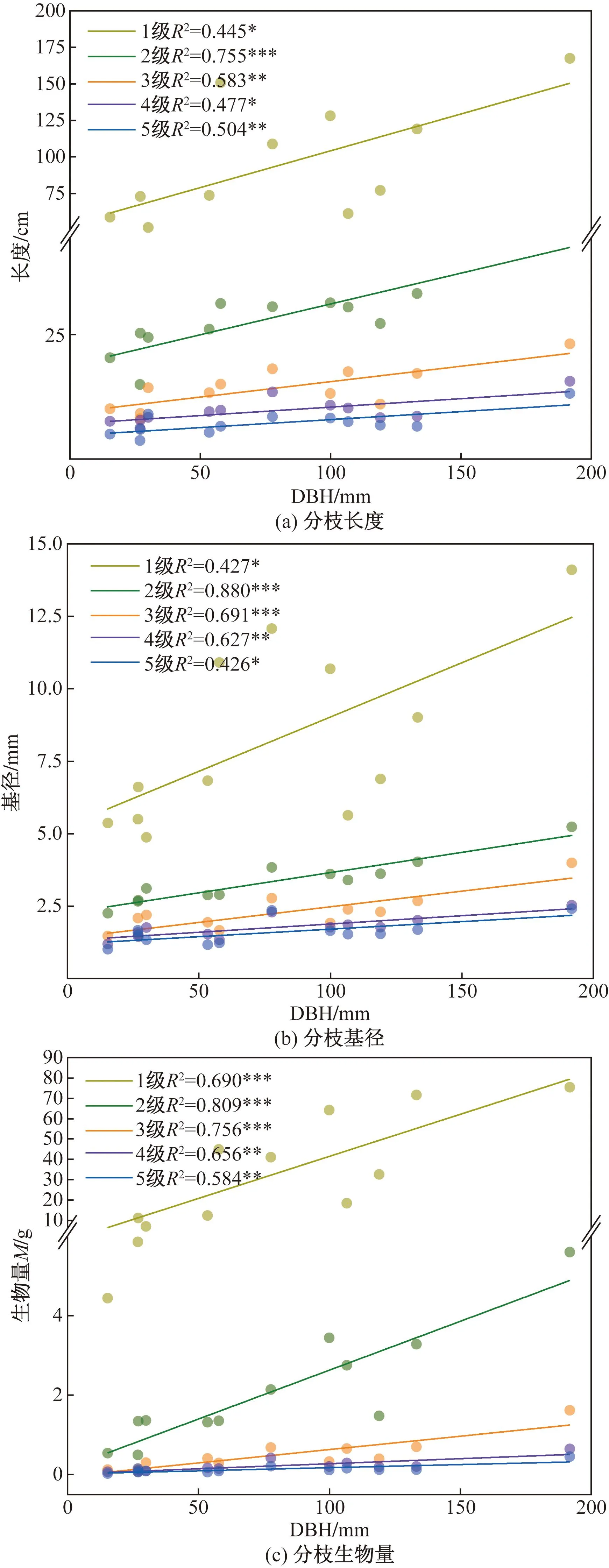
*p< 0.05 为有统计学差异;**p<0.01 为有显著统计学差异;***p<0.001为有极其显著的统计学差异图3 枝长、枝径和生物量与胸径DBH的关系Fig.3 Relationship of branch length, diameter and biomass with DBH

不同字母表示白桦同一分枝等级下枝长、枝径和枝生物量在不同生境下差异显著性(p <0.05)图4 不同生境下白桦枝长、枝径、枝生物量差异比较(中位数)Fig.4 Comparison of birch branch length, diameter and biomass in different habitats (median)
2.3 不同生境下白桦分枝结构差异
分枝长度、分枝基径、分枝生物量等指标能够反映植物在空间中的拓展能力和生产能力。不同生境存在的树种不同,种间对空间光照和土壤营养的竞争,可能会带来分枝结构的差异,这种差异不仅仅和遗传基因有关,和环境因素也存在着微妙的关系。
2.3.1 枝长、枝径和生物量
枝条长度、生物量和基径是通过直观测量得到的指标,用于描述枝条的生长发育状况[32]。如图4所示,在4种生境中明显呈现分枝长度、基径、生物量随分枝等级上升而减小。由Kruskal-Wallis检验(表3)得,不同生境之间同等级枝条长度、生物量和基径均存在极显著性差异(p<0.001)。对于不同的分枝等级,4种生境间也存在着不同的差异。一级枝的3个枝长、枝径和生物量仅混交林Ⅰ和混交林Ⅱ间存在显著性差异,且混交林Ⅰ>混交林Ⅱ;混交林Ⅱ与其他3种生境在2~3级枝的H、M和D上均存在显著性差异,混交林Ⅱ的中位数最小,H、M和D分别为17.5、0.42、2.34(2级),9.99、0.14、1.67(3级);除了白桦纯林与混交林Ⅲ,其他生境两两间4~5级枝的M和D均存在显著性差异。

表3 不同生境下白桦枝长、枝径和枝生物量Kruskal-Wallis检验
2.3.2 枝长比、枝径比和逐级分枝率
相邻级枝条的长度、基径和分枝数的比值描述了上一级枝对下一级枝的承载力以及影响程度[33-34]。在所有生境中,随着分枝等级的增大,RBLi∶i-1、RBDi∶i-1、SBRi∶i-1有不同程度的增长趋势(图5)。除了第5级枝的枝长比,其他生境间的RBL、RBD和SBR均存在极显著性差异(p<0.001)。两两对比发现,混交林Ⅰ与其他3种生境在RBL1∶0、RBL2∶1、RBL3∶2上均存在显著性差异,混交林Ⅰ为最小值,分别为0.1、0.13和0.2;混交林Ⅰ与混交林Ⅱ在5个RBD上均存在显著性差异,1~3级枝上混交林Ⅰ的枝径比显著小于混交林Ⅱ,而4~5级枝上混交林Ⅱ的枝径比显著小于混交林Ⅰ;混交林Ⅲ在SBR1∶0显著高于其他生境,值大小为0.55;SBR2∶1、SBR3∶2在不同生境间存在显著差异,两者的中位数大小排序相同:白桦纯林>混交林Ⅲ>混交林Ⅰ=混交林Ⅱ,如表4所示。
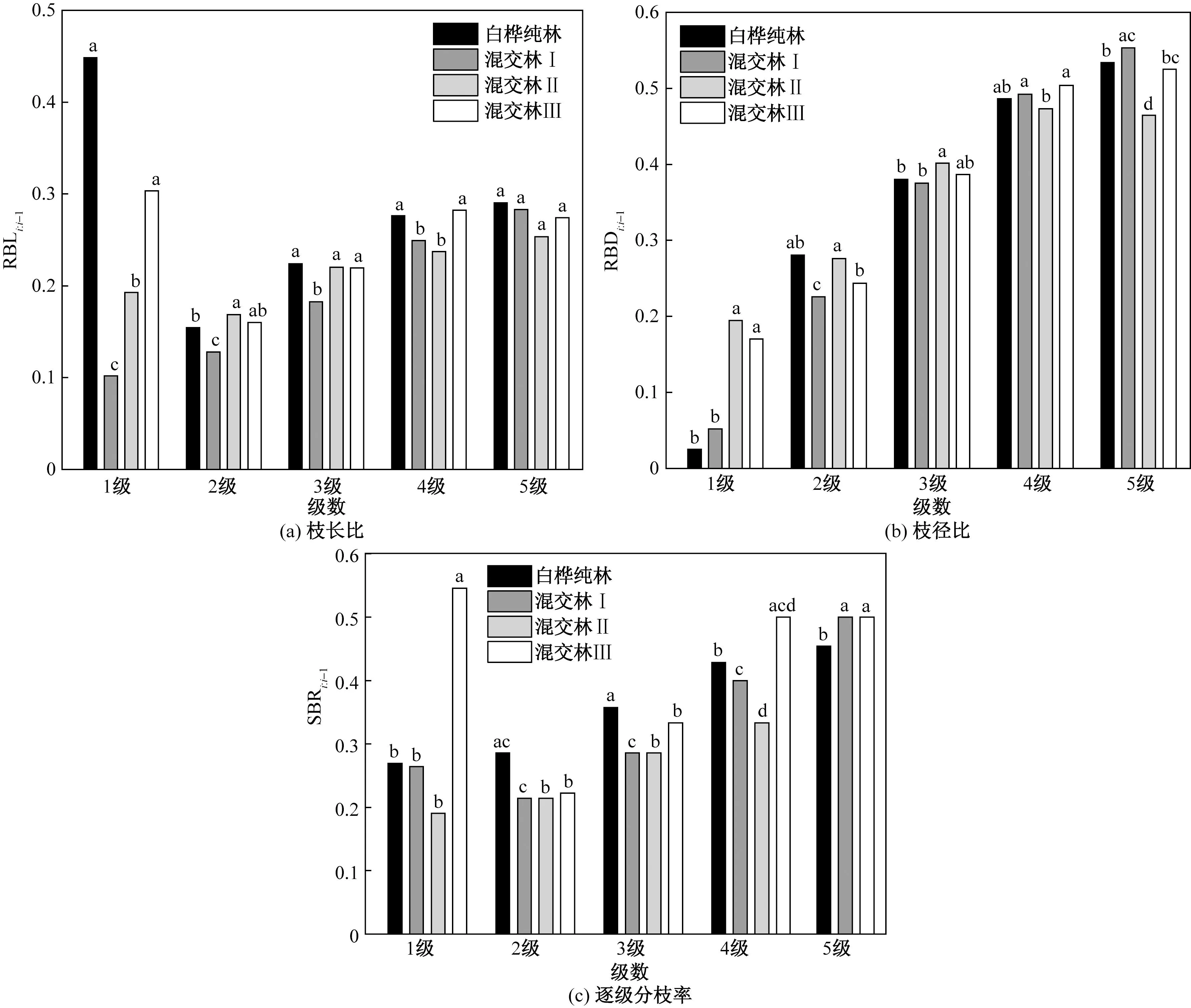
不同字母表示白桦同一分枝等级下枝长、枝径和枝生物量在不同生境下差异显著性(p <0.05)图5 不同生境下白桦枝长比、枝径比和逐级分枝率差异(中位数)Fig.5 Comparison of RBL, RBD and SBR in different habitats (median)
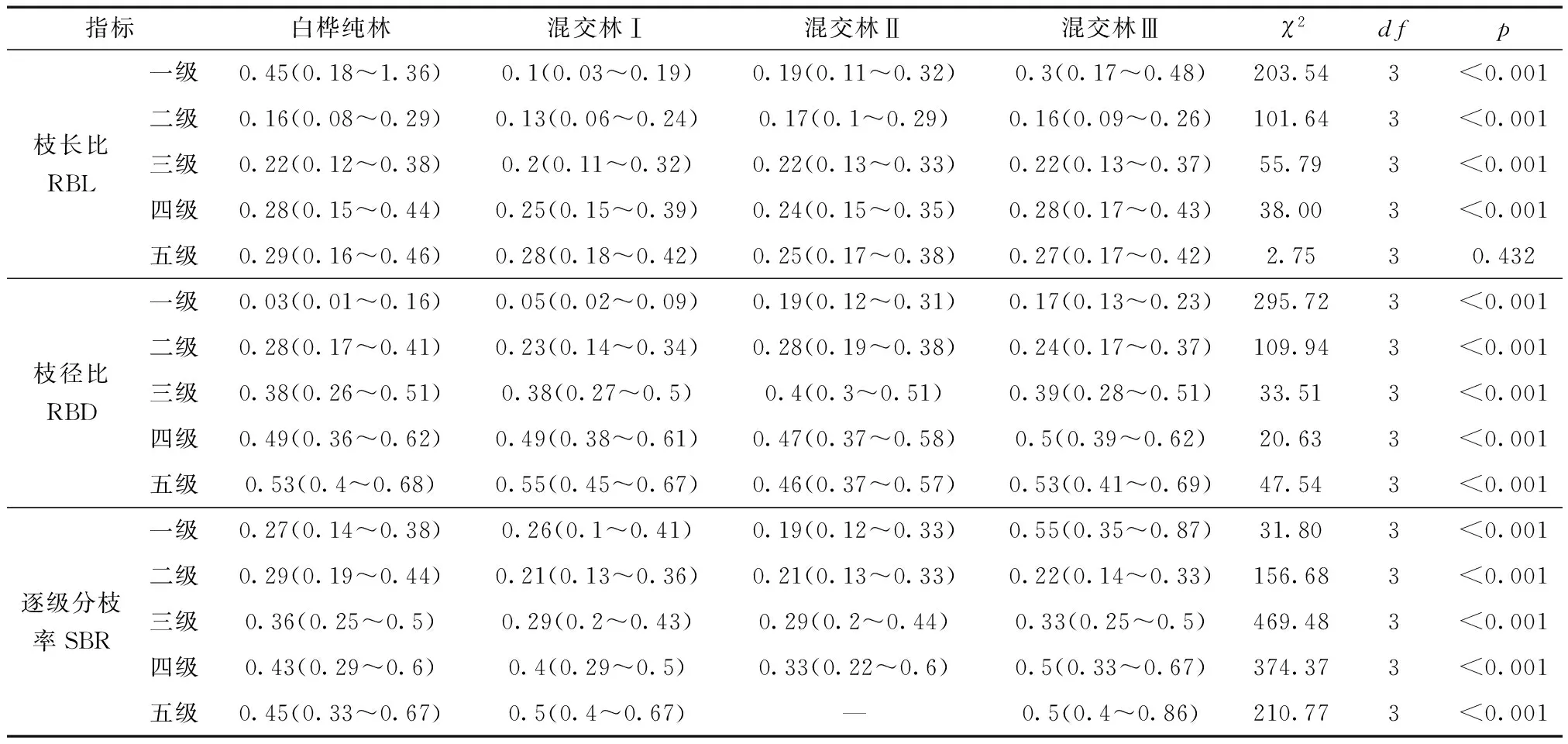
表4 不同生境下白桦枝长比、枝径比和逐级分枝率Kruskal-Wallis检验Table 4 Kruskal-Wallis test of RBL, RBD and SBR in different habitats
2.3.3 各级分枝密度
分枝密度BFDi用来表示在植物枝条附近空间中任一点发现枝条的可能性[35]。根据Kruskal-Wallis H检验(如表5所示)得:对比4种生境下白桦的BFD发现,混交林ⅡBFD1显著高于其他生境,中位数为0.14,同时混交林Ⅱ与白桦纯林和混交林Ⅰ的BFD2、BFD3、BFD4均存在显著性差异,混交林Ⅱ为三者的最小值,分别为0.12、0.12、0.13。除了混交林Ⅰ与混交林Ⅱ、Ⅲ,其余生境在BFD2、BFD3、BFD4和BFD5均存在显著性差异,如图6所示。
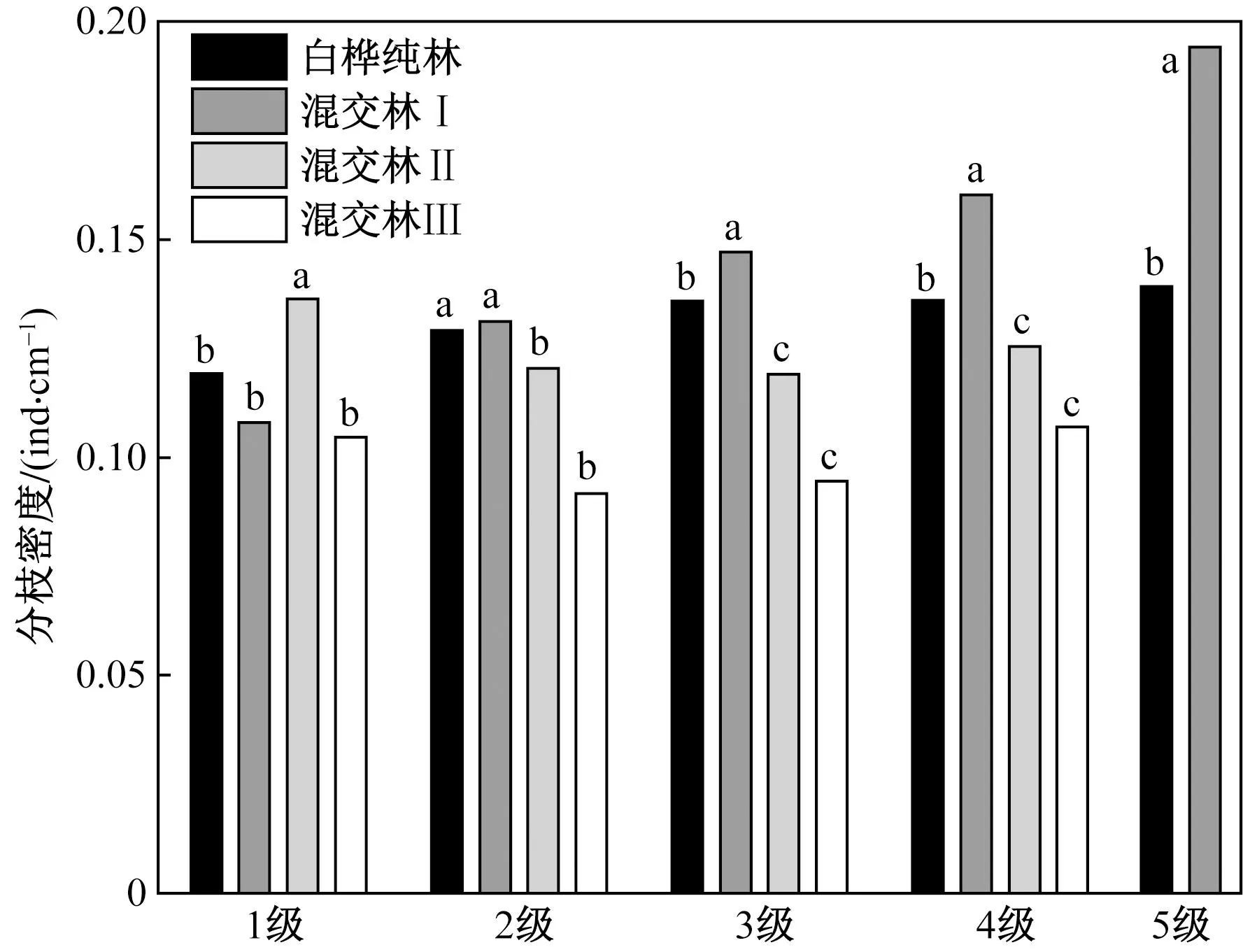
不同字母表示白桦同一分枝等级下枝长、枝径和枝生物量在不同生境下差异显著性(p <0.05)图6 不同生境下白桦分枝密度差异(中位数)Fig.6 Comparison of BFD in different habitats (median)

表5 不同生境下白桦分枝密度Kruskal-Wallis检验
3 讨论
植物在同一地区不同生境下,受到不同光照、温度、水分、营养元素和竞争物种等因素的影响,从而形成不同的分枝格局[36-38]。单个树枝的长度、基径、生物量代表着枝条的大小[39-40],枝长比、枝径比和逐级分枝率代表着枝条生长过程中的变化[15, 34],分枝密度代表枝条分布的密集程度[35],这些属性决定树冠结构,值的大小决定树木的生长情况[41-42],也是树木可视化模拟系统中的重要形态参数[43]。
本研究分析了冀西北山地白桦树干胸径和不同物种竞争生境下枝构件差异,发现:在同一树种基因相同的情况下,树干胸径对分枝结构的影响较小,而在不同物种生存的生境下分枝结构存在显著差异,表明生境会对分枝结构产生影响。
树干胸径与白桦分枝长度、基径和生物量存在显著相关性,R2在0.426~0.880,这与前人研究建立枝长枝径模型时将树干胸径作为协变量引入模型有相似之处[27, 44],均考虑了胸径对分枝结构的影响。本研究将胸径对枝长、枝径和枝生物量的影响进行了量化,每增加1 mm胸径三者分别增加0.032~0.504 cm、0.005~0.037 mm、0.002~0.414 g,表明胸径是分枝构型的重要影响因素之一,并非主要影响因素。
在混交林Ⅰ下的白桦,相对其他生境,1级和2级的H、M和D最大,而混交林Ⅱ各级枝条的H、M和D均较小,这可能是该生境中白桦和人工种植的云杉都为较小幼苗,生长发育阶段对资源的竞争激烈。研究区域生态退化比较严重,混交林Ⅱ分布植被少,土壤涵养水分少,不利于枝条的伸长[15, 45-47]。因此白桦生长慢个体小,平均高度、胸径分别为3.96、3.70 cm。从2级开始至5级,白桦纯林的枝长在4种生境中相对较长,白桦纯林不存在竞争树种,且林分密度最小(692 株/ha),拥有充足的生长空间。不论何级分枝,白桦均表现出分枝长、生物量和基径与胸径呈正相关(如图2所示),这与张锐的研究结论一致[48]。各级分枝基径、生物量与分枝长有近似的排列规律,例如一级枝的枝长和基径中位数顺序为混交林Ⅰ>混交林Ⅲ>白桦纯林>混交林Ⅱ,表明三者存在较强的相关性。
不同生境下各级RBL呈现相同的规律:白桦纯林>混交林Ⅲ>混交林Ⅱ>混交林Ⅰ。一般枝长比与分枝密度呈反比,由于分枝数量较多时,上一级枝条的营养物质输送被分散,枝条的生长受到限制。在图5(a)和图6的2~4级分枝中该规律较为明显。混交林Ⅰ与混交林Ⅲ中白桦在不同竞争生境下表现出不同的生长策略,混交林Ⅰ分枝密集,而混交林Ⅲ枝条较长[49]。研究发现,所有生境下的RBD均随枝级增大而增大。混交林Ⅰ在RBD1∶0和RBD2∶1均较小,而RBD4∶3和RBD5∶4较其他生境最大,表明混交林Ⅰ在3~5级枝生长旺盛。除了混交林Ⅲ,SBR在其他生境下均随枝级的增大而升高。混交林Ⅲ的SBR1∶0显著高于其他生境。经实地调查发现,该样地的植被丰富度最高,有白桦、杨树、落叶松、杨树以及伴生的榛子树、金露梅、山丁子等小乔木,且丰富度高的群落土壤营养物质和微生物也高[50-51],更利于枝条生长,同时提高一级枝的逐级分枝率可以优先占据上层空间的能源利用。
前期研究表明,分枝率的研究结论具有争议性,认为植物内部分枝率具有稳定性,而外部条件又能带来差异性[52-53],这说明了植物的可分离性和自相似性[35]。逐级分枝率代表着相邻枝级间枝条的分配关系。在一定范围内逐级分枝率是稳定的,外界人为对植物较大程度的干扰,比如截枝、砍伐,会提高植物的逐级分枝率,解除顶端优势生长[54]。本研究表明,对于同一地区的同一物种,逐级分枝率与胸径不存在显著相关性,而在不同生境下存在显著性差异,证实了分枝率的稳定性与差异性的对立统一,分枝率差异可能与种内种间竞争、气候、养分、水分等因素有关。
在不同生境条件下,在混交林Ⅰ中,随着分枝等级的增加,分枝密度在4种生境中逐渐突出。为与共存的落叶松竞争光、水和热等资源,白桦选择提高分枝密度的生存策略[55],最大可能地弥补因竞争而带来的物质积累和能量代谢方面的损失,通过分枝格局的调整来平衡对资源的竞争与利用。
4 结论
树干胸径和4种生境对白桦分枝结构均存在显著性影响。白桦分枝长度、基径和枝生物量随树干胸径显著线性增长(p<0.05),树干胸径的影响程度随1~5级分枝逐级减小。混交林Ⅰ下白桦的分枝基径、生物量、分枝长以及分枝密度均优于白桦纯林、混交林Ⅱ和混交林Ⅲ,能在竞争中获取更多的空间资源,其分枝结构具有一定的优势性。本研究量化分析了树干胸径对分枝结构的影响,揭示了树干胸径在建立分枝结构模型中的重要性。本研究关于环境对植物分枝格局的影响仅选择了不同物种所组成的不同生境,而各种环境因素(如气候、地形和土壤等)对分枝格局的影响程度尚不清晰,是进一步研究的重要方向。
猜你喜欢
今日中国·西班牙文版(2021年4期)2021-04-09
文苑(2019年20期)2019-11-20
文苑·经典美文(2019年10期)2019-10-16
四川林业科技(2019年5期)2019-07-07
安徽农学通报(2019年1期)2019-02-14
湖北林业科技(2019年6期)2019-01-06
现代园艺(2018年2期)2018-03-15
湖北林业科技(2017年2期)2017-05-25
华人时刊(2016年16期)2016-04-05
广西林业科学(2016年1期)2016-03-20
