
关帝山3个典型森林群落优势种的氮素利用策略差异
2024-03-12 06:50张芸香吕世琪刘泰瑞李晋芳郭晋平
林业科学 2024年2期
张芸香 吕世琪 刘泰瑞 李晋芳 郭晋平
(1. 山西农业大学林学院 太谷 030801; 2. 山西文峪河国家级湿地公园管理局 交城 030599)
土壤氮素供应水平是森林生产力的主要限制因子(Aberet al.,1998; Näsholmet al.,1998),准确认识森林群落优势种生长发育对氮素的需求和不同土壤氮素供应条件下的氮素利用效率,对把握树种在群落演替过程中的替代关系,预测森林群落组成结构动态,理解森林生产力形成和影响机制具有重要意义(Yin,1994;陈伏生等,2004)。北温带森林生长通常受到土壤有效氮的限制,一定程度的外源氮输入可以促进森林生物量的生长和碳固定,形成氮促碳汇效应(冯继广等,2020;王银柳等,2021),进而导致森林生态系统碳循环过程的反馈性响应(Chenet al.,2015),还能够通过调节细根生长,显著提高森林地上和地下生产力(Nadelhofferet al.,1999;Ciaiset al.,2008;宋平等,2017)。但对上述认识在不同区域和研究对象之间一直存在争议。Magill等(2004)指出,长期施氮显著抑制了美国赤松(Pinus resinosa)针叶林的生长,而明显促进硬阔叶林优势种黑栎(Quercus velutina)和北美红栎(Quercus rubra)的木质生物量生长,即长期施氮对温带针叶林和阔叶林有相反的作用。Högberg等(2006)研究发现,长期中低剂量的氮添加能够促进欧洲赤松(Pinus sylvestris)林的生长,但高剂量的施氮却表现为抑制作用。方运霆(2005)等研究表明,较高剂量施氮(100 kg·hm-2a-1)能促进鼎湖山林区马尾松(Pinus massoniana)林乔木优势种的胸径生长,但降低季风常绿阔叶林优势种木荷(Schima superba)和锥栗(Castanea henryi)的胸径生长。Tu等(2011)研究发现,施氮能显著促进华西雨屏区苦竹(Pleioblastus amarus)人工林的净初级生产力。Wei等(2012)对杉木(Cunninghamia lanceolata)林的研究发现,20~30 kg·hm-2a-1是林地最佳施氮剂量。可见,不同区域和森林类型,特别是不同群落优势种,可能具有不同的氮素利用功能特性,对土壤不同氮素供应水平也可能具有不同的响应模式,且植物对不同形态氮素具有不同的利用潜力和偏好(曹翠玲等,2004)。然而,对处于氮限制影响下的森林类型,短期氮输入是否显著提高森林和林木生物生产力,不同森林群落优势种的氮素利用策略是否存在显著差异,这些问题的研究还很不充分。因此,开展不同森林群落的多梯度施氮试验,探究群落优势种生物生产力对土壤氮素供应水平的响应,可加深理解不同群落优势种的氮素利用功能特性及氮素利用策略差异。
华北落叶松(Larix principis-rupprechtii)林、白桦(Betula platyphylla)林和沙棘(Hippophae rhamnoides)林是华北山地广泛分布的3种森林类型,在山西省关帝山林区山地森林植被中具有典型代表性。本研究选择这3种典型森林群落为研究对象,通过野外多梯度施氮控制试验,分析群落优势种新生枝叶氮含量、枝叶新增生物量和氮素利用效率随土壤施氮量的变化,分析比较其响应方式的类型间差异,阐明不同森林群落优势种氮素利用特性的分异以及形成的氮素利用差异化类型,为氮沉降背景下的森林质量提升和精准经营提供依据。
1 研究区概况
研究地区在山西省关帝山林区,属于暖温带落叶阔叶林区域。年平均气温4.3 ℃,年均降水量822.6 mm,年日照1 900~2 200 h,年蒸发量1 100~1 500 mm,全年无霜期100~130 天。主要森林类型有:华北落叶松林、油松(Pinus tabuliformis)林、青杄(Picea wilsonii)林和白杄(Picea meyeri)林等针叶林,白桦林、山杨(Populus davidiana)林、青杨(Populus cathayana)林和辽东栎(Quercus wutaishanica)林等落叶阔叶林、针阔混交林,以沙棘、土庄绣线菊(Spiraea pubescens)、刺果茶藨子(Ribes burejense)、黄刺玫(Rosa xanthina)和胡枝子(Lespedeza bicolor)等为主的灌木林。华北落叶松林、白桦林和沙棘林是当地中高海拔山地和河谷地段的典型森林群落类型。
试验地于2017年5月初设在文峪河上游孝文山林场(111°22′—111°33′E,37°45′—37°55′N)的大草坪沟两侧缓坡带,海拔1 820~1 830 m,立地条件基本一致,土壤为山地淋溶褐土,厚度60~80 cm,质地中壤至沙壤。3种森林群落为华北落叶松林、白桦林和沙棘林,林下植物主要有金花忍冬(Lonicera chrysantha)、黄瑞香(Daphne giraldii)、美蔷薇(Rosa bella)、毛榛(Corylus mandshurica)、胡枝子、土庄绣线菊、山荆子(Malusbaccata)、刺果茶藨子、草问荆(Equisetum pratense)、披针叶苔草(Carex lanceolata)和糙苏(Phlomis umbrosa)等。在每个森林群落的林地内部,距离林缘≥10 m,分别设面积50 m×30 m的试验样地,并分为3个面积50 m×10 m的重复试验区组,3个林型共设了9个区组,样地概况见表1。
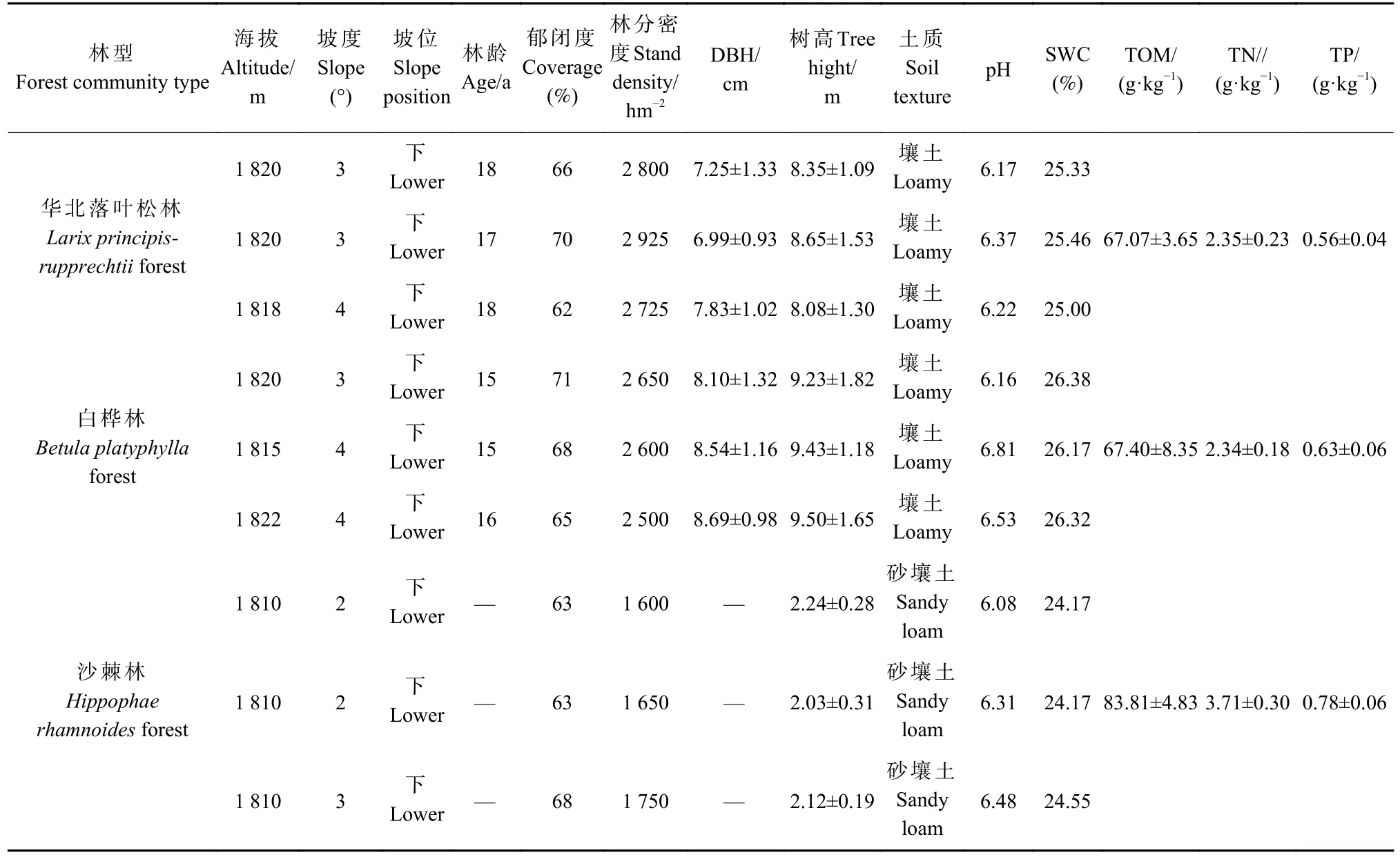
表1 3种森林群落类型的试验样地和区组的立地条件概况①Tab. 1 Site condition of experimental plots and blocks in 3 forest community types
2 研究方法
2.1 试验设计和施氮方法
根据样地土壤基础氮含量,按施氮后30 cm土层平均氮含量不超过最高值的1.5倍并形成明显梯度的原则,参考国内外有关施氮试验文献(Vestgardenet al.,2001;Luet al.,2013;Tuet al.,2014;朱荣玮等,2019),设5个施氮量:对照N0(0 g·m-2a-1)、低剂量N1(9 g·m-2a-1)、中剂量N2(18 g·m-2a-1)、高剂量N3(27 g·m-2a-1)和超高剂量N4(36 g·m-2a-1)。将前述每个林型的3个区组分别划分成5个10 m×10 m的样方,随机布设5个施氮量处理,每个处理在3个区组中形成3次重复。以含氮46.4%的尿素CO(NH2)2为施氮材料,按5个施氮量计算出不同处理的尿素用量,分为两等份,先后于5月初和6月初,溶于5 L蒸馏水中,均匀喷洒到样地内,对照N0喷洒等量蒸馏水。
2.2 样品采集与测定
2018年7月(生长旺盛期),在华北落叶松林和白桦林样地中,按乔木层、灌木层和草本层分别取样,在沙棘林中按灌木层和草本层分别取样。在乔木层和灌木层中选择代表性样株,根据枝条形态特征,结合枝条颜色和芽痕等特征,识别新生枝和叶,按树冠高度分层剪取适量新生枝,摘取针叶或叶片,枝和叶分别混匀后取适量(0.3~0.5 kg)样品;草本层取样时,将(1 m×1 m)样方地上部分全部收获,混匀后取适量样品。所取样品分别装入塑料自封袋,贴好标签,带回基地,用恒温鼓风干燥箱105 ℃杀青30 min,85 ℃持续烘干至恒质量,采用凯氏定氮仪(FoodALYT D5000,德国)测定枝和叶烘干样品氮含量。
9月(生长末期)的取样,以所设样地为单位,剪取全部新生枝,摘取针叶或叶片,草本层仍按样方进行地上部分全部收获后带回基地,用电子天平分别称量并记录乔木层、灌木层和草本层的新生枝、叶、草鲜质量,按前述方法烘干至恒质量,称量记录样品干质量即生物量,分别推算单位面积新生枝、叶、草本的生物量。分别从烘干的枝、叶和草本样品中取氮含量分析样品,仍用前述方法测定样品氮含量。
2.3 数据处理与统计分析
群落新生枝叶生物量(TBM)为各层次新生枝叶生物量之和,计算公式为:
式中:BMc、BMb和BMh分别为乔木层、灌木层和草本层新生枝叶生物量,BMc= BMcs+ BMcl,BMb=BMbs+BMbl,BMcs和BMcl分别为乔木层新生枝和新生叶的生物量;BMbs和BMbl分别为灌木层新生枝和新生叶的生物量。
群落新生枝叶氮储量(TNS)是群落单位面积各层次新生枝叶氮储量之和,计算公式为:
式中:NSc、NSb和NSh分别为乔木层、灌木层和草本层的新生枝叶氮储量,NSc =NCcs×BMcs+NCcl×BMcl,NSb=NCbs×BMbs+NCbl×BMbl,NSh=NCh×BMh,NCcs为乔木层新生枝氮含量,NCcl为乔木层新生叶氮含量,NCbs为灌木层新生枝氮含量,NCbl为灌木层新生叶氮含量,NCh为草本层氮含量。
群落氮利用效率(NUE)为群落新生枝叶生物量与新生枝叶氮储量之比(Finziet al.,2007),公式为:
对数据进行方差齐性和一致性检验后,利用SPSS 17.0(SPSS Inc., USA)软件中的one-way ANOVA进行方差分析,并在α=0.05水平下利用LSD法进行多重比较。通过R4.0.5软件的Lavan包和semplot包完成结构方程建模,利用R 4.0.5和SigmaPlot 12.0软件进行图形化和变化趋势分析。
3 结果与分析
3.1 不同森林群落优势种新生枝叶氮含量随施氮量的变化
分析3种群落优势种新生枝叶氮含量在生长盛期(7月)和生长末期(9月)随施氮量的变化趋势和差异(图1)。华北落叶松在2个生长期的叶氮含量都显著(P≤0.01)高于枝氮含量,生长盛期显著(P≤0.01)高于末期;枝和叶氮含量在施氮量之间的差异总体显著(P≤0.05),且随施氮量的增加而出现明显饱和点,其中枝的盛期最高氮含量为11.01 mg·g-1,末期为9.50 mg·g-1;叶的盛期最高氮含量为25.56 mg·g-1,末期为20.79 mg·g-1。
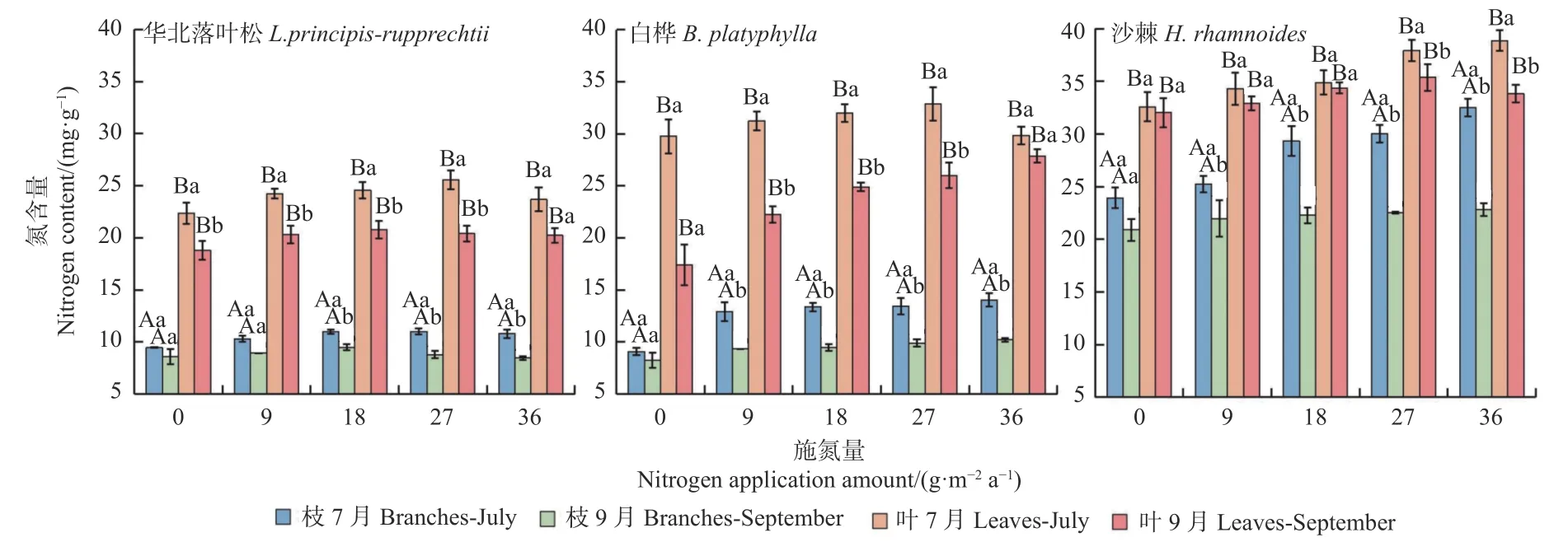
图1 3个森林群落优势种新生枝和叶氮含量在2个季节随施氮量的变化Fig. 1 Seasonal changes of nitrogen content of new branches and leaves of 3 dominant species in 3 forest community types with different nitrogen application amount
白桦(图1)在2个生长期的新生叶氮含量显著(P≤0.01)高于新生枝氮含量,生长盛期新生枝、叶氮含量都显著(P≤0.01)高于生长末期;生长末期叶氮含量随施氮量增加而持续升高,没有出现明显的饱和点;生长盛期叶氮含量在施氮量N3(27 g·m-2a-1)时达到最高,相应的土壤氮含量为32.88 mg·g-1。
沙棘(图1)在2个生长期的新生叶氮含量显著(P≤0.01)高于新生枝氮含量,且生长盛期新生枝、叶氮含量都显著(P≤0.01)高于生长末期,但2个生长期枝和叶氮含量都高于华北落叶松和白桦,而枝与叶氮含量的差别和2个生长期之间的差别都小于华北落叶松和白桦;生长盛期的枝和叶都在最大施氮量N4(36 g·m-2a-1)才出现最大氮含量,而生长末期的叶饱和施氮量明显在N3(27 g·m-2a-1),相应的土壤饱和氮含量为35.38 mg·g-1。
3.2 不同森林群落地上新增枝叶生物量随施氮量和枝叶氮含量的变化
从3个群落单位面积新生枝叶生物量(TBM)随施氮量的增大而提高的趋势来看(图2),华北落叶松林和白桦林在施氮量27 g·m-2a-1时出现明显拐点(达到最大),华北落叶松林表现最为明显,而沙棘林在所设施氮量范围内没有表现出达到最大值,但在施氮量36 g·m-2a-1时表现出增长趋于平缓。其中,华北落叶松林的增幅最大,白桦林的增幅最小。3个群落的TBM与群落新生枝叶氮含量之间的线性正相关关系有显著差异(图3),随新生枝叶氮含量提高,TBM的增长率表现为华北落叶松林>沙棘林>白桦林。
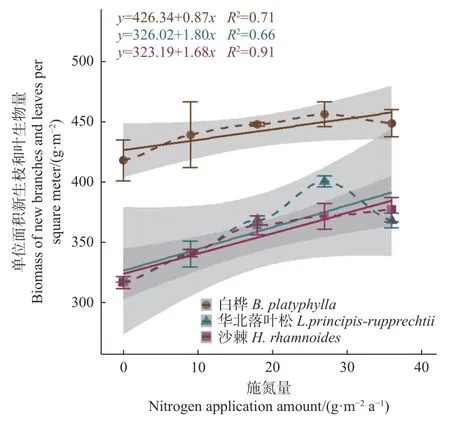
图2 3 个群落新生枝和叶生物量随施氮量的变化Fig. 2 Biomass changes of new branches and leaves with nitrogen application amount in 3 forest communities
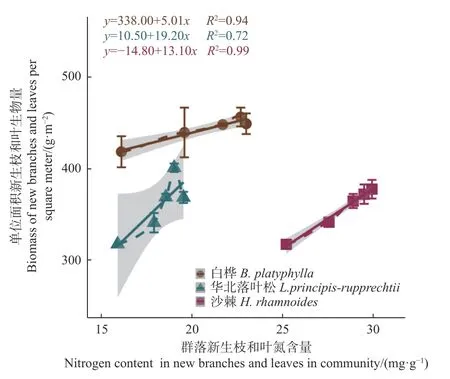
图3 3 个群落新生枝叶生物量随新生枝和叶氮含量的变化Fig. 3 Biomass changes of new branches and leaves with nitrogen content of new branches and leaves in 3 forest communities
3.3 不同森林群落氮素利用效率随施氮量和新生枝叶氮储量的变化
分析群落氮利用效率(NUE)随施氮量的变化(图4)发现,在对照条件下,华北落叶松林和白桦林之间差异不显著,但都显著高于沙棘林;随施氮量增加,3个森林群落的NUE都随施氮量的增大而呈显著降低趋势,三者的差异更趋明显,且表现为华北落叶松林>白桦林>沙棘林。进一步分析NUE与新生枝叶氮储量之间的关系(图5)表明,3个群落都呈显著负相关关系,在对照条件下,华北落叶松林的NUE略高于白桦林,两者都高于沙棘林,且都随新生枝叶氮储量的升高而降低,三者的下降率表现为白桦林>华北落叶松林>沙棘林。
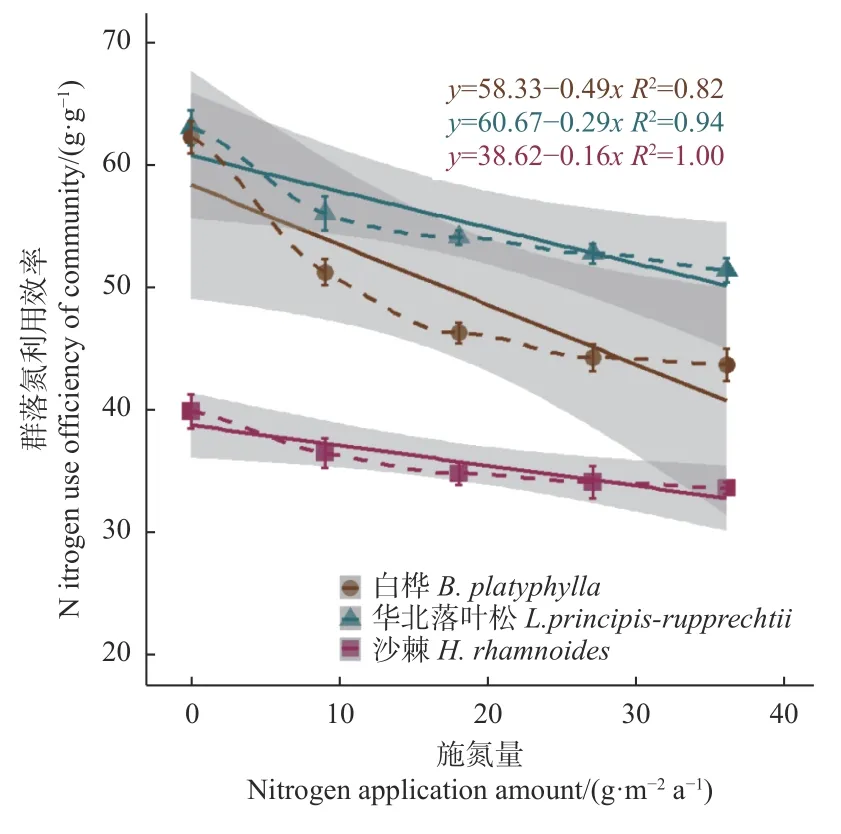
图4 3 个群落氮利用效率随施氮量的变化Fig. 4 Changes of NUE of three forest communities with nitrogen additions
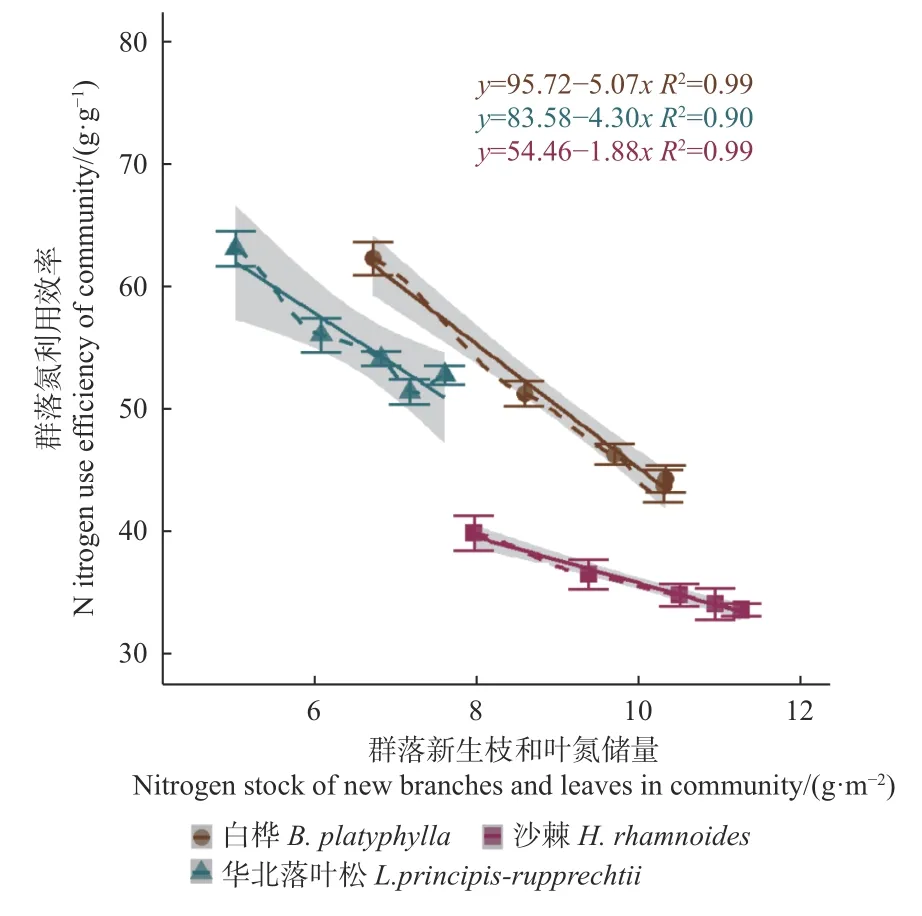
图5 3 个群落氮利用效率随新生枝叶氮储量的变化Fig. 5 Changes of NUE of three forest communities with nitrogen stock in new branches and leaves
3.4 土壤氮添加对群落新增生物量的作用路径
通过R4.0.5的Lavan包和semplot包建立结构方程模型,进一步解读群落类型和土壤施氮量对新生枝叶生物量积累的作用路径(图6)。从施氮量和群落类型出发,路径系数表明,群落类型对群落新增生物量的直接作用很小,而施氮量几乎没有直接作用,但两者对群落新生枝叶氮储量和氮素利用效率有显著的直接作用,且群落类型的作用强度都显著高于施氮量;群落新生枝叶氮储量与氮素利用效率两者之间的直接作用很微弱,而群落新生枝叶总氮含量和氮素利用效率都显著正向驱动群落的生物量积累。
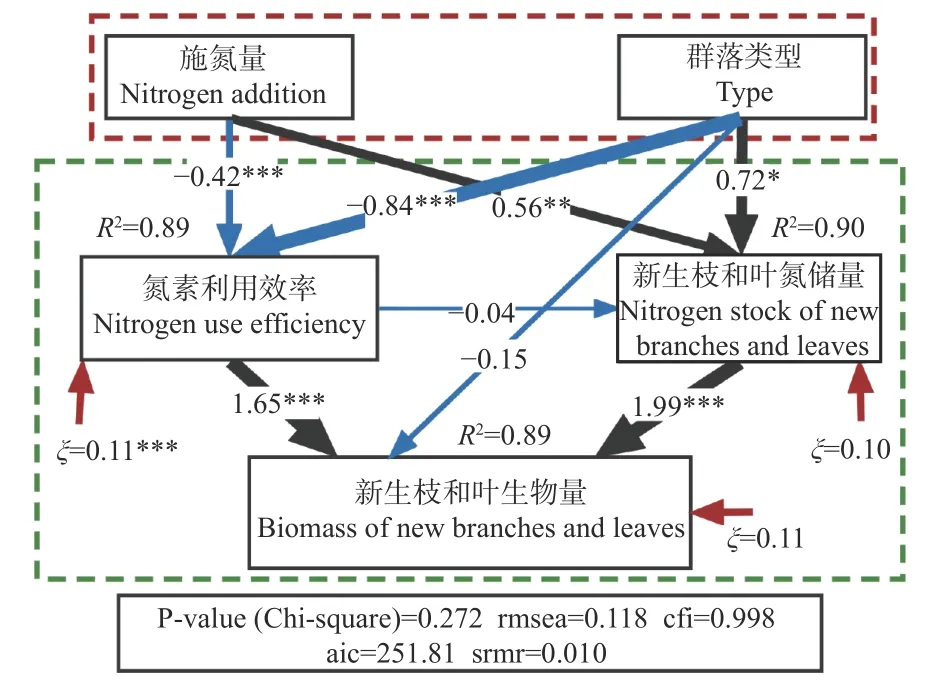
图6 群落新生枝叶生物量影响因子作用路径Fig. 6 Effector and functional pathways of the new branches and leaves biomass
4 讨论
4.1 森林群落优势种新生枝叶氮含量对土壤施氮的响应
不同森林群落优势种新生枝叶氮含量对土壤施氮具有不同的响应模式。有研究表明,施氮能显著提高兴安落叶松(Larix gmelinii)幼苗叶氮含量(傅民杰等,2009),也可显著提高秦岭西部华北落叶松人工林新生叶的氮含量,并因林地土壤全氮含量较低(1.16 g·kg-1)所致(徐福利等,2014);但也有研究显示,兴安落叶松持续5年的施氮处理对提高针叶氮含量无显著作用,且成年乔木树种新生叶氮的主要来源并非根系直接吸收而是茎的氮储存(赵琼等,2010)。本研究结果表明,3个群落优势种华北落叶松、白桦和沙棘的新生枝叶氮含量都以生长盛期显著高于生长末期,且不因土壤施氮而改变;新生枝叶氮含量以沙棘最高,华北落叶松最低,对土壤施氮都表现出正响应,但响应强度表现为华北落叶松>白桦>沙棘;土壤施氮能显著提高3个群落优势种的新生枝叶氮含量,但都表现出具有一定冗余度的饱和氮含量,且树种间差异显著,新生枝的变幅和冗余度高于新生叶,3个树种枝和叶的饱和氮含量顺序不因土壤施氮而改变。
4.2 森林群落地上部分新增生物量对施氮的响应
森林群落地上部分生物生长量(新增生物量)及其对土壤施氮的响应模式因不同优势种而异。多个树种的幼苗短期施氮试验表明,施氮能显著促进苗木生长(郭盛磊等,2005;吴茜等,2011),而一些学者坚信,持续施氮对森林生产力的长期影响往往需要数年时间才能明确表现出来(王建宇等,2018);事实上,美国东北部硬阔叶林的长期施氮显著提高了森林净生产力(Magillet al.,2004),而兴安落叶松持续5年的试验说明,施氮对提高生物量增长无显著作用(赵琼等,2010),对北美红松(Pinus resinosa)林的生长甚至表现出抑制作用(Keltyet al.,2004);在对鼎湖山主要森林类型的研究中,针阔叶树种的胸径生长量都对外源施氮有显著正响应,且针叶树种的响应高于阔叶树种(方运霆等,2005)。关帝山林区的华北落叶松林凋落物中树枝的占比大,凋落叶氮含量低,且分解速率较低(郭晋平等,2009;张芸香等,2011),或者其他原因造成树木生长的氮限制(Boet al.,2020)。对中国东北温带针阔混交林主要优势种水曲柳(Fraxinus mandshurica)、黄菠萝(Phellodendron amurense)和红松(Pinus koraiensis)的氮素形态偏好试验表明,在其他条件一致的情况下,树种对氮素的总需求量不受氮素形态的影响(周明欣,2019)。本研究中,施氮对华北落叶松地上部生物量积累的短期效应在施氮量27 g·m-2a-1时达到饱和,远高于一些长期施氮试验的剂量(小于15 g·m-2a-1)(Xiaet al.,2008;徐福利等,2014),说明其生长明显受限于土壤氮素供应不足,而白桦林和沙棘林的土壤氮限制较轻;本研究中的3个森林群落新增生物量均因施氮而提高,且随施氮量的增大而增大,其效应强度表现为华北落叶松林>白桦林>沙棘林;新增生物量随施氮量的增大均表现出效应极限,即出现饱和施氮量和枝叶饱和氮含量,虽然其具体数值未必精准,但三者的差异已得到明确表达。
4.3 群落氮素利用效率对土壤施氮的差异化响应
研究地区3个群落氮素利用效率及其对土壤施氮的响应模式有显著差异。有研究表明,华北落叶松针叶氮素重吸收效率较高,且速生期显著高于幼林期和近熟期,外源施氮导致重吸收率降低,降幅表现为幼林期>近熟期>速生期(张文等,2018);沙棘是固氮树种,不仅通过固氮显著增加土壤氮含量,而且氮需求和吸收量较大(李根前等,2000;阮成江等,2002),而白桦林的土壤氮矿化速率高于华北落叶松林(陈立新等,2012)。本研究中,3个群落的氮利用效率有显著差异,表现为华北落叶松林>白桦林>沙棘林,且三者的差异随施氮量的增大而逐步增大;3个森林群落的氮素利用效率均随枝叶氮含量的增大而下降,以白桦林的下降率最高,沙棘林的敏感性最低。
4.4 土壤氮添加对群落新增生物量积累的作用路径
通过建立结构方程模型并解析其路径系数,可以理解和把握各因素的关系和效应强度(Hildreth,2012)。不同群落类型主要反映在物种组成和结构上,特别是优势种的差别(Heet al.,2010)。本研究结果表明,群落类型直接决定了群落的新生枝叶氮储量和氮素利用效率,进而影响群落新增生物量,是群落对施氮量差异化响应的根本成因和主导因素,而施氮量通过改变土壤供氮水平作用于新生枝叶氮含量,影响其氮储量和氮素利用效率,进而调节或影响群落新增生物量。
5 结论
华北落叶松林、白桦林和沙棘林3个森林群落新增生物量的差异主要取决于群落类型及其优势种的差异,直接受群落优势种的氮素利用效率和新生枝叶氮含量的影响,并受土壤供氮水平的调节;三者在氮需求、氮利用效率、环境氮敏感性、饱和氮供应水平4个方面表现出氮素利用策略上的分异;氮素利用策略类型依次为“低需求-高效率-高敏感-低饱和”、“较低需求-低效率-较高敏感-较低饱和”和“固氮型-高需求-低效率-低敏感-高饱和”。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
今日农业(2021年10期)2021-11-27
当代陕西(2020年17期)2020-10-28
山东林业科技(2019年2期)2019-06-03
现代营销(创富信息版)(2018年2期)2018-08-15
中成药(2018年3期)2018-05-07
中成药(2017年7期)2017-11-22
农业环境科学学报(2017年2期)2017-03-20
传奇故事(破茧成蝶)(2015年1期)2015-02-28
防护林科技(2015年3期)2015-02-18
