
SYVN1介导FASN泛素化降解促进骨肉瘤细胞失巢凋亡
2024-03-12 08:54钟南山刘志礼
实用癌症杂志 2024年3期
钟南山 杨 枫 刘志礼
FASN(fatty acid synthase,脂肪酸合成酶)在多种肿瘤中高表达,促进肿瘤恶性表型[1]。我们前期研究也发现FASN通过介导细胞失巢凋亡抵抗促进骨肉瘤细胞增殖转移[2],但是FASN在骨肉瘤细胞中的表达水平调控机制有待于进一步阐明。泛素化修饰是蛋白翻译后修饰的重要方式之一,蛋白泛素化异常是肿瘤发生发展的重要因素[3]。在本研究中,作者通过构建失巢细胞模型,探讨了FASN受到泛素化修饰调控的机制,并鉴定出了SYVN1是其E3连接酶,过表达SYVN1促进细胞失巢凋亡抵抗而抑制骨肉瘤转移。
1 材料与方法
1.1 人骨肉瘤细胞培养
人骨肉瘤143B细胞系购自美国典型生物资源保藏中心(American type culture collection,ATCC)的细胞库。该细胞系通过短串联重复(STR)分析进行鉴定,并在MEM-α培养基(Gibco,美国)中培养。细胞补充10%胎牛血清(Gibco,美国)和100 U/mL青霉素-链霉素(Gibco,美国),并在37 ℃和5%二氧化碳环境下培养。
1.2 超低吸附培养皿法构建骨肉瘤细胞失巢凋亡抵抗模型
使用超低吸附培养皿(Corning,美国)培养细胞,将消化后的骨肉瘤细胞重悬后铺入超低吸附培养皿中,加入含20%FBS的MEM-α培养基进行培养,48 h后将含细胞的悬液进行收集,低速(200 rpm)离心,小心吸去上清,加入PBS洗涤细胞一次,再次低速离心去除上清剩余的细胞加入新鲜含20% FBS的MEM-α培养基继续培养,待“失巢凋亡抵抗”细胞生长球体直径至大于约30个细胞时,重复上述收集离心洗涤步骤,洗涤后的失巢凋亡抵抗细胞使用Accumax(Sigma,美国)解离液37 ℃孵育酶解10 min,解离成单个细胞后按1∶2~1∶3的比例再次种入超低吸附培养皿中。
1.3 慢病毒载体的构建
根据目标分子对应CDS的序列,在5'端和3'端引入酶切位点,合成全长的目标分子模板,连接至经双酶切的pLvTHM载体,转化DH5α大肠杆菌并挑取重组阳性克隆,经PCR、双酶切鉴定,鉴定正确的质粒送DNA测序。插入shRNA序列或转录本编码:shSYVN1:5′-GAGACAGTTTCAGATGATT-3′SYVN1:NM_032431.3,FASN:NM_004104.5。
慢病毒的包装与扩增:将重组的慢病毒载体,转染HEK293细胞,以产生具有感染能力的重组慢病毒。获得的重组慢病毒反复感染HEK293细胞以扩增病毒,采用终点稀释分析方法测定慢病毒滴度。
1.4 qRT-PCR法检测相关基因mRNA的表达
使用Trizol(Thermo,美国)提取总RNA并定量后,用逆转录试剂盒(诺维赞,南京)逆转录为cDNA,加入目的基因、上下游引物、含SYBR greenⅠ扩增酶(诺维赞,南京)等进行荧光定量PCR检测,以ACTB为内参。具体方法参见《分子克隆实验指南(第四版)》和有关试剂盒说明书。引物序列:FASN-F,5′-TGCTAGCTGATCGATCGATCGTCG-3′;FASN-R,5′-CGTAGCTGATCGATGCTAGCTAGC-3′;SYVN1-F,5′-AACCCCTGGGACAACAAGG-3′;SYVN1-R,5′-GCGAGACATGATGGCATCTG-3′;ACTB-F,5′-CATGTACGTTGCTATCCAGGC-3′;ACTB-R,5′-CTCCTTAATGTCACGCACGAT-3′。
1.5 流式细胞术
应用凋亡检测试剂盒(贝博,苏州)来检测骨肉瘤细胞的细胞周期分布和凋亡细胞的百分比。所有细胞系都使用不含EDTA的胰蛋白酶进行消化,并使用Annexin V/PI进行染色,经安捷伦流式细胞仪,检测APC-PEtexRED通道上细胞凋亡比例分布。
1.6 免疫印迹分析
使用RIPA裂解法(Thermo,美国)提取骨肉瘤细胞的总蛋白。蛋白质浓度由BCA检测法(Thermo,美国)确定。等量的蛋白质经SDS-PAGE电泳后通过湿转法转移到经甲醇活化的PVDF印迹膜上(Millipore,美国)。膜在室温下用5%脱脂牛奶-TBST封闭2小时,然后根据上样情况用FASN、SYVN1、UB、FLAG、HA等(CST,美国)一抗在4 ℃下孵育过夜。次日TBST清洗膜3次,用适当的辣根过氧化物酶(HRP)结合的二抗(抗兔和抗鼠,CST,美国)孵育1h,再次用TBST清洗膜3次。通过ECL成像(Bio-Rad,美国)检测免疫复合物在膜上的情况。
1.7 免疫共沉淀实验
收集细胞,IP buffer(Thermo,美国)裂解后4 ℃离心机15000 g离心15 min,收集上清,少量全细胞裂解液留于Western blot分析,其余加入目的蛋白/标签抗体和50 μl洗涤过的protein A/G-beads(MCE,美国)4 ℃孵育过夜,离心后去上清,取约30 μl 1x loading buffer 95 ℃加热磁珠10 min,产物用于Western blot检测目的蛋白及与其结合的其他蛋白。
1.8 裸鼠原位异种移植骨肉瘤肺转移模型
选取4周龄的雌性BALB/C-nude鼠(杭州子源实验动物科技有限公司,SCXK(浙),2019-0004,中国杭州)。首先建立AR-143B细胞裸鼠皮下荷瘤模型:将约1×107经不同处理方式处理的AR-143B细胞分别皮下注射至裸鼠左和右侧肩胛处,3周成瘤后脱颈法牺牲裸鼠,解剖取出瘤体,并分别将其切割约1 mm3大小的瘤块,植入裸鼠胫骨近端骨髓腔内;4周后脱颈法牺牲裸鼠,荧光活体成像仪检测原位成瘤情况及肺部转移情况,解剖取出瘤体和肺组织,测量瘤体体积V=长×宽×高×π/6,大体计数肺部转移灶数量。本实验方案得到了南昌大学第一附属医院伦理委员会的批准(江西,中国;编号:CDYFY-IACUC-202211QR012)。
1.9 统计学分析
离散数据采用非参数的Wilcoxon排名和检验分析。所有连续数据以均值±标准差表示。在双样本分析中使用t检验,在多样本分析中使用单因素方差分析。所有的分析都是用SPSS统计软件22.0版(SPSS,美国)进行的。
2 结果
2.1 骨肉瘤143B细胞悬浮状态下FASN泛素化水平降低而表达升高
Western Blot检测显示:在贴壁143B细胞中FASN蛋白水平显著低于AR-143B细胞中FASN表达水平(图1A),但是mRNA水平并无明显差异(图1B);提示失巢凋亡抵抗的骨肉瘤细胞中FASN表达的升高,可能与其转录后修饰相关。
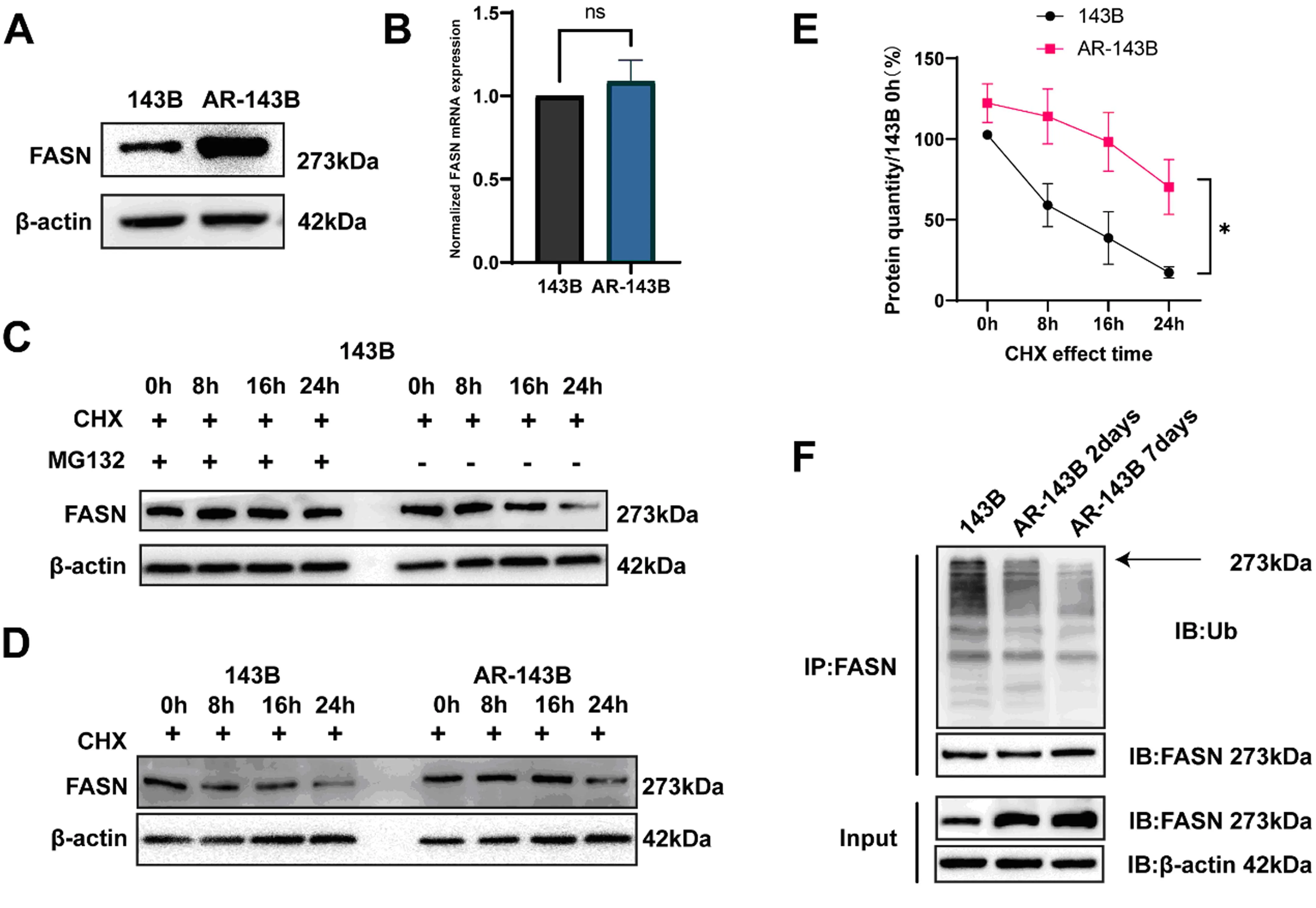
注:A&B为AR-143B与143B细胞中FASN的蛋白和mRNA表达情况;C为降解实验检测FASN在143B细胞内降解模式;D&E为降解实验检测143B与AR-143B细胞中FASN蛋白的半衰期;F为泛素化实验检测失巢凋亡抵抗过程中FASN表达量及其泛素化修饰水平的变化。
CHX-MG132降解实验显示,FASN的降解受蛋白酶体抑制剂MG132的抑制(图1C),并且相比贴壁143B细胞,AR-143B细胞中FASN蛋白降解速度有所下降(图1D、E);进一步的泛素化实验显示:AR-143B细胞中的FASN蛋白具有更低的泛素化水平(图1F),这提示AR-143B细胞中FASN水平的升高与FASN蛋白的泛素化降解水平降低有关。
2.2 FASN相互作用的E3-连接酶
通过蛋白组学测定了在143B细胞与AR-143B中具有差异的蛋白,通过功能富集把其中泛素化功能相关蛋白富集,其中AR-143B样品中表达水平降低的泛素化相关蛋白有12个,升高的泛素化相关蛋白有3个(图2A);与在线泛素化相关蛋白预测网站Ubibrowser(http://ubibrowser.bio-it.cn/ubibrowser/home/index)上FASN可能存在的E3泛素连接酶或去泛素化酶(图2B)进行交集,结果显示:E3连接酶SYVN1是其交集(图2C);Western Blot实验证实SYVN1在AR-143B细胞中相比普通143B细胞表达更低(图2D),并且免疫共沉淀实验显示:在HEK-293T细胞和AR-143B细胞中FASN和SYVN1这两个分子可以相互富集(图2E、F);这提示SYVN1很可能是FASN的E3连接酶。
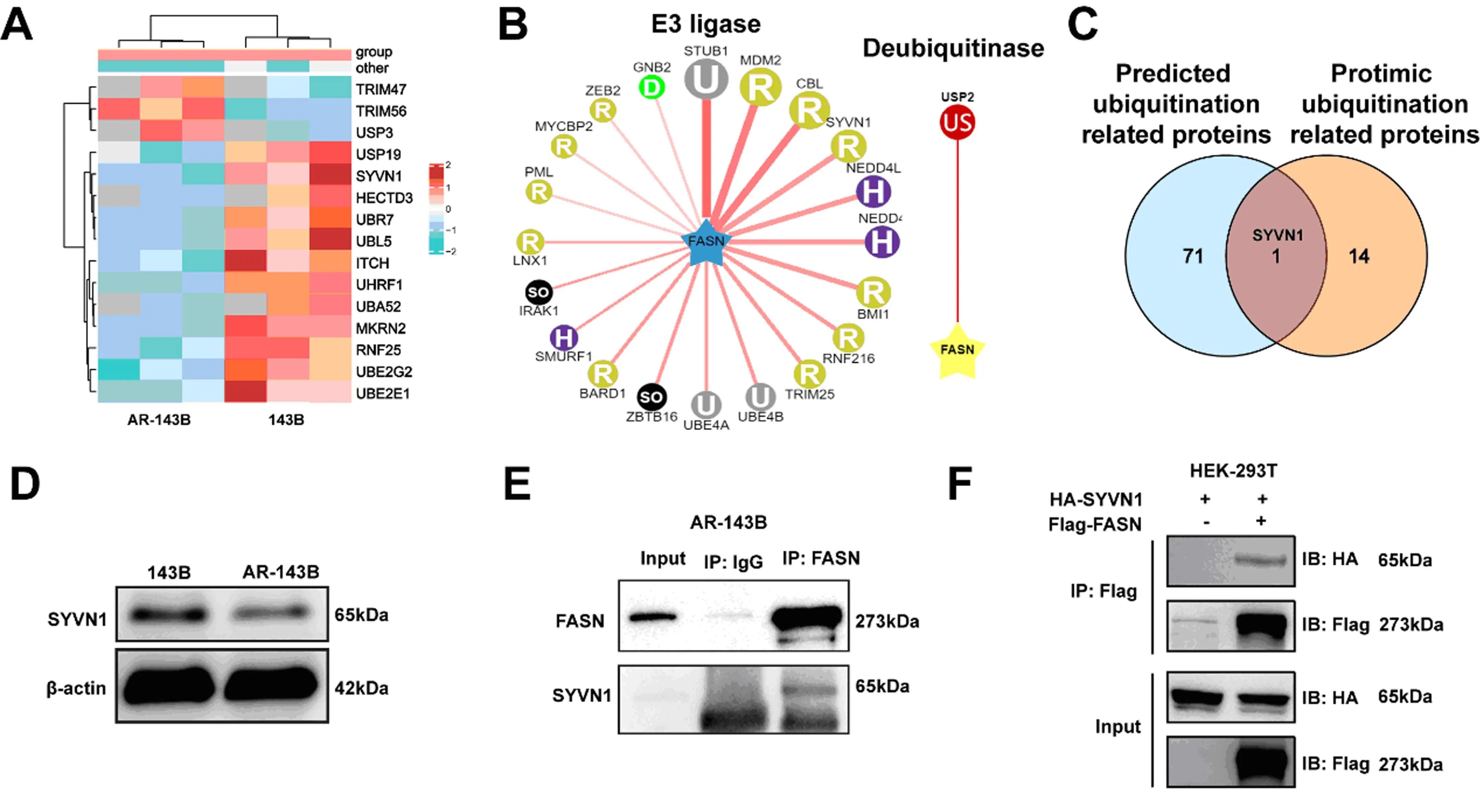
注:A为热图显示iTRAQ蛋白组学中143B与AR-143B细胞的泛素化相关蛋白水平差异;B为Ubibrowser预测FASN的E3泛素连接酶和去泛素化酶;C为蛋白组学结果和在线预测结果交叠;D为AR-143B与143B细胞中SYVN1蛋白水平验证;E&F为免疫共沉淀检测FASN和SYVN1间相互作用。
2.3 SYVN1促进FASN泛素化降解
在AR-143B细胞中通过慢病毒载体过表达或沉默SYVN1,qRT-PCR、Western blot验证其过表达或沉默效果,结果提示,经筛选的稳定细胞株中SYVN1的水平都被成功上调与下调(图3A、B),过表达SYVN1后FASN的泛素化水平升高,并且蛋白水平降低(图3C);沉默SYVN1后,FASN的泛素化水平随之下降而蛋白水平升高(图3D)。以上结果提示SYVN1可作为FASN的E3连接酶通过泛素化修饰FASN而影响其稳定性。
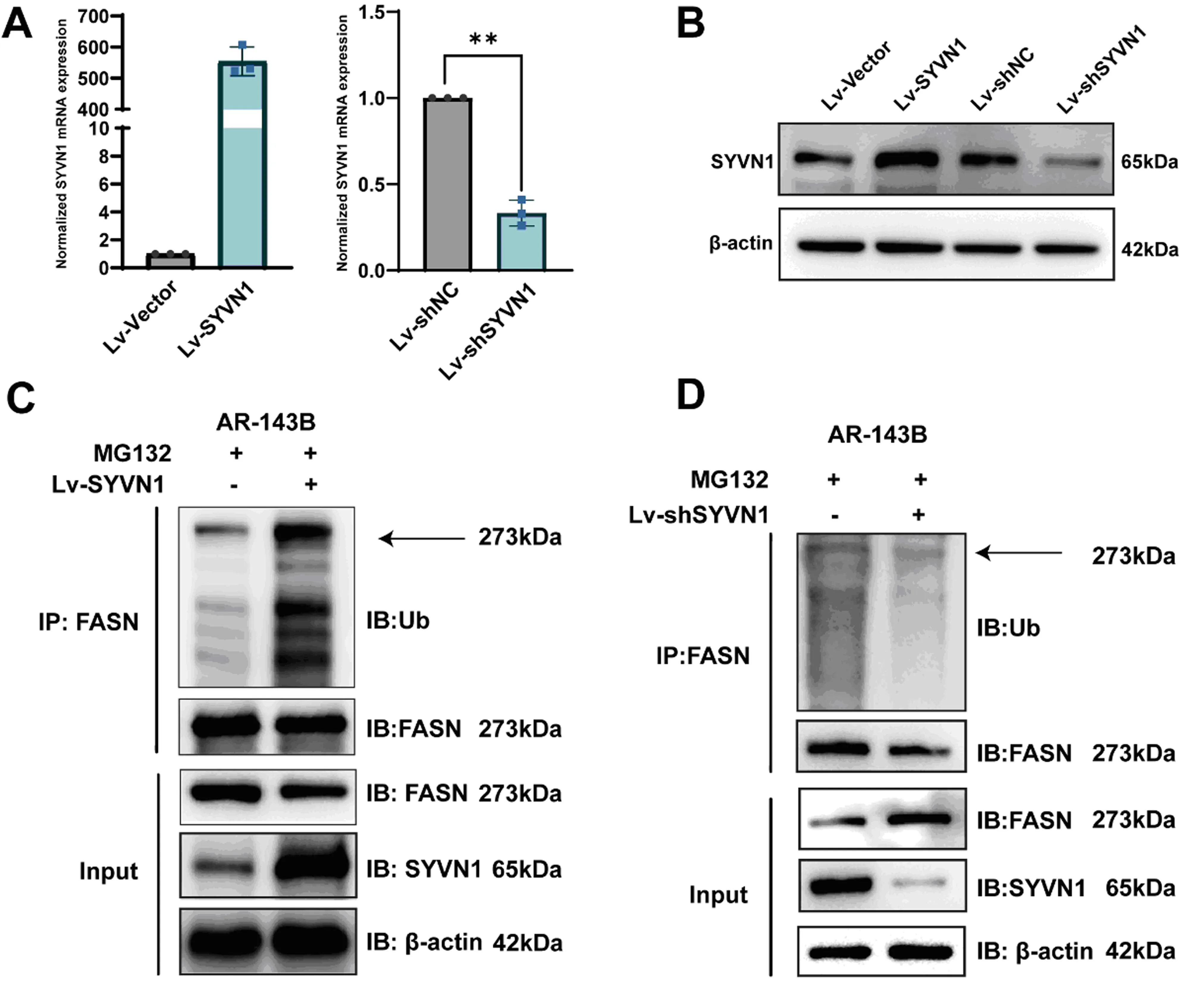
注:A与B为Western blot和qRT-PCR验证过表达或沉默SYVN1效果;C为过表达SYVN1可显著增加FASN泛素化水平使其降解;D为沉默SYVN1可显著降低FASN泛素化水平。
2.4 SYVN1降解FASN促进骨肉瘤143B细胞失巢凋亡
为分析SYVN1对骨肉瘤细胞失巢凋亡抵抗能力的影响是否通过降解FASN实现,使用FASN标签质粒转染过表达SYVN1的稳定细胞系进行回复实验。Western blot结果提示,在上调SYVN1表达的稳定细胞株中FASN的量有所下降,转染1 μg FASN质粒后FASN表达有所上升(图4A)。肿瘤成球实验检测细胞增殖活力,流式细胞仪检测细胞凋亡情况,结果显示:相对于阴性对照组,Lv-SYVN1转染的AR-143B细胞中,细胞失巢凋亡率显著升高,转染FASN质粒的细胞中凋亡细胞比例明显下降,而同时上调SYVN1与FASN,相较于只上调SYVN1的组别,其凋亡细胞比例有所下降(图4B);肿瘤成球实验显示,AR-143B细胞上调SYVN1水平后肿瘤球直径显著小于阴性对照组,而上调FASN则会增强细胞成球能力,同时上调SYVN1与FASN,相较于只上调SYVN1的组别,其细胞成球能力有所提升(图4C)。以上结果提示SYVN1通过介导FASN泛素化降解,促进143B细胞失巢凋亡。
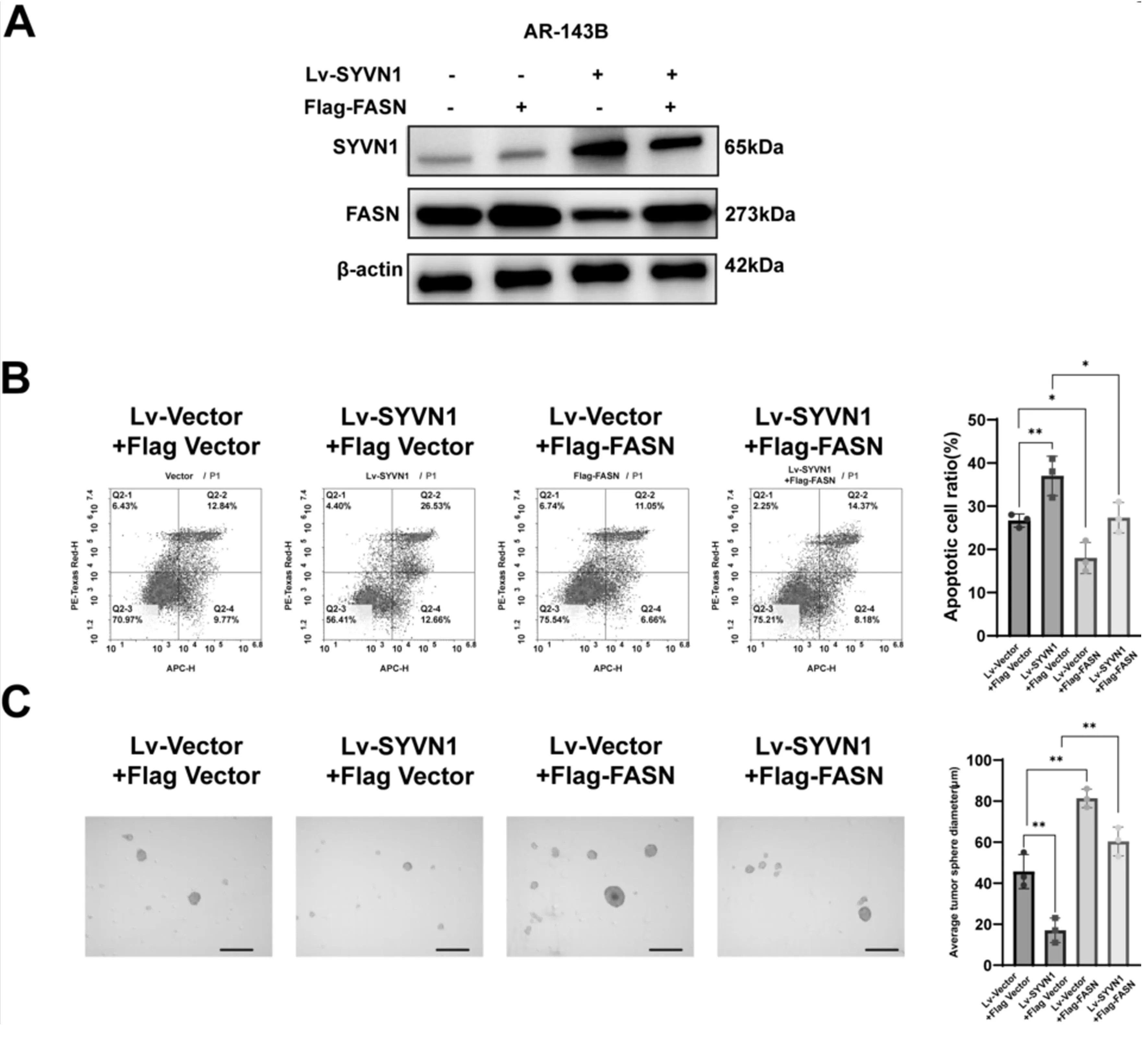
注:A与B为Western blot检测;C为流式细胞术检测;D为肿瘤成球实验检测。
2.5 SYVN1抑制骨肉瘤细胞异种原位成瘤模型肺部转移
将约1×107分别经Lv-SYVN1和Lv-Vector转染(带GFP荧光蛋白标签)的AR-143B细胞皮下注射裸鼠左右两侧肩胛处(n=1),3周后两组细胞均皮下成瘤,颈椎脱臼法处死裸鼠,解剖取出瘤体并切割成小块原位移植至裸鼠胫骨近端髓腔,建立裸鼠骨肉瘤原位模型(n=6)。4周后荧光活体成像观察成瘤情况,发现经Lv-SYVN1感染细胞组3只裸鼠未见瘤体形成(成瘤率50%),经Lv-Vector感染细胞组裸鼠均成瘤(成瘤率100%)(图5A),但肺部未见显著转移性结节的荧光信号;处死裸鼠取出瘤体,测量计算体积,大体观察肺部微小转移结节数,结果显示:Lv-SYVN1感染细胞组瘤体体积显著小于经Lv-Vector感染细胞组(图5B);Lv-SYVN1感染细胞组未发生肺部转移,经Lv-Vector转染细胞组3只发生肺部转移(图5C)。这提示过表达SYVN1可抑制异种原位移植的骨肉瘤细胞增殖和远处转移。
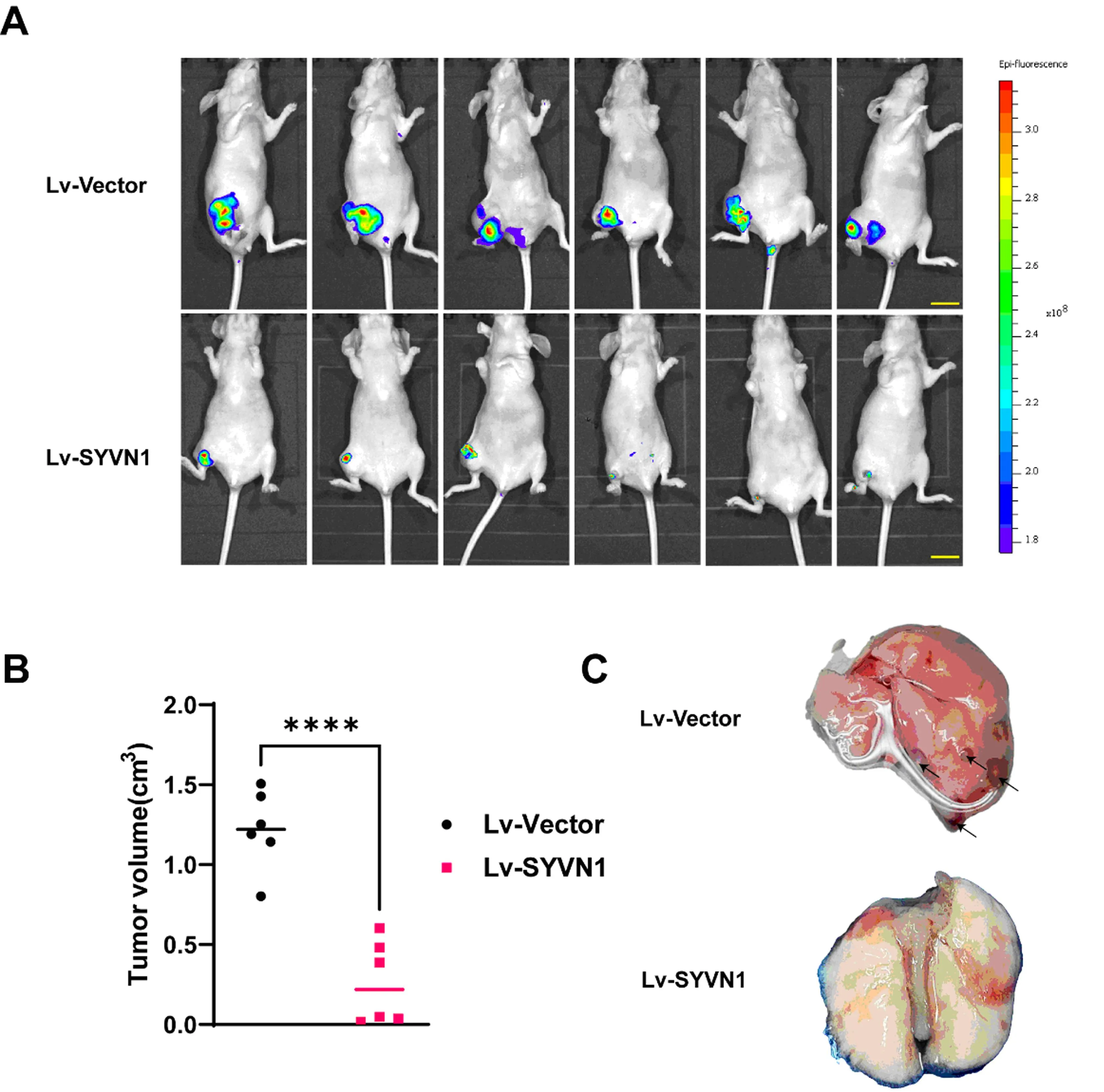
注:A为裸鼠原位成瘤实验;B为裸鼠原位骨肉瘤成瘤体积测量;C为大体计数裸鼠肺转移结节,黑色箭头为转移灶。
3 讨论
失巢凋亡是细胞脱离原位灶后,自发性产生凋亡的一种独特现象,这在肿瘤转移的过程中是必经的步骤[4]。肿瘤经循环系统转移的步骤包括从原位病灶脱落,穿过细胞外基质,经由血管内皮进入微小血管,产生失巢凋亡抵抗特性,进而经循环系统(又称为循环肿瘤细胞)到达适宜的再生长部位,形成转移肿瘤灶[5],而在这个过程中,最重要的就是逃避失巢凋亡。课题组在前期研究中,已成功通过悬浮培养法构建失巢凋亡抵抗的细胞株AR-143B,并观测到FASN在其中表达量高于普通贴壁培养的143B,沉默FASN会显著降低骨肉瘤细胞的失巢凋亡抵抗能力[2],但FASN升高的机制当时未作进一步探讨。
FASN作为经典的癌基因之一,与肿瘤细胞恶性程度息息相关,其升高往往预示着肿瘤预后不良[6]。Yasmina等报导FASN促进脂质相关的磷脂结构合成原料生成,这可以增加自噬小体的产生,从而支持肿瘤细胞在更恶劣的环境中生存下去[7],Zou等[8]报导在肝癌中由于脂质堆积异常导致肝癌细胞脂代谢重编程,升高的FASN在其中利用丰富的内源性脂质合成原料生成更多可供肿瘤细胞增殖和胞内转运的膜及囊泡结构--这一效应是由SREBP1驱动FASN转录实现的。但一直以来有关FASN在肿瘤转移过程中升高的机制却未见阐明。在本研究中,我们利用失巢凋亡抵抗模型,模拟肿瘤在远处转移过程时经历的循环肿瘤细胞阶段,并利用分子生物学手段结合蛋白组学定量技术,初步探究了FASN在骨肉瘤细胞转移过程中的升高是由于其E3连接酶SYVN1对其泛素化修饰减少导致的,并首次在骨肉瘤中初步研究了E3连接酶SYVN1的功能,其能够降解FASN而对肿瘤失巢凋亡抵抗产生抑制作用。在体内实验中,GFP标记的骨肉瘤细胞难以在活体成像中探测出肺部微小转移灶,而大体观察结果提示仅有Lv-Vector组别细胞存在三只肺转移个体,总体结果提示过表达SYVN1可以有效抑制骨肉瘤细胞原位成瘤及远处转移,但在过表达SYVN1的组别中,存在三只成瘤效果不佳的个体,且总体肺转移率较低。这是本实验的不足之处,但SYVN1对骨肉瘤成瘤的抑制效果仍是显著的,不影响本文结论成立。
FASN的抑制剂一直以来都备受关注,从最早发现的FASN天然抑制剂浅蓝菌素[9],到最近新开发的FASN蛋白分子靶向抑制剂6p[10],针对这一前景广阔的癌症抑制靶点,众多学者不断致力于开发更精准的分子抑制剂,并取得了显著的体内外效果[11],并在一些疾病(如酒精性脂肪肝)的动物模型治疗上取得了令人满意的疗效[12],然而,直至今日,FASN抑制剂在肿瘤中的效果仍然不完全令人满意。
直接靶向FASN将影响其他代谢正常细胞的脂质从头合成过程,从而导致一系列的毒副作用[13]。所以,针对异常升高的FASN进行精准降解,才是将这个强力肿瘤治疗靶点应用性提高的重要措施。泛素化是细胞内最常见的蛋白质平衡调控机制,异常表达或错误折叠的蛋白大多经泛素-28S蛋白酶体系统调控,降解为氨基酸,重新加入蛋白质合成的过程[14]。促进癌基因或肿瘤代谢关键通路分子的泛素化修饰已成为开发肿瘤靶向药物的常用手段,其优势在于利用细胞内天然的降解回收系统,让水平异常靶标分子的水平回复一个趋近正常的状态,而不是针对所有细胞的同一靶蛋白同步作用,其导致的毒副作用将相对更小[15]。靶向FASN的泛素化降解可以更好地控制这一癌基因异常高表达导致的肿瘤恶性程度增加,同时更好的保护正常细胞中的代谢不受到单分子抑制剂的广泛影响。FASN泛素化机制的研究将促进针对这一分子的精准靶向调控得到进一步发展。本研究已阐述SYVN1调控FASN泛素化降解的作用,但SYVN1水平在失巢凋亡的骨肉瘤细胞是为什么而降低,后期我们将对这方面进一步探索。
综上所述,我们研究发现在失巢凋亡抵抗的骨肉瘤细胞中SYVN1表达下调,从而减少FASN泛素化水平使其得到稳定,并且SYVN1能够抑制骨肉瘤失巢凋亡抵抗能力,减少其远处转移。本研究初步阐述了FASN与SYVN1在骨肉瘤发展进程中的作用,将为后期FASN与SYVN1作为骨肉瘤潜在的治疗靶点提供理论基础,为未来骨肉瘤的治疗提供新的理论依据。
猜你喜欢
特产研究(2022年6期)2023-01-17
植物保护(2021年5期)2021-10-12
天然产物研究与开发(2018年5期)2018-06-13
郑州大学学报(医学版)(2016年4期)2016-08-11
癌症进展(2016年10期)2016-03-20
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
中国医学科学院学报(2015年5期)2015-03-01
现代检验医学杂志(2015年2期)2015-02-06
火炸药学报(2014年1期)2014-03-20
四川生理科学杂志(2014年3期)2014-02-28
