
脑膜免疫在中枢神经系统疾病中的研究进展
2024-03-13 04:47高俊英
南京医科大学学报(自然科学版) 2024年3期
孙 芃,高俊英
南京医科大学基础医学院,江苏 南京 211166
1 脑膜的结构及相关免疫细胞
1.1 脑膜的基本结构
过去认为,脑膜位于颅骨和脑之间,从外向内分别由硬脑膜、脑蛛网膜和软脑膜3 层组成。硬脑膜是厚而坚韧的双层膜,其外层附着于颅骨内面,内层为脑膜层,两层膜在一些特定部位相互分离,形成硬脑膜窦。硬脑膜上有大量神经、血管分布,并含有淋巴管。脑蛛网膜薄而透明,缺乏血管和神经,由一层外层细胞组成,包围着蛛网膜下腔,其间充满脑脊液(cerebrospinal fluid,CSF),大部分脑脊液由脑室内的脉络丛(choroid plexus,CP)产生,跨越蛛网膜下腔的富含胶原的薄层结构,称为小梁。脑蛛网膜的细胞通过紧密连接连接在一起,是中枢神经系统实质的第一个不透水屏障,因此,分子不能从硬脑膜自由扩散到蛛网膜下腔,其作为一层物理屏障将CSF 与脑组织间质液(interstitial fluid,ISF)分隔开[1]。脑蛛网膜在上矢状窦两侧形成许多绒毛状突起突入上矢状窦内,称为蛛网膜粒,参与CSF流入静脉血的重吸收[2]。
1.4 统计学分析 采用SPSS 21.0统计软件对数据进行分析处理,计量资料用表示,组间比较采用t检验,计数资料用例(%)表示,组间比较采用χ2检验,P<0.05为差异有统计学意义。
近期,研究人员在蛛网膜和软脑膜之间发现了第四层脑膜,将蛛网膜下隙划分为浅部、深部两个空间,其在结构和免疫表型方面与外周脏器所覆盖的间皮组织相似,研究团队将其命名为蛛网膜下淋巴样膜(subarachnoid lymphatic -like membrane,SLYM)[3]。SLYM位于蛛网膜下隙,与脑膜静脉窦紧密排列,内含血管和免疫细胞,故认为SLYM参与了CSF 和静脉血之间小分子溶质的物质交换,并通过在小鼠蛛网膜下隙注射微球体以及示踪剂,发现SLYM可以限制多数小分子肽和蛋白质在蛛网膜下隙浅、深两层之间交换,而其参与脑膜免疫以及液体引流的具体机制尚待进一步探索。脑蛛网膜和软脑膜合称软膜,但随着SLYM的发现,软膜这一概念是否需要被扩充或修正,值得探究。
一种是基于字符串匹配(词典)的算法,它是将需要分析的语言与词典中的词条进行匹配的方法。学者们研究字符串匹配时多种策略,如正向或逆向扫描、最大或最小的匹配原则、纯分词法或包含标注的一体化方法等;
1.2 脑膜相关的免疫细胞
生理状态下,脑实质所含有的免疫细胞仅有小胶质细胞。该类细胞起源于原始卵黄囊细胞,是一种终身定居于脑实质的特殊类型巨噬细胞。此外,通过CSF 引流,脑膜中也存在来源于骨髓造血干细胞和淋巴样干细胞的免疫细胞[4]。3层脑膜中,硬脑膜含有大量的免疫细胞,如树突细胞(dendritic cell,DC)、肥大细胞(mast cell,MC)、固有淋巴细胞(innate lymphoid cell,ILC)、脑膜巨噬细胞、T 细胞和B细胞[5]。虽然软脑膜上也含有免疫细胞,但少于硬脑膜,主要含有巨噬细胞、DC 和MC[5]。CSF 中仅含有记忆T细胞、少量B细胞和单核细胞[6]。位于中枢神经系统的非小胶质巨噬细胞被称为屏障相关巨噬细胞(border-associated macrophages,BAM),过去,所有组织特异性巨噬细胞都被认为来源于骨髓祖细胞,然而通过大规模单细胞RNA测序结合原基作图,发现硬脑膜与伴血管的非实质巨噬细胞均源自胚胎时期的卵黄囊髓样祖细胞,并在许多方面与小胶质细胞类似。脑膜、CP 和脑血管周围空间由BAM填充,其主要表达蛋白质标志物CD206,参与抗原识别和递呈,维护血-脑屏障(blood-brain barrier,BBB)的完整[4]。硬脑膜被蛛网膜从中枢神经系统的其他部分隔离出来,因此应该有自己特异的免疫细胞亚群。这些脑膜相关淋巴细胞参与免疫屏障的具体机制尚待进一步探索。
2 脑膜免疫渠道
2.1 胶质淋巴系统
CSF 在循环过程中,通过覆盖在大脑微血管周围的星形胶质细胞(astrocytes,AC)终足,与ISF进行密切的物质交换。其中,CSF围绕动脉进行流动,其所在的动脉旁间隙(Virchow-Robin spaces,VRS)的边界由血管腔壁和AC终足构成[7]。Iliff等[8]在活体双光子显微镜下通过免疫荧光追踪,证实经过蛛网膜下CSF 循环使脑实质中的溶质进入CSF,并发现此通道依赖血管周围AC 终足表达的水通道蛋白-4(aquaporin 4,AQP4)。他们将这种CSF 与ISF 之间进行小分子物质交换的系统命名为glymphatic 系统,即胶质淋巴系统。一方面,该系统的物质交换依赖胶质水通道;另一方面,其清除废物的机制又与外周淋巴系统相似。基于胶质淋巴系统的清除特性,研究者将其与阿尔茨海默病(Alzheimer’s disease,AD)中的大分子物质,如Aβ和tau蛋白沉积联系起来,认为淋巴胶质系统的功能退行可能导致Aβ或tau 蛋白沉积[9]。而增强其清除能力以缓解AD 病理进程的具体机制和临床应用,尚待进一步探索。
《坤舆图》说:“狮性最傲,遇者亟俯伏,虽饿亦不噬……又最有情,受人德必报。”所以狮子纹饰的精神之美也是历来受人称颂。狮子勇猛但是性情仁义,重情重义,知恩图报,狮子纹饰于是有四海升平,万国咸宁的美好寓意。
2.2 脑膜淋巴管(meningeal lymphatic vessel,MLV)
MLV是硬脑膜上沿静脉结构排列的管状结构,除了静脉窦周围,颅底也尤为发达,具有周围淋巴管的许多相同属性,如生理条件下表达淋巴管内皮细胞的大部分经典标志物,包括prospero homeobox-1(PROX1)、血管内皮生长因子受体3(vascular endothelial growth factor receptor 3,VEGFR3)、淋巴管内皮透明质酸受体-1(lymphatic vessel endothelial hyaluronan receptor-1,LYVE1)、podoplanin和C-C基序趋化因子配体21(chemokine ligand 21,CCL21)等,但无平滑肌细胞。其总体结构和分布又具有某些特点,如脑膜淋巴管不表达如整合素-α9(integrin α9)等瓣膜特征性标志物,其直径小于外周淋巴管[10]。
CCR7-CCL21 途径被认为是T 细胞和DC 进入淋巴系统及在淋巴系统中迁移、循环的主要途径。Louveau 等[18]将CCR7-WT 和CCR7-KO 的T 细胞以1∶1的比例共注射到naïve 小鼠的脑脊液中,发现与CCR7-WT T 细胞相比,CCR7-KO T 细胞在dCLN 中的引流显著减少。流式细胞荧光分选技术(fluorescence activated cell sorting,FACS)分析CCR7 启动子下表达GFP 的小鼠脑膜T 细胞,发现约40%的脑膜CD4+T 细胞表达CCR7。该小鼠脑膜免疫染色显示CCR7+T细胞主要与表达CCL21的淋巴内皮细胞密切相关。由此可见,T 细胞将沿MLV 最终进入dCLN,从而离开脑膜,此过程依赖CCR7-CCL21 途径。但MLV 对免疫细胞的具体迁移机制尚待进一步确认。另外,MLV 对T 细胞等免疫细胞的激活是否存在影响及其途径也值得深入探究。
2.2.1 MLV引流CSF的途径
除经典的蛛网膜粒硬脑膜窦吸收途径外,CSF也可以沿着颅神经鞘,通过MLV或鼻腔嗅觉系统淋巴管流入外周淋巴管,间接进入血液循环。多种动物实验表明,在正常颅内压力下,很少量的CSF通过经典途径转运到颅静脉系统,该转运途径仅在病理性颅内压增高的情况下发挥作用[13]。基于此,大量研究开始聚焦CSF 的淋巴引流途径。Louveau 等[11]发现,给小鼠静脉注射伊文思蓝30 min 后,染料在MLV 与dCLN 中均被检测到,但在周围的非淋巴组织中几乎没有分布。结扎dCLN 的输入淋巴管后,与假手术对照组比较,结扎小鼠的dCLN 无明显染料累积,由此可见,MLV和dCLN之间直接偶联。而Aspelund 等[12]通过建立血管内皮生长因子缺失的转基因小鼠模型,也发现MLV发育不良会导致脑内大分子物质清除减弱,以及阻碍蛛网膜下腔向dCLN的引流途径。由此可见,MLV 是ISF 及其来源的细胞成分及可溶性代谢产物通过CSF 引流到dCLN 的主要途径。
脑膜作为脑内免疫细胞的驻留场所,通过解剖结构和其免疫细胞及细胞因子发挥脑内抗原和代谢物清除的作用,由此实现脑内的免疫监视作用(图1)。而在脑膜维护大脑免疫稳态的过程中,是否存在其他外周途径影响其免疫行为也同样值得探索。
2.2.2 MLV介导免疫细胞的迁移
过去观点认为,BBB 的存在使大脑处于“免疫豁免状态”,但随着MLV的发现,揭示了脑实质内的小胶质细胞,还包括T 细胞、巨噬细胞等免疫细胞(其中部分通过MLV引流),动态监视着大脑的健康状况,即MLV存在运输大脑免疫细胞的功能。研究发现,CSF 中的T 细胞主要是CD4+/CD45RA-/CD27+/CD69+记忆T 细胞,这些细胞表达高浓度的CC-趋化因子受体7(CC-chemokine receptor 7,CCR7)和L 蛋白选择素,负责生理状态下CSF 和血管周围区域的免疫监视,并保留参与自身免疫反应和回流入二级淋巴器官的功能,即如果这些淋巴细胞在监视过程中未发现抗原,就会通过MLV 回流入dCLN[15-16]。健康状态下,T 细胞由血液循环进入脑膜的途径有3 条:软脑膜表面血管、硬脑膜上的脑膜血管或CP[17]。
淋巴内皮细胞已被证明在炎症环境中会增殖和扩张。Louveau等[18]使用多发性硬化动物模型——实验性自身免疫性脑脊髓炎(experimental autoimmune encephalomyelitis,EAE)诱导中枢神经系统的强烈炎症。研究表明,在疾病晚期,位于筛板中枢侧的淋巴管扩张,且MLV 内部和周围的T 细胞密度增加,这意味着引流淋巴结的脑膜免疫细胞输出增加。通过手术结扎dCLN的传入淋巴管会延迟EAE的进展。此外,与野生型小鼠相比,淋巴管完整性缺陷型Prox1het小鼠内源性脑膜T细胞的数量显著增加。注射入2 月龄Prox1het 小鼠脑脊液中的T 细胞,不能像野生型小鼠那样有效地引流到dCLN中。与Proxy1het 小鼠相似,dCLN 的传入淋巴管结扎后,注射到小鼠脑脊液的T 细胞、DC 未能流入dCLN。这些结果表明,脑膜对T细胞的引流依赖于淋巴管的完整性。
实验课是科学探究的一种方式,在上实验课之前,应该先由教师检验实验成果,与此同时,必须直接控制实验室的设备状态。科学研究是学生学习知识的重要方式,实验课可以引导学生主动探索,亲身经历实务研究发展的过程,有利于保护学生的好奇心和激发学生学习科学知识的主动性。实验课还有许多要求,第一,教师应该告诉学生课程的安排,保证实验室的安全系统,要求学生们必须在实验前做好准备。第二,教师应该明确学生们的任务要求和课程安排,如果没能在规定的时间内完成任务要求,也不要插手,让学生们学会独立思考解决问题。
从分布来说,MLV 可能起始于眼部,行于嗅球上方,之后与上矢状窦并行。与膈肌的淋巴管相比,MLV相对细小,形成相对简单的网络,覆盖相对少的组织[11]。MLV 的存在揭示了脑内液体在中枢神经系统内存在定向运输,一些证据表明CSF 内的示踪剂可以穿过蛛网膜进入硬脑膜淋巴系统,也有观点认为示踪剂并非由外周途径而是从颅骨上的小孔流出[5]。近期研究表明,MLV 携带大量免疫细胞,将脑实质有关的细胞因子与外周免疫系统相联系,并由此将脑实质与CSF 中免疫细胞及细胞因子从蛛网膜下隙引流入颈深淋巴节(deep cervical lymph node,dCLN),部分进入颈浅淋巴节(superficial cervical lymph node,sCLN)[11-12]。
3 脑膜免疫与中枢神经系统
3.1 脑膜:有效的免疫屏障
脑膜作为中枢神经系统的“守门员”,不仅参与大脑的基本运转,在免疫监视中也具有举足轻重的作用,能够有效阻止病原体进入脑实质[5]。Van Hove 等[19]通过解剖脑边缘区域以及RNA 测序和高维细胞计数法,证实了脑膜在生理状态下以巨噬细胞为代表的多样性与动态变化,并发现相比其他中枢神经系统免疫屏障,脑膜相关DC、T细胞、B细胞、NK 细胞等免疫细胞的数量更为庞大。由此可见,脑膜免疫对中枢神经系统的免疫调控具有重要作用。
3.2 脑膜免疫相关细胞因子
细胞因子是免疫细胞产生的最重要的信使分子,通过自分泌或旁分泌机制调节许多生物功能。多项研究表明,脑膜免疫可以通过分泌神经调节细胞因子控制神经信号,进而影响机体行为和认知[10]。除免疫细胞释放的炎症因子产生炎症环境增加中枢神经系统灰质的脱髓鞘和神经退行性疾病风险之外,也有研究发现脑膜免疫细胞来源的某些细胞因子可促进学习和记忆行为,证实了脑膜免疫相关细胞因子对中枢神经系统功能存在正反两面的影响[20]。
综上所述,当前随着社会的不断发展,对于我国社会生态环境带来了一系列的影响,甚至出现了辐射污染的现象。辐射环境监测作为辐射环境评价的基础,同时也起到了良好的监督作用,我们要充分的意识到辐射环境监测对于辐射环境评价的重要性所在,不断强化辐射环境监测工作,进而为接下来辐射环境评价工作提供有利的数据参考,切实提高辐射环境监测质量,促进我国辐射环境监测的可持续发展。
研究表明脑膜T 细胞分泌的白细胞介素(interleukin,IL)-4 在认知功能方面发挥重要作用[21]。例如,IL-4缺失的小鼠表现出学习障碍,通过腹腔注射富含IL-4 的T 细胞可逆转上述病理。现已明确CD4+αβT细胞可释放IL-4和IL-13,拮抗促炎细胞因子对AC的有害作用,促进AC表达脑源性神经营养因子(brain-derived neurotrophic factor,BDNF)、胰岛素样生长因子(insulin like growth factor,IGF)-1和转化生长因子(transforming growth factor,TGF)-β等,从而影响神经元活性[21]。在稳态条件下,CD4+γδ T细胞是脑膜中IL-17 的主要来源,它们通过增加海马神经元的谷氨酸突触可塑性来促进小鼠的短期记忆[22]。此外,Filiano 等[23]发现健康脑膜中T 细胞倾向于产生大量γ干扰素(interferon γ,IFN-γ),缺乏IFN-γ的小鼠表现出社交缺陷。大脑抑制性神经元直接对脑膜T细胞分泌的IFN-γ产生应答,引起γ-氨基丁酸能神经元传递增强,参与小鼠社交行为[23]。
(4)俄罗斯在实施创新发展战略和发展数字经济时,相当重视人工智能基础科学的研究。我国在发展人工智能过程中,除了加大与俄罗斯在立法领域的交流和借鉴外还需广泛地开展基础研究领域的合作,通过建立联合实验室等做法,引进高级专家,培养和锻炼自己队伍,以促进我国人工智能关键技术的研发。■
最近的文献也证明了其他细胞因子信号对神经元的作用。研究发现小鼠脑膜γδ T细胞表达趋化因子受体6(chemokine receptor 6,CCR6)及IL-17a,后者调控小鼠焦虑样行为,有利于群居共生[24]。Shi等[25]通过单细胞RNA测序和流式细胞技术,发现小鼠中风后1~5 周内调节性T 细胞开始浸润大脑,并通过其衍生的骨桥蛋白(osteopontin,OPN)增强小胶质细胞的修复活性,促进少突胶质细胞的再生以及白质的修复,并分泌IL-2 免疫复合体,有助于维护白质以及中枢神经系统的功能,从而证实脑膜调节性T 细胞来源的细胞因子有助于维护脑稳态。MC分泌的细胞因子也被证明参与大脑免疫,其分泌的IL-6、IL-1β、TNF-α等细胞因子参与了神经再生以及脑膜固有细胞的免疫行为[22]。Chikahisa 等[26]通过研究MC缺陷的小鼠模型,发现MC释放的组胺促进觉醒,参与调节昼夜节律。
以上研究表明,学习、记忆、空间认知以及社交行为等中枢神经系统功能的维持依赖正常免疫相关细胞因子水平维持的脑膜免疫稳态。而疾病状态下这种稳态的破坏是各种脑功能行为异常的重要原因。未来,更多的脑膜免疫相关细胞因子和它们所产生的效应及具体机制都值得深入探究。
3.3 脑膜介导中枢与外周免疫信息交互
脑膜作为维护中枢神经系统免疫稳态的重要屏障,在免疫监视大脑内部稳态的同时,还参与介导中枢与外周免疫信息的交互,通过外周免疫细胞的迁移或感知外周免疫信息等途径维护大脑稳态。
MS 作为最常见的慢性中枢神经系统炎症性脱髓鞘疾病,多发于年轻女性,部分患者可能出现认知功能障碍,且症状随时间推移逐渐加重[32]。Choi等[33]通过免疫组化分析MS 患者死后的脑组织,发现随着脑膜炎症细胞的浸润,脱髓鞘现象更为广泛,大脑灰质的轴突丢失也随之发生,脑膜炎症的严重性与MS 病情严重程度和死亡率呈正相关。Lucchinetti 等[34]通过活体脑组织成像技术,发现脑膜炎症常常发生在MS 早期,甚至可能早于白质病变的出现。在MS病程中,大脑活性T细胞入侵中枢神经系统,诱导自我损伤性炎症反应发生,T细胞浸润最早在脑膜中被观察到,早于脑实质浸润以及临床症状的出现,而在CSF中也发现了T细胞浸润[35]。由此可见,脑膜炎症与MS的发病和进展紧密相关。
此外,也有团队将放射性同位素注入CSF后,发现示踪剂主要富集在大鼠嗅鼻甲处,在3月龄呈高速流动状态,而在12月龄和15月龄明显下降,由此可见,大量的CSF 通过筛孔进入嗅淋巴管的运输也代表了CSF淋巴运输的范式,并具有实际优势,而其引流效率与年龄呈负相关[13]。Kwon 等[14]将吲哚青绿(indocyanine green,ICG)注入AD小鼠和野生型小鼠蛛网膜下腔,并用近红外荧光成像技术对比二者的CSF 引流情况,发现ICG 立即进入野生型小鼠的蛛网膜下腔并通过筛血管引流至下颌下淋巴结,而AD 小鼠该淋巴结内荧光强度显著降低,且外周淋巴管收缩频率降低,说明AD 小鼠的CSF 流出途径严重受损,CSF清除障碍,考虑与外周淋巴管功能受损相关,这证实鼻腔嗅觉系统淋巴管流入外周淋巴管的功能下降同样导致淋巴管堵塞,从而限制CSF的流出。可见,两种淋巴途径均对CSF 存在引流作用,但二者的权重,特别是MLV 引流的具体机制尚待探究。
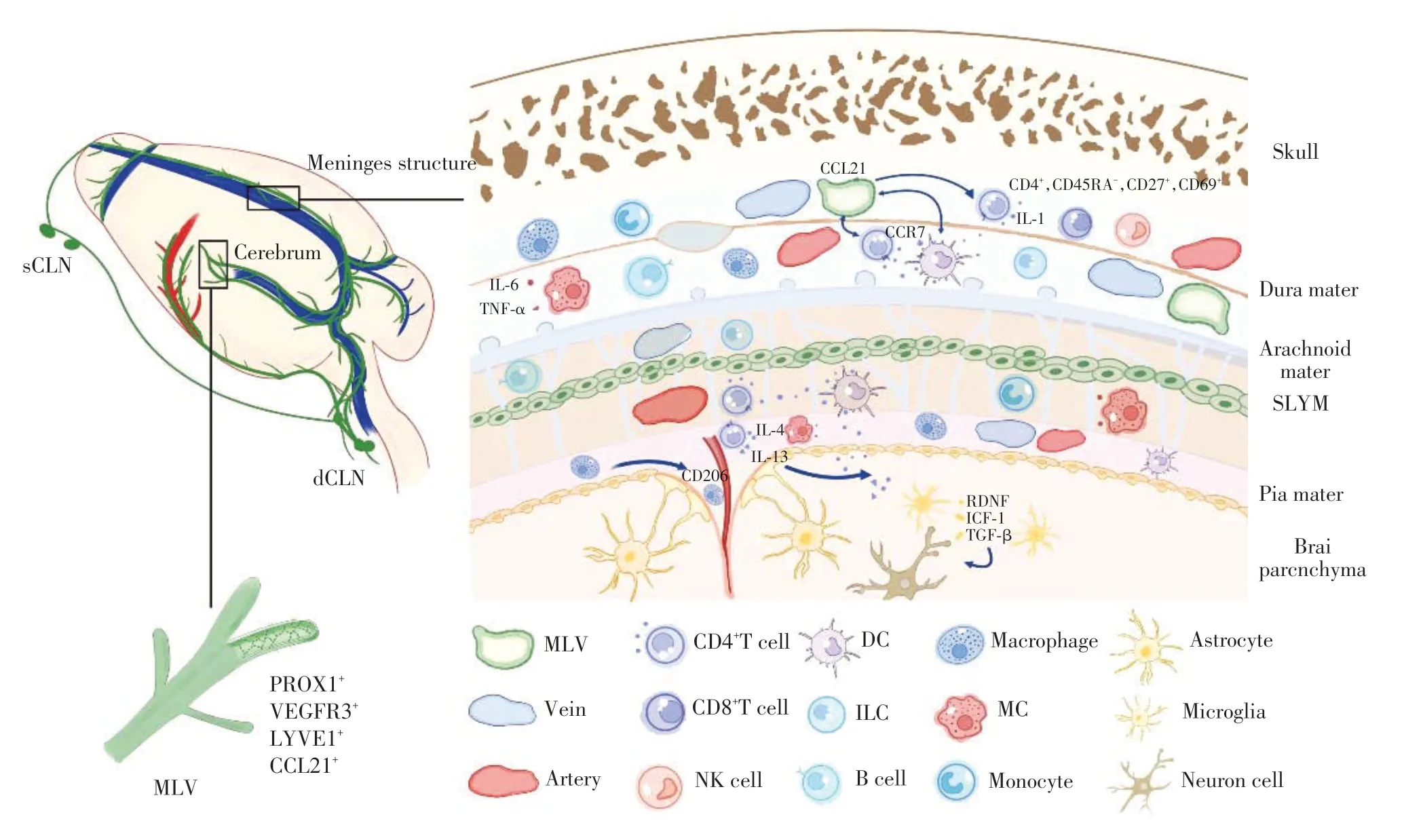
图1 脑膜结构和免疫功能相关示意图Figure 1 Structure and immune function of meninges
4 脑膜免疫与中枢神经系统疾病
4.1 多发性硬化症(multiple sclerosis,MS)
近年来,越来越多的研究发现胃肠道与中枢神经系统之间存在特殊的联系,彼此双向影响,此系统被命名为“脑-肠轴”[27]。Fitzpatrick等[28]发现在生理状态下的小鼠和人脑膜中有一种IgA 分泌型浆细胞,其毗邻硬脑膜静脉窦,并被证明来源于肠道,这种肠道来源的浆细胞有助于维护静脉窦稳态,证实了来自外周的免疫细胞影响脑膜的体液免疫。Benakis等[29]通过建立小鼠脑卒中模型,发现脑卒中后调节性T 细胞会从肠道迁移至软脑膜,并且大脑神经保护依赖于肠道相关的IL-10 和IL-17,证明外周免疫细胞也参与脑膜的细胞免疫过程。另外,肠道微生物群的代谢产物及病原体相关分子模式也被证明对脑膜免疫细胞存在影响[30]。研究还发现,将帕金森病患者的肠道微生物群移植入α突触核蛋白转基因鼠,加剧了该类小鼠神经变性进程,这可能与肠道微生物群衍生的短链脂肪酸相关[31]。脑-肠轴作为大脑与外周免疫系统信息交互的一种渠道,值得进一步深入探究。
由于MS 患者标本不易获取,最理想的研究模型是EAE 模型鼠,目前主要通过自身反应性T 细胞过继转移诱导被动性EAE模型鼠,或通过髓磷脂抗原免疫注射诱导主动性EAE 模型鼠[5]。有研究在EAE 鼠的病理过程中观察到,有活性的T 细胞进入脑实质,而无活性的T细胞则进入CSF,经过软脑膜的筛选,T 细胞得以接触抗原,引起脑组织损伤,诱导MS 的病理过程[36]。Furtado 等[37]发现,在EAE 病理进程中,T细胞活化最早发生在颈淋巴结,手术切除颈淋巴结可以改善EAE的病理情况,进一步证实了EAE的发生与脑膜T细胞活化相关。Louveau 等[18]研究表明,在EAE 活跃期,脑膜中CD4+T 细胞的浸润数量高于脑实质,破坏MLV有助于延缓EAE的病理进程,改善EAE模型鼠的症状。以上发现不仅佐证了在中枢神经系统炎症过程中脑内浸润的T细胞从MLV 引流入dCLN 这一免疫活化途经,也为临床治疗MS提供了新的思路。
除T 细胞外,MC也被认为参与了EAE 的进程。文献报道称,位于硬脑膜和软脑膜的MC通过释放TNF-α诱发嗜中性粒细胞进入脑膜,加剧中枢神经系统的炎症细胞浸润,使EAE 病情恶化。并且,脑膜中MC的活化以及其相关嗜中性粒细胞浸润被认为发生在EAE 早期,甚至早于临床症状的出现,于是研究者猜测,此过程可能损害了BBB,导致大量免疫细胞浸润中枢神经系统,从而引发炎症反应[38]。Russi等[38]通过对比野生型小鼠和遗传性MC缺乏小鼠,发现后者的脑膜不会出现自身反应性T细胞聚集的现象,证明MC具有招募T细胞在脑膜聚集的功能,不仅如此,MC似乎还会增强自身反应性T细胞的免疫应答。
由此可见,脑膜和其免疫细胞及细胞因子在MS的发病和进程中发挥有效的屏障作用,可作为疾病干预的有效靶点。
4.2 神经退行性疾病
许多与年龄相关的神经退行性疾病的发生和发展与脑内代谢废物的积聚有关。MLV具有清除脑内代谢物、引流免疫细胞以及吞噬细胞的功能,对治疗这些疾病有积极价值。
AD 是一种最常见的以认知功能障碍为特征的神经退行性疾病,目前认为其神经病理改变包括记忆相关脑区Aβ沉积导致氨基酸肽结构的变化从而形成胞外原纤维、Tau 蛋白过度磷酸化所致的神经原纤维缠结、神经元与突触丢失等[39-40]。有研究者认为慢性神经炎症抑制了神经元功能,导致上述相关病理变化[41]。而神经炎症发生的中心——脑膜,被认为是AD病理变化的重要因素,脑膜受损,脑膜相关免疫细胞群受到刺激慢性活化,周围免疫细胞在BBB中聚集,导致炎症和神经毒性因子释放,结合血管旁CSF和ISF内大分子引流与物质交换减慢,从而导致一系列功能和认知损害[39,42]。研究发现通过破坏转基因鼠的MLV,发现其脑膜内Aβ沉积增加,而增强老龄AD模型小鼠的MLV功能可以改善其认知功能障碍,证明了AD病理变化与脑膜转运功能降低相关[42]。Da Mesquita 等[43]通过阻断AD 小鼠的MLV,也发现Aβ沉积和小胶质细胞的炎症应答等AD病理改变加剧,抗Aβ被动免疫疗法疗效减弱。
此外,多项研究表明,颅内压增高可通过诱导血管闭塞、降低脑血管搏动等途径抑制胶质淋巴系统循环,可能引起dCLN引流受损和MLV功能障碍[56]。由此可见,靶向MLV的清除功能及其特定的免疫细胞可以改善各类脑卒中的病程及预后,成为疾病干预的潜在靶点。
脑膜的原发性恶性肿瘤极为罕见,最常见的是继发性脑膜癌。原发性中枢神经系统肿瘤,包括儿童的髓母细胞瘤和室管膜瘤,以及成人的多形性胶质母细胞瘤等,通常转移到脑膜。脑膜可能成为脑肿瘤发生、转移的渠道。胶质母细胞瘤是一种恶性原发性脑肿瘤,平均总生存期<15个月,临床报道表明复发性胶质母细胞瘤可通过颈部淋巴结转移[57]。此外,动物实验表明,在患有胶质瘤的小鼠中,由MLV 排出的CSF 显著减少[58]。注射胶质瘤和黑素瘤细胞的小鼠背侧MLV的直径和密度明显增加,提示脑瘤可促进MLV 生成。同时,MLV 是DC 等免疫细胞向dCLN迁移和建立对脑肿瘤免疫应答的主要途径[59]。Visudyne介导的MLV消融削弱了抗PD-1/CTLA4免疫治疗的疗效,而肿瘤细胞中过表达的血管内皮细胞生长因子-C(vascular endothelial growth factor-C,VEGF-C)则以CCL21/CCR7 信号依赖的方式发挥相反的作用,VEGF-C 诱导MLV 生成,导致抗肿瘤环境的产生,促进T 细胞的浸润、启动和招募[59]。由此可见,MLV通过运输不同的免疫组分在肿瘤脑内外转移的过程中发挥复杂的作用。根据不同的抗肿瘤原理结合MLV 的调控可以起到更好的抗肿瘤作用。
4.3 脑卒中
脑卒中可被分为两类:血管破裂引起的出血性脑卒中[如蛛网膜下腔出血(subarachnoid hemorrhage,SAH)、脑实质出血(intracerebral hemorrhage,ICH)等[46]]和血管阻塞引起的缺血性脑卒中[47]。尽管脑卒中主要造成脑实质损害,但脑膜的细胞活化和免疫应答往往早于脑实质损伤[29]。
在小鼠急性SAH模型中,dCLN引流减少,脑组织和脑膜中大分子和免疫细胞异常聚集,提示SAH可损害或阻碍MLV的功能,从而可能导致并加重疾病[48]。SAH发生后,纤维蛋白原和纤维蛋白在血管周围间隙沉积,导致胶质淋巴系统的功能失调,出入脑内的CSF和ISF流量均显著下降,从而引起血管炎、广泛微梗死和神经炎症[49]。通过阻断SAH小鼠模型的MLV,观察到引流至dCLN的红细胞显著减少,随之SAH引发的神经炎症及神经损害程度显著升高,提示SAH后MLV清除外渗红细胞的能力减弱,增加此清除功能可能有助于减轻SAH引发的脑部损害[48]。
C语言属于一种高级的计算机编程语言,通过利用C语言可以编写计算机应用程序,早期的应用软件很多都是通过C语言完成开发的。包括目前的IT开发领域,很多都是以C语言为基础发展而来的高级语言。该门课程主要阐明了基本的编程思想、方法和理念,是学生学习其他高级语言的基础。C语言教学过程非常注重对实践能力的锻炼和提升。学生要将自己编写的代码输入计算机,由计算机来判断代码的准确性。
缺血性脑卒中时,BBB 的破坏会导致白细胞,尤其是中性粒细胞和巨噬细胞在脑实质内和MLV中聚集[50]。大脑产生的抗原(微管相关蛋白2 和髓鞘碱性蛋白)在淋巴结巨噬细胞中富集,这些抗原可激活抗原递呈细胞DC;脑脊液通过脑膜将抗原、T 细胞和其他免疫细胞引流到dCLN[51]。DC通过上调趋化因子受体CCR7,与MLV 表达的配体CCL21结合,迁移到dCLN。研究表明,短暂性大脑中动脉闭塞(transient middle cerebral artery occlusion,tMCAO)诱发卒中后,结扎dCLN 可增加脑水肿和梗死面积[52]。缺血性脑卒中发生后,T细胞可通过BBB、CP和脑膜3 个途径浸润脑实质[53]。基于“脑-肠轴”理论,有研究发现效应T 细胞在缺血后首先从肠道进入软脑膜,通过分泌IL-17加剧炎性反应,导致脑实质的趋化因子数量增多,从而引起包括中性粒细胞、单核细胞在内的细胞毒性免疫细胞群的MLV内浸润[53]。另外,有学者发现,在缺血早期,脑膜中IL-17+γδ T细胞数量明显增多,甚至早于其在缺血区的聚集,这可能与脑损伤相关趋化因子基因CXCL1/CXCL2的上调有关[54]。通过阻断内皮细胞VEGFR3的表达,缺血区附近的淋巴结所释放的趋化因子及细胞因子减少,促炎性巨噬细胞的增殖被抑制,从而有效缩小脑梗死区的面积[55]。即脑膜在缺血性脑卒中炎症发生中起到守卫关卡的作用。
在脑膜相关固有免疫细胞方面,鉴于固有小胶质细胞与单核巨噬细胞均可吞噬、消化Aβ,而炎症时调控单核巨噬细胞进入脑内的趋化因子受体CCR2 缺乏可降低下游小胶质细胞聚集,清除代谢物功能减弱,进而增加Aβ沉积,由此可推测通过调控上游趋化因子来募集小胶质细胞,促进其脑内聚集有助于延缓AD病理进程,可成为治疗AD的新思路[44]。除了借助脑膜免疫渠道治疗AD,昼夜节律以及脑膜与外周的信息交互等角度也有助于探索AD治疗的新策略,Oxana等[45]通过对比清醒与睡眠期的小鼠,发现生理性睡眠过程增强MLV清除脑内代谢物的功能,另外,睡眠中应用光生物调节法(photobiomodulation,PBM)可以显著增加Aβ清除率,对AD起到治疗作用。
4.4 肿瘤
由此可见,在临床治疗AD的过程中,基础免疫治疗辅以改善MLV 功能的疗法也许能够获得更好的疗效。
“绿色”,主要指绿叶蔬菜和瓜果。如绿色蔬菜主要包括:芹菜、油菜、菠菜、韭菜、绿辣椒、空心菜、雪里蕻等。每人每天至少吃250 g蔬菜和水果,它能提供人体所需的维生素和纤维素以及人称第八营养要素的黄酮类,各种绿叶菜以色深为佳。此外,绿豆、茶叶也不可少;
4.5 脑膜与其他中枢神经系统疾病
随着科学研究的深入,其他中枢神经系统疾病的病理机制与脑膜的联系正在被逐渐挖掘。通过观察小鼠神经元,发现来源于硬脑膜的γδ17 T 细胞通过IL-17 途径调控焦虑样表现,这为临床探索焦虑症的病理机制与治疗方向提供了思路[24]。另外,有学者通过脂多糖(lipopolysaccharide,LPS)诱导幼龄小鼠炎性反应,证明早期炎性反应通过
猜你喜欢
天津医科大学学报(2019年3期)2019-08-13
中国临床医学影像杂志(2019年4期)2019-06-18
国际呼吸杂志(2019年4期)2019-03-12
兽医导刊(2016年6期)2016-05-17
中华老年多器官疾病杂志(2016年7期)2016-04-28
中国卫生标准管理(2015年18期)2016-01-20
转化医学电子杂志(2015年4期)2015-12-27
中国当代医药(2015年16期)2015-03-01
湖北科技学院学报(医学版)(2012年4期)2012-08-15
组织工程与重建外科杂志(2011年2期)2011-03-27
