
采煤沉陷区复垦对土壤细菌群落组装及固碳功能的影响-以东滩矿区为例
2024-03-18 09:20宋晓君董文雪朱燕峰尤云楠
煤炭科学技术 2024年1期
陈 浮 ,宋晓君 ,董文雪 ,朱燕峰 ,尤云楠 ,马 静,
(1.河海大学 公共管理学院, 江苏 南京 211000;2.中国矿业大学 化工学院, 江苏 徐州 221000;3.中国矿业大学 矿山生态修复教育部工程研究中心, 江苏 徐州 221116)
0 引 言
东部平原煤矿区井工开采对地表生态系统破坏极为严重,地表沉陷积水,土壤潴育化,陆生植被淹死,不利于生态系统功能维持和生物多样性保护[1-2]。恢复土壤功能是东部平原矿区复垦的核心任务,并最终形成健康、肥沃的土壤生产力,维持农田生态系统的可持续发展[3-4]。然而,复垦土壤功能系统发育和恢复是一个漫长的过程,它取决于恢复微生物群落内不同组分间功能关联所需的时间[5]。微生物是生态系统分布最广、又最活跃的组分,驱动地球生物化学循环、能量代谢和土壤-植物相互作用等过程,是生态系统重要过程的调解者和催化剂[6-7]。因此,厘清复垦土壤微生物群落系统发育对认识矿山生态修复中微生物功能和作用至关重要[8]。
近年来,国内外学者非常重视采矿扰动和土地复垦下土壤生态系统演化[9],微生物群落组成与结构、功能差异和响应模式等一直备受关注[10]。这些研究多集中于不同复垦年份、不同植被类型和复垦技术下土壤微生物群落结构、物种和功能多样性变化及其影响因素的考察。例如,采矿污染导致固碳和硫酸盐还原相关的功能基因丰度降低,而反硝化相关的功能基因丰度得到提升[11-12]。此外,微生物相互作用(如共生或竞争)可能会改变微生物群落结构以适应这些不利的环境,同时这些变化可直接影响群落稳定性[13]。解开矿区微生物群落组装过程对于理解微生物如何应对和适应采矿环境的演替至关重要。微生物的组装过程,即确定性过程(环境选择)和随机过程(如扩散和漂移)已被用于阐明各种生态系统中的微生物群落组装,如农业土壤,河水和水库沉积物[14-15]。随机漂移和扩散过程被发现是未受干扰的森林土壤中微生物群落组装的关键驱动因素。在重金属污染的农业生态系统中,确定性过程主导着微生物群落的组装,且其影响随着重金属浓度的增加而增加[16]。然而,矿区复垦影响细菌群落组装及机制方面仍存在认知缺口,且群落组装过程如何重塑细菌群落的结构分布和功能尚未研究清楚,不利于充分理解功能性微生物的生态组合及其对矿区生态修复的贡献。东部采煤沉陷区土地复垦方向主要为恢复耕作,但复垦土壤结构差、肥力相对低下,且先前研究集中于监测复垦土壤理化动态和微生物群落结构与组成,对微生物群落组装及固碳功能极少关注。因此,探明东部平原矿区复垦土壤微生物群落组装机制及其生态响应对于采矿生态系统的管理和恢复至关重要。
那么,东部平原矿区复垦及时长究竟如何影响微生物群落结构变化?复垦土壤微生物群落组装到底受确定性过程控制,还是随机性过程主导?复垦如何影响土壤碳固持?为此,选择山东省邹城市东滩煤矿沉陷复垦区为研究对象,采集9 a、12 a、15 a 和18 a 4 个复垦年限和1 个对照样共65 个土样,利用高通量测序和零模型分析方法,探索复垦及时长对土壤微生物群落结构、构建机制及固碳基因丰度的影响,为东部平原矿区沉陷复垦土壤发育和生态恢复提供新见解。
1 材料与方法
1.1 研究区概况
东滩煤矿沉陷复垦区位于山东省邹城市(35°25'52''N、116°52'39''E),属于暖温带季风气候区,年降水量752.9 mm,年均气温14.1 ℃。该区为典型的山前冲积平原,土壤类型为黄潮亚土,砂粒(>0.02 mm)、粉 粒(0.002~0.02 mm)和 黏 粒(<0.002 mm)分别占22.3%、65.9%和11.8%。2002—2011 年,分4 次采用表土剥离、煤矸石充填(200~400 cm),再覆盖表土(60~80 cm)和机械压实,每次客土来源一致。复垦后土地由矿山企业的农业开发公司统一种植小麦-大豆,一年两熟。小麦单季施底肥600 kg/hm2(常规复合肥),拨节时施氮肥150 kg/hm2,机械收割时一般留茬高15~20 cm。大豆单季施底肥450 kg/hm2(常规复合肥),机械收割时自动粉碎还田。除复垦时间不同外,研究区地形、气候、水文、土壤母质、种植模式和田间管理等完全一致,构成一个完善的“空间换时间”复垦序列。
1.2 样品采集与测试
2020 年8 月22 日,采用随机五点采样法从复垦9 a、12 a、15 a 和18 a 样地分别采集13 个表土(0~20 cm)混合样,分别记为R9、R12、R15 和R18。并选取附近未受采矿影响、由农业开发公司统一种植农田随机收集13 个表土样作为对照(CK),共采集土样65 个(图1)。样品采集后现场剔除根系、残体和砾石等,充分混匀后分为2 份。一份通过风干、研磨后过2 mm 筛,用于测定土壤理化性质和部分土壤酶活性;另一份鲜土样无菌密封储存于-20 ℃冰箱中,用于微生物相关信息测定。土壤pH 是通过测定25 mL 水掺1 g 土的水悬液获得。易氧化有机碳(EOOC)通过高锰酸钾氧化法测定。有机碳(SOC)采用重铬酸钾氧化法和硫酸铁铵滴定法测定。用碳酸氢铵萃取后用钼锑钪比色法测定土壤有效磷(AP)。通过凯尔文蒸馏滴定法测定土壤全氮(TN),用氯化钙浸提后用分光光度法测定硝态氮(AN)。过氧化氢酶(CAT)酶活采用高锰酸钾滴定法测定,碱性磷酸酶(PO)采用磷酸苯二钠比色法测定,多酚氧化酶(PPO)酶活采用邻苯三酚比色法测定,β-葡萄糖苷酶(BG)酶活采用硝基酚比色法测定,脲酶(UE)酶活采用苯酚钠-次氯酸钠比色法测定,亮氨酸氨基肽酶(LAP)酶活采用S-LAP 试剂盒测定,脱氢酶(DH)酶活采用氯化三苯基四氮唑法测定,蛋白酶(PRO)酶活采用加勒斯江法测定,FDA 水解酶(FDA)酶活通过荧光素比色法测定[17-18]。

图1 研究区位置和采样区分布Fig.1 Location of study area and sampling area distribution
1.3 微生物信息测定
根据E.Z.N.A.®soil 试剂盒(Omega Bio-tek,Norcross,GA,U.S.)说明书进行土壤总DNA 提取,使用微量紫外分光光度计NanoDrop one 测定所得DNA浓度和纯度,利用1%琼脂糖凝胶电泳检测DNA 提取质量。利用细菌通用引物515F(5′-GTGCCAGCCGGTAA-3′)和907R(CCGTCAATTCMTTRAGTT)对细菌16S rRNA 的V4 和V5 区进行PCR 扩增,扩增产物等物质的量比混匀后,利用NEBNext® UltraTM II DNA Library Prep Kit 制备测序文库。构建好的文库经Qubit 和Q-PCR 定量确认文库合格后,使 用Illumina Nova 6000 平 台 对 扩 增 子 文 库 进 行PE250 测序。
土壤碳循环功能基因丰度采用QMEC(Quantitative Microbial Element Cycling)基因芯片进行定量化测定。具体方法如下:按照MagaBio 土壤DNA 提取试剂盒方法,取0.25 g 土壤样品,提取土壤DNA,进行总量及纯度检测。检测合格后分别将样品板和引物板试剂添加至高通量qPCR 芯片SmartChip MyDesign Chip( Takara Biomendical Technology,Clontech)微孔中,在SmartChip 实时PCR 系统中执行qPCR 反应及荧光信号检测,自动生成扩增曲线和溶解曲线。使用Canco 软件获得各基因在样本中的检出情况和扩增循环数(Ct 值),并以16SrRNA 作为内参数据进行标准化获得各基因相对定量信息。再根据Roche 检测16SrRNA 基因相对定量信息,最终换算其他基因的绝对定量信息。
1.4 微生物群落构建机制分析
利用R studio 中的“picante”包计算平均最近物种距离(MNTD),β-平均最近物种距离(βMNTD)和β-最近物种指数(βNTI)等参数来评估样品间系统发育。当βNTI 值大于2 或者小于-2,表示群落组装过程为确定性过程(分别表示异质性选择过程和同质性选择过程)。相反,当|βNTI|值小于2,群落组装则以随机性过程为主。为进一步确定随机性过程的分类,利用“vegan”包计算了基于群落的Bray-Curtis 矩阵的 Raup-Curtis 矩 阵( Raup-Curtis matrix, 简 称RCbray)。RCbray 值 大 于+0.95、|RCbray|值 小 于0.95 和RCbray 值小于-0.95 分别代表扩散限制、漂移和同质性扩散[19]。
1.5 数据处理与统计分析
利用美格基因云平台进行微生物信息分析。土壤理化性状和酶活性采用SPSS 26.0 软件(IBM,美国)进行ANOVA 分析,使用mantel test 检验评估物理化学因素对细菌群落结构的影响。采用R studio(MathSoft,美国)构建中性群落模型和随机森林模型[20],利用ggplot2 包进行柱形图绘制。利用Amos 21.0 软件拟合结构方程模型(Structural Equation Model,SEM)分析土壤理化性质、酶活性以及群落组装生态过程和碳循环功能基因丰度的关系。其中,对土壤碳循环功能基因丰度进行归一化处理[21],划分为碳降解、碳固定和甲烷代谢3 大功能类群,具体公式如下:
式中:xi为样本单个基因丰度;x'为碳降解、碳固定和甲烷代谢微生物功能类群归一化丰度;i、n分别为样本数量和基因数量。
2 结果与分析
2.1 不同复垦年限下土壤理化性状和酶活性变化特征
图2 显示了不同复垦年限下土壤理化性质和酶活性变化。复垦土壤偏弱碱性,pH 随复垦时长先升后降,复垦处理与CK 呈极显著差异(P<0.001)。土壤有机碳、有效磷及硝态氮含量随复垦年限表现为下降趋势,且复垦土壤SOC 含量均低于CK。铵态氮与复垦时长呈正相关,R12 中AN 含量已略高于CK。复垦时长对CAT 影响显著,与PO 一样随复垦时长持续增加,但复垦处理PO 活性仍显著低于对照(P<0.001)。PPO 活性先升后降,逐渐与CK 持平。复垦处理LAP 均高于CK,且与CK 呈极显著差异(P<0.001)。UE、BG 和PRO 活性与复垦时长呈负相关关系,但R18 中UE 活性仍极显著高于CK(P<0.001)。综上,与CK 比,随复垦年限增加,土壤酶活性总体呈上升趋势(PO、DH、PRO 除外);土壤有机碳、有效磷及硝态氮含量则呈下降趋势。

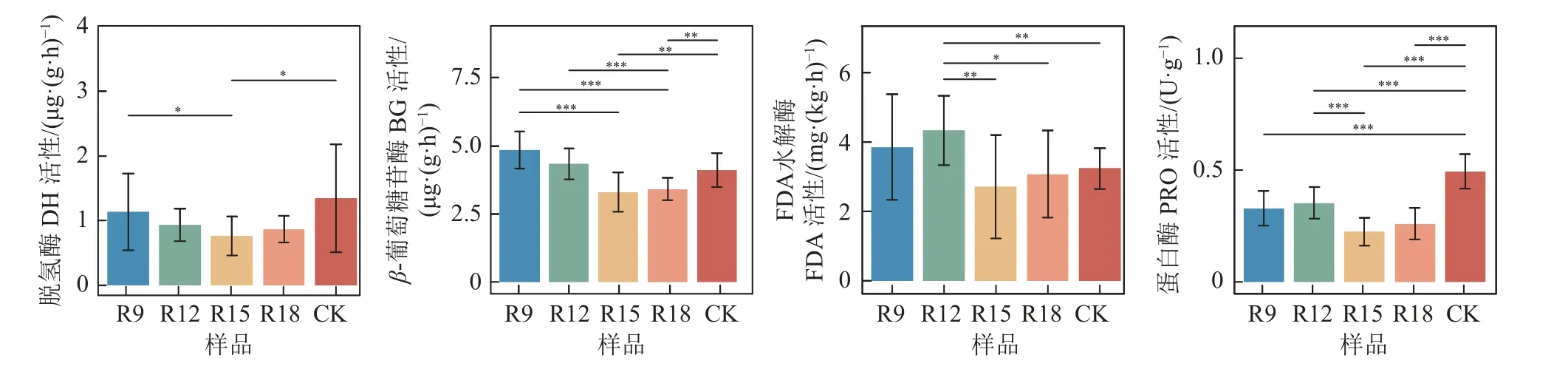
图2 不同复垦年限下土壤理化性状和酶活性变化Fig.2 Changes of soil physicochemical properties and enzyme activities under different reclamation years
2.2 不同复垦年限下土壤细菌群落构建过程
根据零模型,|βNTI|<2 被划定为土壤微生物群落构建的随机性过程。本研究中,不同复垦处理组|βNTI|<2 的数据约占90%,故不同复垦时间土壤细菌群落构建以随机性过程为主导(图3)。为进一步探究群落组装机制,根据RCbray 值细分显示,随机过程中扩散限制对群落组装的贡献最大,并随复垦时长增加而扩大。NCM 模型中R2值越高拟合效果越好,越接近中性群落模型,即群落构建受随机性过程的影响越大,受确定性过程的影响越小。本研究评估结果表明土壤细菌群落随机过程的相对贡献随复垦时长增加先升后降,解释方差均高于80.0%,反映OTU 频率及其相对丰度之间关系拟合程度较好,说明复垦土壤细菌群落构建受随机性过程影响大(图4,图中Xr,mean为平均相对丰度),这与零模型分析所得结果一致。
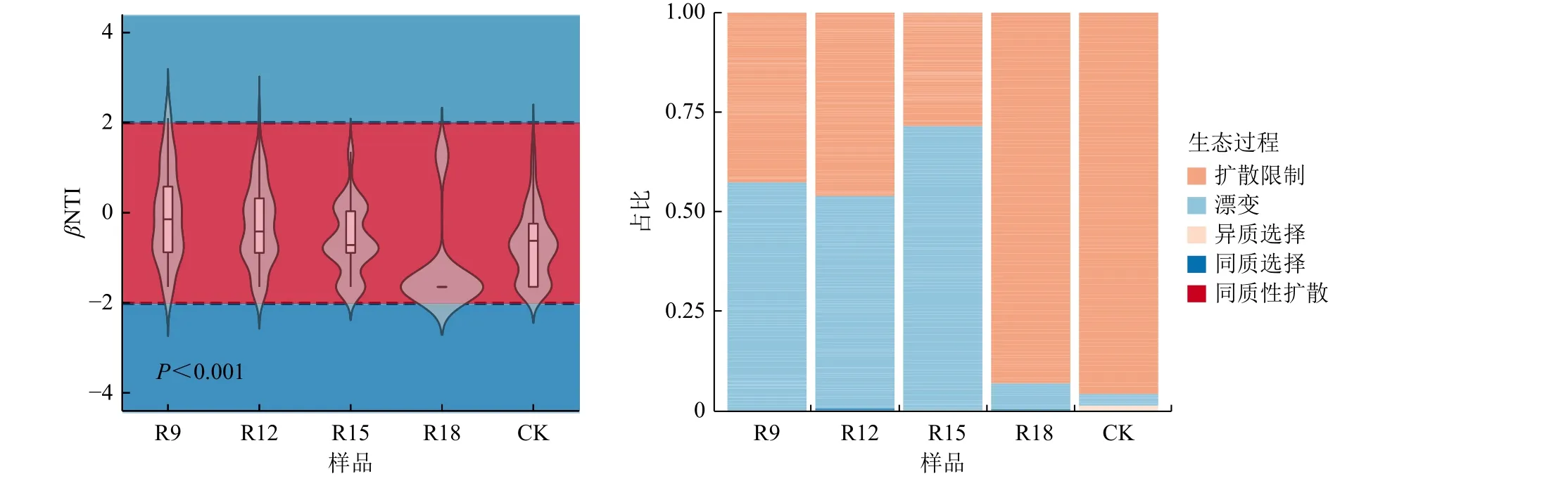
图3 不同复垦年限下细菌群落βNTI 值和组装过程Fig.3 βNTI and assembly process of bacterial community under different reclamation years
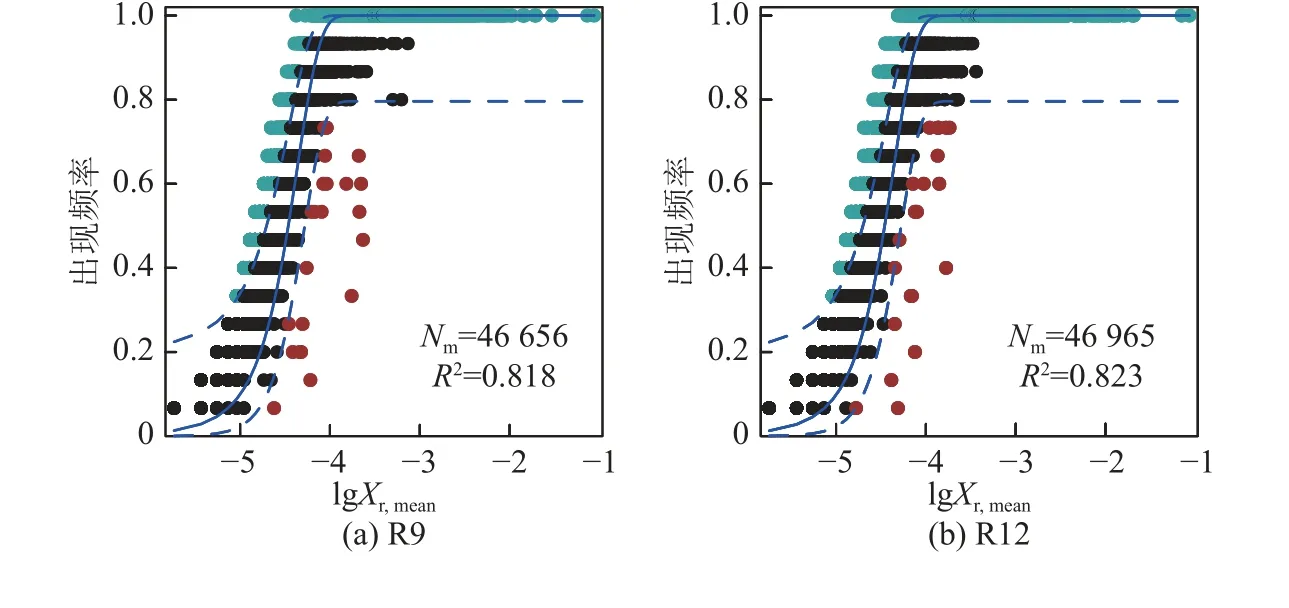
图4 不同复垦年限下土壤细菌群落结构的中性群落模型Fig.4 Neutral community model of soil bacterial community structure under different reclamation years
2.3 不同复垦年限下土壤碳循环功能基因丰度变化
图5 显示(图中Xa为基因的绝对丰度),不同复垦年限下淀粉、半纤维素、纤维素、几丁质、果胶和木质素等6 类物质降解过程及相关基因有显著差异(P<0.05)。其中,与淀粉降解相关的基因apu、amyA和amyX在R18 中丰度最高,复垦处理组丰度远高于CK(图5a)。与半纤维素降解相关的基因abfA、xylA和manB及与纤维素降解相关的CDH(编码纤维二糖脱氢酶基因)随复垦时长增加呈上升趋势(图5a)。与几丁质降解相关的chiA和与木质素降解相关的基因mnp、glx、pox和lig在R18 中丰度最高,远高于CK(图5a)。总体上R18 中碳降解相关功能基因丰度最高。碳固定相关的基因acsA、mct和rbcL丰度最高,acsB最低(图5b)。参与卡尔文循环的rbcL基因丰度随复垦时长呈增加趋势,除R9 外,均高于CK。参与还原三羧酸循环的korA基因和参与还原乙酰辅酶A 途径的acsA和acsB丰度在R18 中最高,约比CK 高一倍(图5b)。甲烷代谢功能基因丰度(pqq-mdh、pmoA、mxaF和mmoX基因)不同处理组几乎无显著差异(图5c)。

图5 不同复垦年限下土壤碳循环功能基因丰度的变化Fig.5 Variations of abundances of soil carbon cycling functional genes under different reclamation years
2.4 土壤碳循环微生物功能基因丰度与环境因素的相关性分析
利用随机森林模型评估了环境因子对碳循环功能基因丰度影响的重要性(图6),发现土壤固碳功能基因与土壤理化性状差异具有相关性。如图6c 所示,土壤pH 与甲烷代谢功能基因丰度相关,但其与碳降解、碳固定功能基因丰度并无显著相关性(图6a和图6b)。整体来看,BG、SOC、UE、TN、NN、CAT、AP、AN 与碳降解、碳固定和甲烷代谢功能基因具有相关性,且这些土壤理化性质和土壤酶活性可能是预测固碳功能的重要变量。
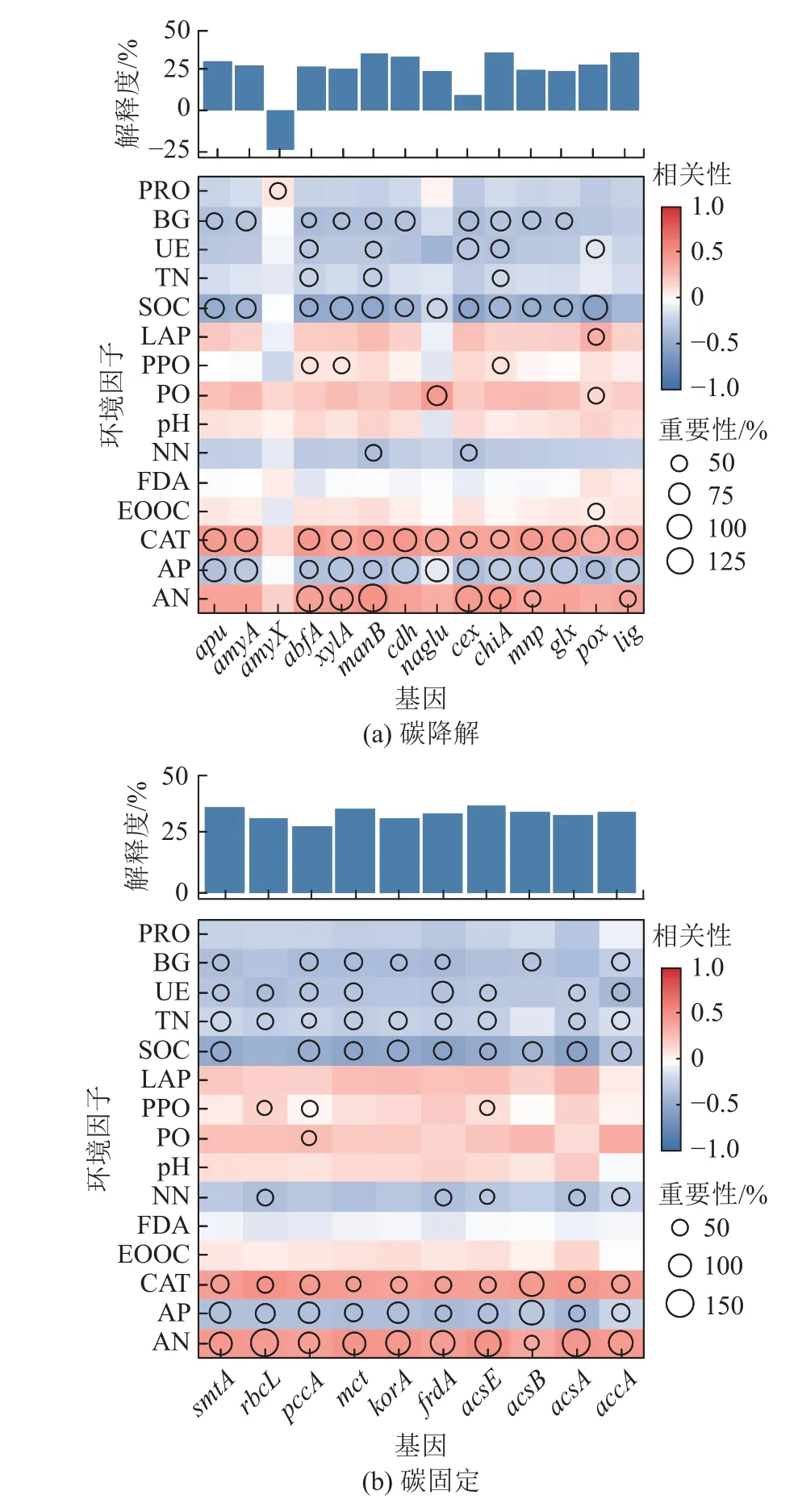
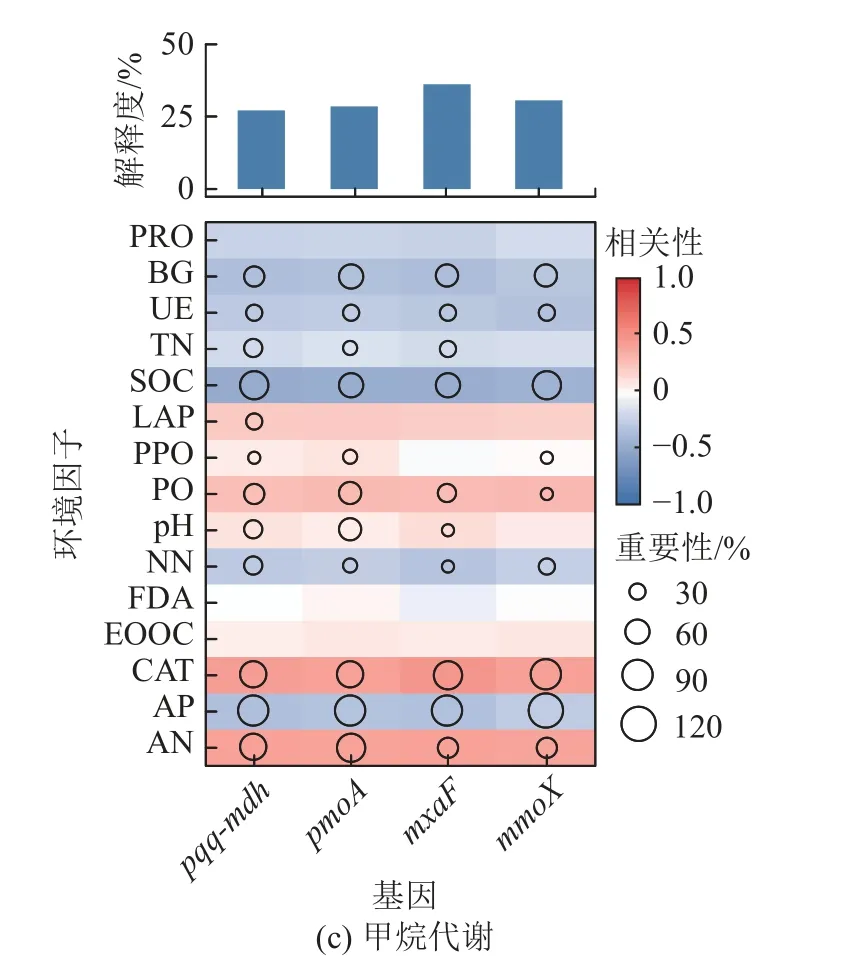
图6 环境因子对碳循环功能基因的潜在贡献Fig.6 Potential contribution of environmental factors to carbon cycle function genes
利用结构方程模型,建立土壤理化及酶活性、群落组装生态过程与碳循环功能基因丰度之间潜在的直接和间接联系,拟合分析理化性状对碳循环功能基因丰度的贡献。结果表明,复垦时长既可以直接影响SOC 含量和BG 酶活性,也可以通过不同生态过程对碳降解功能基因丰度产生间接影响(图7a)。此外,复垦时长能够影响土壤SOC 含量和BG、UE酶活性,进而间接影响碳固定过程功能基因丰度。同时,不同复垦年限通过影响SOC、BG 和UE 酶活性,再通过扩散限制、同质选择和异质选择间接影响碳固定相关功能基因丰度(图7b)。复垦及时长直接影响土壤AP、SOC 含量和BG 酶活性,再通过同质选择、扩散限制间接影响甲烷代谢相关功能基因丰度(图7c)。综上分析表明,复垦显著改善了矿物土壤SOC 和BG 酶活性,从而调控确定性过程和随机性过程占比,最终影响碳循环功能基因丰度。


图7 复垦过程中不同路径影响碳循环功能基因丰度的结构方程模型Fig.7 Structural equation models of rehabilitation affecting soil carbon cycle functional gene abundance through different ways
3 讨 论
3.1 复垦对土壤理化性质和酶活性的影响
东部平原矿区土壤偏弱碱性,丰富的降水导致Ca2+和Mg2+向下淋溶淀积强烈。沉陷复垦将下层土壤翻至上层,导致复垦土壤pH 增加。但耕作利用后pH 会下降[22-23]。这可能是本研究中土壤pH 变化趋势的主要原因。复垦土壤SOC(有机碳)不断下降,一方面是复垦改善了土壤通气,加速土壤碳释放;另一方面是大量氮肥引入,加速有机碳分解[24]。研究表明,土壤酶活性影响土壤养分有效性[25-26]。脲酶与氮循环相关,本研究中脲酶活性下降,可能与耕作大量施用氮肥有关,降低了对含氮有机物水解的需求[27]。复垦土壤PO(碱性磷酸酶)和PRO(蛋白酶)显著低于对照,表明重构土壤肥力远达不到未受采矿影响的耕地,这与张振佳等[28]研究结论一致。煤矸石充填带来大量金属离子,易与土壤中磷反应,形成不可溶性磷酸盐[27]。土壤PO 随复垦时长持续增加,说明复垦土壤需要大量PO 将不溶性磷转化为可溶性磷酸盐,提高磷的生物利用率和满足农作物生长[29]。土地复垦改善土壤理化和酶活性,养分供给、水分及物质循环[5],这与于亚军等[30]的研究结果一致。本研究中复垦土壤铵态氮、全氮、过氧化氢酶、亮氨酸氨基肽酶活性等持续增加,说明复垦后种植管理改善了土壤水肥状况,并随复垦时间持续积累,农田生态环境不断改善。
3.2 复垦年限对细菌群落组装过程的影响
了解微生物群落的组装过程能够识别微生物群落对复垦活动的响应[31]。确定性过程和随机性过程共同作用于微生物群落的构建,但二者相对重要性的大小由各种环境因素介导[32]。如资源供应充足可以增加随机性过程的重要性,大部分物种能够较好的生长时,随机性过程起主导作用。LAN 等[33]发现森林土壤细菌群落组装主要以随机性过程为主。随着外界环境变得苛刻,确定性过程起主导作用。本研究发现东部平原矿区复垦土壤细菌群落组装以随机性过程为主(图3 和图4),说明东部平原矿区土壤细菌群落结构与功能受随机选择操控,反映东部平原矿区自然环境限制并不十分苛刻。但不同处理组随机性组装过程中扩散限制不同,说明随复垦时长增加,微生物易利用资源不断增加,营养胁迫和环境选择压力逐渐削弱,改善了土壤环境,从而驱动群落组装过程中扩散限制增强[34]。此外,矿区土地复垦仅属局部扰动,不同于温度、降水等大尺度空间过程的影响,进一步证实了随机性过程通常发生于局部环境变化的小尺度空间[10]。
3.3 复垦年限对细菌群落固碳功能的影响
微生物功能基因编码参与物质循环的酶,能够反映土壤养分循环活动的强度。在本研究中,与碳降解相关的功能基因均在R18 中丰度最高,表明复垦及复垦时长提高碳降解酶对易分解和难分解碳的分解潜力,包含淀粉、半纤维素和纤维素的降解。复垦土壤中大量外源碳被土壤碳降解酶分解,释放出低分子的糖,为微生物生长代谢提供碳源和能量[35]。本研究发现除R9 外,复垦土壤rbcL基因丰度均高于CK,rbcL基因可作为分子标记用来反应微生物的固碳能力[36],可见长期复垦活动可以有效提高土壤的碳固持能力。研究表明,复垦活动通过改善土壤理化性质、微生物群落多样性等,进而影响土壤碳循环过程[37-39]。赵姣等[35]对黄土高原露采矿山排土场不同植被恢复下土壤碳循环功能基因研究,发现BG 和AN 含量是操纵碳循环相关功能基因丰度的主要因素。同样地,本研究发现BG、SOC、NN、CAT、AP 和AN 是预测固碳功能基因的重要变量,表明复垦介导的土壤养分状况密切影响土壤固碳微生物的活性和功能。结构方程模型结果进一步验证,复垦及时长可直接影响土壤SOC 含量和BG 酶活性,或通过SOC 含量和BG 酶活性间接影响扩散限制和同质选择过程,从而影响碳循环相关功能基因丰度。未来应持续监测复垦土壤SOC、BG 酶活性等参数,尝试田间管理措施调节这些参数达到影响微生物群落组装过程和提高固碳基因丰度的目的,促进复垦土壤的固碳潜力,提升矿区生态碳汇功能,同时为预测采矿干扰后土壤生态系统恢复提供了新的见解,对东部矿区生态复垦方案的提出和管理策略提供了理论依据。
4 结 论
1)东部平原矿区复垦对土壤理化性状和酶活性影响显著,复垦处理组土壤pH、铵态氮、过氧化氢酶及磷酸酶随复垦时间增长呈显著增加,有机碳、有效磷、硝态氮及脲酶、β-葡萄糖苷酶、蛋白酶则显著下降。
2)复垦改变土壤环境,从而影响细菌群落多样性、结构和分布模式。随机性过程主导了不同复垦年限土壤细菌群落的组装过程,且扩散限制的贡献最大。
3)有机碳、硝态氮、铵态氮、有效磷、β-葡萄糖苷酶及过氧化氢酶与复垦土壤固碳功能基因丰度之间相关性显著,东部平原矿区复垦活动有效增强土壤碳固持能力。
4)复垦或直接影响土壤有机碳含量和β-葡萄糖苷酶活性,或通过有机碳含量、β-葡萄糖苷酶活性间接影响扩散限制和同质性选择,进而影响碳降解、碳固定及甲烷代谢功能基因丰度。
猜你喜欢
阅读(科学探秘)(2023年3期)2023-04-10
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
中国科技纵横(2016年20期)2016-12-28
电测与仪表(2016年12期)2016-04-11
通信电源技术(2016年5期)2016-03-22
中国学术期刊文摘(2016年1期)2016-02-13
中国资源综合利用(2016年9期)2016-01-22
水生生物学报(2015年1期)2015-02-28
河南科技(2014年18期)2014-02-27
