
温州瓯江口春、秋季主要游泳动物生态位及其分化
2024-03-19 02:13周维维王晓波韩庆喜
海洋通报 2024年1期
周维维,王晓波,韩庆喜
(1.宁波大学 海洋学院,浙江 宁波 315832;2.禹治环境科技(浙江)有限公司,浙江 宁波 315199)
瓯江口位于浙江省南部近岸,是我国列长江口、黄河口、珠江口、钱塘江口之后的一大主要河口[1]。除此之外,其还是温台渔场的重要组成部分,海域内的游泳动物物种丰富。而游泳动物处于海洋生态系统的食物链顶端[2],是主要的海洋生物资源捕捞利用对象[3]。通过对瓯江口海域的游泳动物研究,不仅可为海洋生态系统的功能研究提供基础资料,也可为渔业资源管理策略的制订提供科学依据[4]。
生态位为当前生态学研究的核心内容之一,其理论在种间关系、群落结构、物种多样性及种群进化研究中被广泛应用[5]。生态位能体现自然生态系统中的一个种群在时间空间上的位置、相关种群之间的功能关系及利用资源的能力[6]。生态位被运用在不少陆生生物研究[7-8]中,但也被越来越多的学者运用到海洋生物群落的相关研究,如在舟山[9-10]、三门湾[11-12]等局部区域开展过少量研究。有关瓯江口附近海域海洋生物学的文献资料较少,仅见鱼类[13]、虾蟹类[14]群落结构和浮游动植物[15-16]的相关研究,未见有游泳动物生态位研究的相关报道。为丰富瓯江口海域游泳动物相关研究资料,本文以2020 年秋季(11 月)和2021 年春季(3月)在温州瓯江口附近海域进行的生物和环境调查数据为基础,比较分析春秋两季主要游泳动物的季节差异,利用生态位理论中的生态位宽度、生态位重叠两个方法分析瓯江口海域的游泳动物的生态位特征,根据RDA 结果分析主要游泳动物与环境因子间的关系。
1 材料与方法
1.1 站位分布
在瓯江口海域共布设12 个游泳动物采样站位(表1),时间为2020 年11 月(秋季)和2021 年3月(春季)。采样的12 个站位详见图1。在所有采样站位中,采样站位深度最深的是5 号站位,水深达15 m,深度最浅的为洞头群岛中间的15 号站位,其水深仅3.5 m;海水pH 值在8.11~8.27 之间,呈现弱碱性;温度在20~22.2 ℃之间。

图1 2020-2021年瓯江口海域游泳动物调查站位分布图

表1 2020-2021年瓯江口海域游泳动物调查站位经纬度
1.2 采样方法
在2020 年秋季和2021 年春季租用浙洞渔83006,使用单拖网(10.0 m(宽)×5.0 m(高)),网目范围2~7 cm,其中网囊网目为2 cm,拖网长度20 m,网板规格1.2 m×0.5 m,拖曳作业时网口水平扩展宽度约为6 m,每网拖曳0.5~1 h。海上底拖网调查过程中,规范记录各站位网产量、拖网时间等,调查按《海洋调查规范》(GB∕T12673-2007)和《海洋渔业资源调查规范》(SC∕T9403-2012)进行。调查渔获物总质量在40 kg 以下时,全部取样分析;大于40 kg时,从中挑出大型和稀有的标本后,从渔获物中随机取出渔获物分析样本20 kg左右。对渔获物均采用冰鲜冷藏,带回实验室对渔获物进行种类、渔获重量和尾数统计,记录网产量,并对每个种类进行主要生物学测定(体长、体重、成幼体等)。
除采集该海域的游泳动物外,还采集了对应海域的水样,水样的处理依照《海洋监测规范》(GB17378.4-2007)进行操作与分析。并对底质进行了分析检测,包括有机碳(Or-C)利用重铬酸钾氧化-还原容量法测得;硫化物(S)利用亚甲基蓝分光光度法测得;盐度计测量盐度(Sal);采用过滤重量法测量悬浮物(Turb);使用船载测深仪测量水深(Depth)、溶解氧(DO)通过碘量法测得;化学需氧量(COD)通过海碱性高锰酸钾法测得;活性磷酸盐(Ph-S)通过磷钼蓝分光光度法进行测得;亚硝酸盐-氮浓度通过奈乙二胺分光光度法测得;硝酸盐-氮浓度通过锌-镉还原法测得;氨-氮浓度通过次溴酸盐氧化法测得;叶绿素a(Chla)浓度通过荧光分光光度法测得。此外,亚硝酸盐、硝酸盐-氮与氨-氮加起来的浓度为无机氮(In-N)浓度。
根据海洋动植物的生长、生殖适温范围划分暖水种和温水种,适宜生长在20°C 以上的为暖水种,适宜生长在4~20°C的为温水种。
1.3 数据分析
利用Jaccard 种类相似性指数[17]计算春、秋季游泳动物的相似度,计算公式如下:
式中:a为春季游泳动物种类数;b为秋季游泳动物种类数;c为春秋2 季调查共有的游泳动物种类数。当0≤SJ<0.25 时,为极不相似;当0.25≤SJ<0.5时,为中等不相似;当0.75≤SJ≤1.00时,为极相似。
游泳动物相对重要性指数的计算公式[18]:
式中:N%为某一游泳动物尾数占总尾数的百分比;W%为该游泳动物重量占总游泳动物重量的百分比;F%为该游泳动物的站位出现率;IRI为相对重要性指数。当IRI>500时,该物种为优势种。
Levins生态位宽度计算公式[19]:

式中:R为站位总数;Bi为生态位宽度,数值变化范围为[0,R];Pij为物种i在站位j总物种数中所占的比例;Bi值越大,生态位宽度越大;Pkj为物种k在站位j总物种数中所占的比例;Oik为生态位重叠值,数值变化范围为[0,1],Oik越大,生态位重叠值越大。
1.4 数据处理
利用ArcGIS Desktop 10.6 软件根据采样经纬度制作采样站位分布图;优势度指数、优势种更替率和Jaccard 种类相似性指数均利用Microsoft Excel 2019软件进行计算;生态位宽度值和生态位重叠值利用R4.2.1 软件计算;利用PRIMER 软件对生态位宽度进行相似性等级聚类分析(CLUSTER);冗余分析(RDA)通过Canoco 5.0软件得到。
通过DCA分析,春秋两季的Lengths of gradient的第一轴的分析结果均小于3,所以选择冗余分析(RDA)。
2 结果
2.1 游泳动物优势种
通过相似性指数计算得出,春、秋两季游泳动物的Jaccard 种类相似性指数值为1.00,为极相似。当IRI>500 时,可判断该物种为优势种。秋季,游泳动物优势种共6 种,分别为棘头梅童鱼(Collichthys lucidus)、刀鲚(Coilia nasus)、龙头鱼(Harpadon nehereus)、口虾蛄(Oratosquilla oratoria)、三疣梭子蟹(Portunus trituberculatus)、日本蟳(Charybdis japonica);春季游泳动物优势种共10种,分别为棘头梅童鱼、刀鲚、口虾蛄、三疣梭子蟹、日本蟳、孔虾虎鱼(Trypauchen vagina)、花鲈(Lateolabrax japonicus)、拉氏狼牙虾虎鱼(Odontamblyopus lacepedii)、脊尾白虾(Palaemon carinicauda)、细巧仿对虾(Batepenaeopsis tenella)。从适温性来看,秋季有2 种温水种,4 种暖水种;春季有6种暖水种,4种温水种,如下表2所示。

表2 春秋季主要游泳动物相对重要性指数
2.2 游泳动物优势种生态位宽度
从图2 可知,春秋两季游泳动物优势种的生态位宽度变化范围为1.20~7.53,其中春季的日本蟳(7.53)的生态位宽度最大,最小的是春季的口虾蛄(1.20),根据变化将所有的物种总共分为三类,第一类为4 个广生态位种(Bi≥6),分别为日本蟳(春秋季)、棘头梅童鱼(秋季)、脊尾白虾、三疣梭子蟹(秋季);第二类为7 个中生态位种(3<Bi<6),分别为棘头梅童鱼(春季)、刀鲚(春季)、龙头鱼、口虾蛄(秋季)、三疣梭子蟹(春季)、孔虾虎鱼、花鲈;第三类为3 个窄生态位种(0<Bi≤3),分别为刀鲚、细巧仿对虾、拉氏狼牙虾虎鱼、口虾蛄(春季)。

图2 春秋季主要游泳动物生态位宽度值
秋季,游泳动物优势种的生态位宽度变化范围为2.08~6.86,其中棘头梅童鱼(6.86)的生态位宽度最大,其次为日本蟳(6.72),生态位宽度最小的是刀鲚(2.08)。根据变化情况,秋季有棘头梅童鱼、三疣梭子蟹、日本蟳3 种广生态位种,有龙头鱼、口虾蛄2种中生态位种,有刀鲚1种窄生态位种。并根据聚类分析可将游泳动物分为3组(图3),第1 组为口虾蛄、龙头鱼;第2 组为棘头梅童鱼、三疣梭子蟹、日本蟳;第3 组仅有刀鲚。
春季,游泳动物优势种的生态位宽度变化范围为1.20~7.53,其中日本蟳(7.53)的生态位宽度最大,其次为脊尾白虾(7.35),生态位最小的是口虾蛄(1.20)。根据生态位宽度变化情况,春季有日本蟳、脊尾白虾2 种广生态位种,中生态位种有棘头梅童鱼、刀鲚、三疣梭子蟹、孔虾虎鱼、花鲈5 种;有口虾蛄、拉氏狼牙虾虎鱼、细巧仿对虾3 种窄生态位种。并根据聚类分析可将游泳动物分为4 组(图4),第1 组由棘头梅童鱼、刀鲚、三疣梭子蟹、孔虾虎鱼、花鲈5 种组成;第2 组由细巧仿对虾、拉氏狼牙虾虎鱼2 种组成;第3 组由脊尾白虾、日本蟳2 种组成;第4 组仅由口虾蛄组成。
2.3 游泳动物优势种生态位重叠
根据从表3和表4可知,春秋两季游泳动物优势种生态位重叠变化范围为0.03~0.96。其中秋季生态位重叠变化范围为0.05~0.88。其中,Oik>0.6的种对有8对,占总数的53.3%,生态位重叠值最大的日本蟳和三疣梭子蟹种对;0.3≤Oik≤0.6 的种对2 对,占13.3%;Oik<0.3 的种对5 对,占33.4%,刀鲚和口虾蛄的重叠值最小(0.05)。春季生态位重叠值变化范围为0.03~0.96。其中,Oik>0.6 的种对13 对,占总数的28.9%,重叠值最大(0.96)的种对是拉氏狼牙虾虎鱼和口虾蛄;0.3≤Oik≤0.6的种对20 对,占44.4%;Oik<0.3 的种对12 对,占26.7%,细巧仿对虾和日本蟳的重叠值最小(0.03)。

表3 秋季主要游泳动物生态位重叠值

表4 春季主要游泳动物生态位重叠值
2.4 游泳动物生态位分化
对温州瓯江口游泳动物优势种和有机碳(Or-C)、硫化物(S)、活性磷酸盐(Ph-S)、悬浮物(Turb)、水深(Depth)、盐度(Sal)、叶绿素a(Chl-a)、溶解氧(DO)、无机氮(In-N)、化学需氧量(COD)这几种环境因子进行RDA分析。秋季,第1、2 轴的解释率分别为55.78% 和23.29%,与环境因子的相关系数为0.990 1 和0.865 4且能解释88.18%的现象(表5),说明物种与环境因子相关性较强。春季,第1、2 轴的解释率分别为45.64%和23.32%,与环境因子的相关系数分别为0.977 1 和0.802 3,且能解释83.45%的现象(表5),说明物种与环境因子相关性较强。综合两季的结果可知(图5),对主要游泳动物影响最大的环境因子是叶绿素a和盐度。其中口虾蛄分布主要受叶绿素a的影响,其分布与叶绿素a呈正相关;三疣梭子蟹和日本蟳分布主要受盐度的影响,其分布与盐度呈正相关。
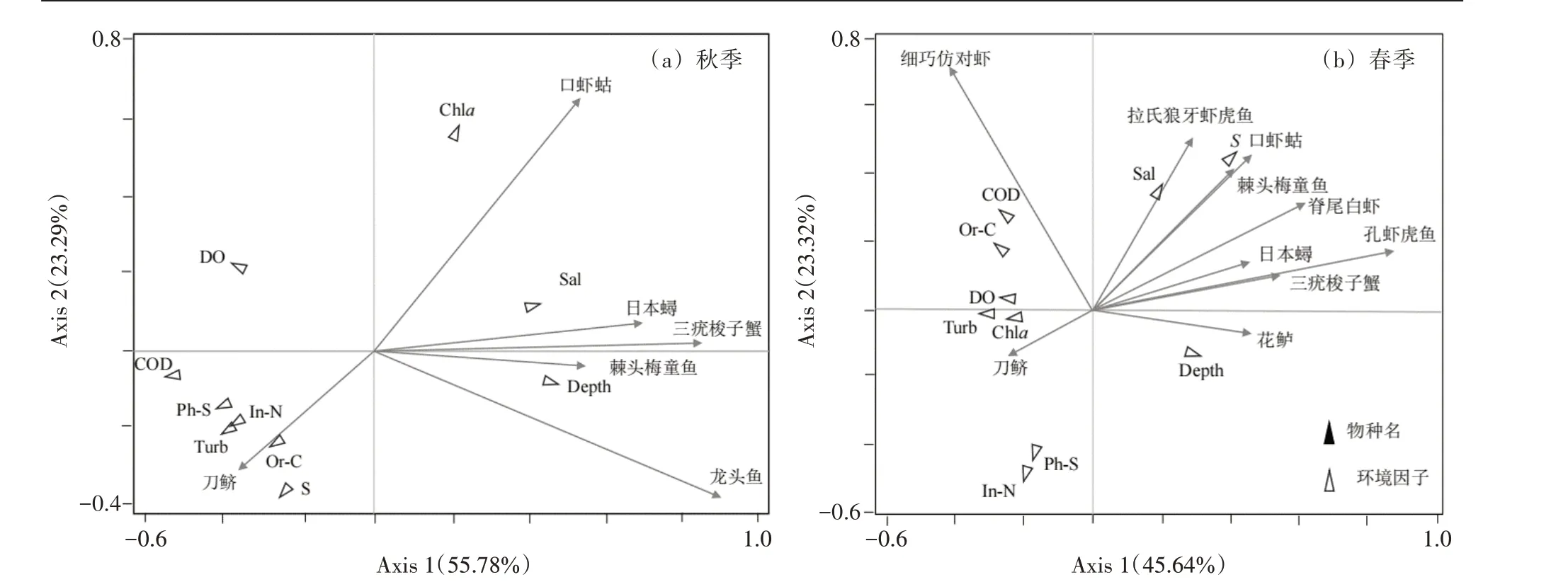
图5 春、秋季主要游泳动物RDA排序图

表5 主要游泳动物与环境因子RDA统计结果
3 讨论
3.1 游泳动物优势种组成分析及其变化
根据海洋动植物的生长、生殖适温范围将它们划分为三个类型:暖水种,包括适宜生长在25°C及以上的热带种和20~25°C 的亚热带种;温水种,包括12~20°C 的暖温种和4~12°C 的冷温种;冷水种,包括0~4°C 的亚寒带种和0°C 的寒带种[21]。本研究发现,该区域春秋季的游泳动物以暖水种和温水种为主(表2),由于瓯江口附近海域地处亚热带季风气候区,受气候影响,该区域降水丰富、气候温暖、湿度较大[22],比较适宜该类物种的生存。春秋两季瓯江口海域物种相对重要性指数较高的有三疣梭子蟹、日本蟳、孔虾虎鱼、龙头鱼、拉氏狼牙虾虎鱼、棘头梅童鱼、口虾蛄、刀鲚、脊尾白虾、细巧仿对虾、花鲈等11 种,与2012年[23]和2016 年[24]乐清湾附近海域调查结果相比,增加了孔虾虎鱼、拉氏狼牙虾虎鱼、口虾蛄、脊尾白虾、细巧仿对虾等5 种。说明瓯江口海域渔业资源较为丰富,而春季优势种较秋季多,但是减少了小黄鱼这一物种,这是因为该物种长期受到过度捕捞、环境污染、挖沙采矿、洄游通道受阻等众多环境压力,再加上小黄鱼的产卵亲体被渔民捕捞,使小黄鱼亲体数量大量减少,久而久之导致物种数量减少。
秋季多数物种相对重要性指数大于春季,但春季优势种较秋季更为丰富。春秋两季共有优势种为棘头梅童鱼、刀鲚、口虾蛄、三疣梭子蟹、日本蟳等5 种,秋季多数物种相对重要性指数大于春季,尤其是口虾蛄,其相对重要性指数远高于其他物种,为两个季节的绝对优势种。这与2019 年[25]、2007 年[26]瓯江口附近海域调查结果一致,说明了瓯江口附近是口虾蛄的索饵和育苗的重要场所之一。虾类物种数量和分布与暖流有关[26],春季,瓯江径流较大,会冲击来自台湾高温高盐暖流[24],形成的冲淡水水团是导致春季口虾蛄物种相对重要性指数低的原因。秋季,瓯江径流减少,汇入海域的淡水减少,海域受台湾高温高盐暖流影响更大,此时海域环境非常适合虾类生长,使得口虾蛄的相对重要性指数明显提升。同样,春季优势物种较秋季更多,也是因为受春季瓯江冲淡水水团的影响,海域混合水域广阔,海洋环境更复杂[27],水中的植物和浮游动物更丰富[28],饵料资源的丰富与分布广泛导致各类物种的分布范围也更广,更容易在采样过程中被获取。
从分析可以看出,较往年相比,虽然瓯江口海域的游泳动物优势种数量丰富了,但是一些物种的减少也不容忽视。深入研究物种的特性,分析其减少的原因迫在眉睫。游泳动物作为海洋生态环境的重要物种,物种的减少会影响群落结构,进一步影响生物多样性。此外,游泳动物生存发展不单单受自身生物学特性限制,还受同一生境中其他生物的影响。所以对部分物种需要采取一些人为的保护措施,如改革传统的渔业方式,严格执行“伏季休渔”的政策,加大对违法捕捞行为的惩罚力度。保护海洋水资源,为游泳动物的生存营造良好的生境。加强对物种功能基因、生物学特性的相关研究,选择适应性强的物种亲体进行培育,提供优质的遗传样本。
3.2 游泳动物优势种生态位宽度特征
从结果可以看出,瓯江口海域游泳动物间的生态位宽度差异明显,且中生态位种的数量多。生态位宽度可以用于评价物种对资源的利用情况,一般来说,生态位宽度越大,其对海域内资源的利用能力越强,适应能力也越强,分布范围越广泛[29]。
通过聚类分析将秋季主要游泳动物分为3 组(图3),第一组为口虾蛄、龙头鱼,因两者生存环境相似,均为广生态位种和暖水种。第二组由日本蟳、棘头梅童鱼和三疣梭子蟹组成,三者均为广生态位种且生态位宽度大,其中棘头梅童鱼的生态位宽度最大,因其生态习性和适宜生境与其他游泳动物相似,而且温州瓯江口海域为棘头梅童鱼的产卵场,亲体分布范围广,对环境内的资源利用大[30]。日本蟳其次,它的全年最高渔获量(8.01 kg∕km2)出现在秋季[31],此外,其在瓯江口秋季的11 个站位和春季的10 个站位中均有分布。第三组,仅有刀鲚,其生态位宽度为秋季最低的物种,刀鲚为常见的溯河洄游型鱼类,仅在繁殖季节(春季)回到瓯江口附近海域进行产卵[32],所以秋季该物种分布少且范围不广,导致其物种生态位宽度小。
春季生态位宽度聚类分析将主要游泳动物分为4组(图4),其中第一组为花鲈、棘头梅童鱼、三疣梭子蟹、刀鲚、孔虾虎鱼,均为中生态位种,主要是因为花鲈、棘头梅童鱼、刀鲚、孔虾虎鱼都会在春季回到瓯江口附近进行产卵,在海域内的分布范围广,生态位宽度大。第二组为拉氏狼牙虾虎鱼和细巧仿对虾,两者均为窄生态位种且生态位宽度小,可能是因为这两个物种在调查海域内资源分布不均,其中细巧仿对虾在春季采样中的站位出现率仅为58%,导致其生态位宽度不大。而拉氏狼牙虾虎鱼在12 个站点的样品中,站位之间的物种数量差距大,如其在22 号站位的采样数量为1 号站位的20 倍,反映在海域中其表现为斑块状分布,分布不均匀导致其生态位宽度窄。该结果与已有文献[33]中的各站点间较大的物种数量或分布差异导致该物种生态位宽度低的结论一致。值得注意的是第四组仅为口虾蛄,其在秋季为中生态位种,但是到了春季却变成了窄生态位种,这是因为虾类分布受水温影响较大,在温度较高的位置往往分布较多[26]。而秋季瓯江口附近海域较春季受台湾暖流影响较大,所以秋季口虾蛄分布范围广,生态位宽度大,春季则反之。
此外,通过比较分析主要游泳动物的生态位宽度和相对重要性指数,发现两者并没有直接关系。如秋季口虾蛄的相对重要性指数(2 763.12)高于三疣梭子蟹(2 147.06),而其生态位宽度(4.54)低于三疣梭子蟹(6.00);春季孔虾虎鱼的相对重要性指数(1 862.58)高于花鲈(1 698.41),而生态位宽度(4.02)低于花鲈(5.00),说明优势种会因为本身的生态特征或者其他原因导致生态位宽度值不高[34],而且在春秋两季的优势种中,都存在广、中、窄三个类型的生态位种,这也验证了优势种并不一定分布在广生态位[35-36]。
综上所述,物种的生态适应能力、竞争能力、资源利用能力,以及分布范围和数量是影响生态位宽度的重要因素,生态位宽度是众多因素综合作用的结果,不能单一地凭靠优势种判断物种的生态位宽度大与否。截至目前的瓯江口附近研究中,日本蟳和三疣梭子蟹的生态位宽度常年保持较高水平,说明日本蟳和三疣梭子蟹是瓯江口海域的主要品种。除此之外,瓯江口海域物种的生态位宽度普遍不高,是海域内天然饵料资源不足、适宜生境不多、物种间竞争压力大的表现,随着海洋生态环境日趋恶劣,很多生态位宽度低的物种受自身对环境变化有着高敏感反应,适应能力差,资源利用能力弱的缺点,导致其在生境中的分布范围越来越小,存在被淘汰的风险。
3.3 游泳动物优势种生态位重叠
瓯江口海域游泳动物生态位重叠明显,且具有季节性差异。生态位重叠值不仅受物种间的竞争关系[37]影响,而且受物种间的捕食关系[38]影响。通常来说,生态位重叠值越高,两个物种竞争关系越激烈或两者存在捕食关系。秋季,龙头鱼与其他物种间的生态位重叠值较高(表3),主要原因是龙头鱼属肉食性,其饵料种类繁多,最主要的是以游泳动物为食,通过对其饵料成分分析发现,饵料成分中游泳动物的出现频率高达67.27%[39],说明龙头鱼会以区域内的游泳动物为食,龙头鱼与其他游泳动物之间的捕食关系导致它们之间的生态位重叠值较高。而刀鲚与其他物种的生态位重叠较小,因为刀鲚以摄食浮游动物为生[40],与其他游泳动物不构成竞争或者捕食关系。春季,生态位重叠值最高的种对为拉氏狼牙虾虎鱼和口虾蛄(0.96),拉氏狼牙虾虎鱼为暖水性的河口定居物种,一般穴居在水底的泥层中[41],摄食多毛类和小型底栖甲壳动物[42],在2-4 月和7 月下旬至9 月繁殖,为海域常见的春季优势种。口虾蛄为杂食性的暖水种,且瓯江口海域为口虾蛄的产卵场和索饵场,尽管两者属于不同的物种,但是两者的生存环境相似,存在一定的竞争关系,导致两者的生态位重叠较高。生态位重叠次高的为棘头梅童鱼和拉氏狼牙虾虎鱼种对(0.92),这是因为两者在分布范围和洄游以及繁殖季节方面重叠程度较高,竞争激烈,从而导致生态位重叠明显。
此外,游泳动物的生态位宽度大,并不代表物种间的生态位重叠值高。如秋季广生态位种棘头梅童鱼(6.86)和窄生态位种刀鲚(2.08)的生态位重叠值为0.07。春季窄生态位种口虾蛄(1.20)和中生态位种棘头梅童鱼(4.83),两者的生态位重叠值高达0.87,原因是两者均为广温广盐性种,都生活在泥沙底质的浅海,分布范围上重叠度较大,当空间资源不足以供给两者正常生存时,会通过竞争的方式获得生存空间,竞争关系会导致两者的生态位重叠值较高。
游泳动物种对间的生态位重叠与优势种、生态位宽度有一定的联系,但并不绝对。在瓯江口海域通常优势度高、生态位宽度大的物种生存能力强,环境适应能力强,能够占据大范围的优质生境,与环境中的其他游泳动物产生激烈的竞争从而导致生态位重叠明显。将三个指标综合起来能够更直观地分析海域内渔业资源衰退的现状,方便相关部门“对症下药”,为维护瓯江口海域的海洋生态环境作出切实努力。
3.4 主要游泳动物的生态位分化
游泳动物的生存与海洋环境密切相关,尤其是海水质量、微生物等环境因子的变化会引起部分敏感度高的游泳动物发生群落结构变化,因此调查分析环境因子与物种分布至关重要。冗余分析能反映物种分布与环境因子之间的关系。本研究发现,瓯江口附近海域主要游泳动物分布会受到叶绿素a和盐度等环境因子的影响。盐度变化能够对游泳动物的生存造成巨大影响[43],盐度梯度为许多生物的生理屏障,对物种的免疫系统至关重要[44]。春季盐度范围为12.2~24.1 且第一轴与盐度的相关系数为0.200 9,说明三疣梭子蟹和日本蟳的分布在春季与盐度呈正相关;还有叶绿素a的浓度变化也会对游泳动物的分布产生影响,叶绿素a的浓度与海域内浮游植物有关,叶绿素a的浓度通过影响浮游植物从而影响部分游泳动物的生存,当水体中的叶绿素a 浓度高时,会促进浮游植物的生长,浮游植物的生长超过水体可承受范围时,会造成水体富营养化,水环境中营养盐、有机质含量升高会消耗水体中的溶解氧,加速水体恶化,不利于游泳动物的生存。瓯江口海域秋季叶绿素a浓度明显高于春季,而口虾蛄对该变化较为敏感,其分布与叶绿素a浓度变化密切相关。
此外,冗余分析不仅可以解释物种分布与环境因子之间的关系,还可解释生态位重叠测定方面得到的一些不能合理解释的数据[44]。如无论刀鲚生态位宽度是否变化,其始终与其他物种间的生态位重叠不显著,主要是因为它与其他游泳动物不具有相似的底栖生境,且分布区域不同,不构成竞争或捕食关系,导致其虽然生态位宽度有变化,但是与其他主要游泳动物生态位重叠不显著。通过冗余分析可知,温州瓯江口海域游泳动物受环境因子影响较小,说明海域环境保护卓有成效,使得渔业资源紧张得到一定缓解。
4 结论
2020 年秋季(11 月)和2021 年春季(3 月)瓯江口附近海域共采获游泳动物5 大类78 种。秋季优势种6 种,春季优势种10 种,春秋两季共有的优势种5种,更替率为31.25%。
2020 年瓯江口海域秋季主要游泳动物生态位宽度范围为2.08~6.86,生态位重叠共15 对种对,其数值范围为0.05~0.88,广生态位种占总物种数的50%。春季主要游泳动物生态位宽度范围为1.20~7.53,生态位重叠共45 对种对,其数值范围0.03~0.96,其中中生态位种占总物种数的50%。生态位宽度与生态位重叠之间并不是绝对正相关。春秋两季主要游泳动物的分布与叶绿素a、盐度等环境因子有关,且能利用冗余分析结果分析种对间生态位重叠中不能合理解释的数据。
温州瓯江口海域游泳动物物种丰富且群落结构稳定,主要为刀鲚、孔虾虎鱼等洄游型鱼类的产卵和育幼场所,随着海域环境保护力度加强,效果显著,使得海域内许多物种资源得到一定恢复。
猜你喜欢
发明与创新·中学生(2024年2期)2024-02-03
发明与创新(2024年6期)2024-01-20
——皮皮虾
科学大众(2023年20期)2023-10-26
科学大众(中学)(2023年10期)2023-10-20
Zoological Research(2022年3期)2022-06-07
今日农业(2021年8期)2021-11-28
水上消防(2021年3期)2021-08-21
上海公路(2019年3期)2019-11-25
人大建设(2019年5期)2019-10-08
中华诗词(2018年7期)2018-11-15
