
PINK1/Parkin 介导的线粒体自噬在力学失衡诱导终板软骨退变中的作用
2024-04-01 12:03吴明凡孙良业许俊胜安徽医科大学附属六安医院骨科安徽六安237001
局解手术学杂志 2024年3期
郑 权,吴明凡,邵 松,孙良业,许俊胜 (安徽医科大学附属六安医院骨科,安徽 六安 237001)
椎间盘退变是引起颈腰痛及神经损害的重要因素,而椎间盘退变主要由异常应力、细胞衰老、炎性介质分泌以及营养供应障碍等引起。目前认为终板途径是椎间盘营养供应的主要途径,终板软骨为透明软骨,终板软骨细胞分泌的Ⅱ型胶原(type Ⅱ collagen,COL-2A)、蛋白聚糖(aggrecan,ACAN)等细胞外基质对于椎间盘的营养供应以及形态维持都具有重要的作用[1]。有研究表明,终板软骨的损伤可使相邻椎间盘的压力分布发生改变,髓核内营养供应减少,从而诱导椎间盘退变[2]。研究显示,异常力学刺激是导致终板软骨退变的主要因素,脊柱稳定性降低会导致椎间盘内应力异常,直接导致终板软骨及椎间盘退变[3]。然而异常应力诱导终板软骨退变的机制仍需进一步深入研究。
自噬是细胞降解和再利用功能异常蛋白质和其他大分子物质的过程,通过自噬降解及重复利用细胞内部结构,有助于细胞适应外界的刺激及存活。线粒体自噬作为一种选择性自噬,能够清除细胞内受损的线粒体,维持细胞内环境及功能稳定。目前认为线粒体自噬主要是由PINK1/Parkin 信号通路以及不依赖PINK1 的受体介导[4]。PINK1/Parkin 信号通路作为介导线粒体自噬重要且较为成熟的信号通路,具有维持软骨细胞功能的作用[5]。有研究表明,在关节软骨退变过程中,由PINK1/Parkin 信号通路介导的线粒体自噬水平明显降低,过表达Parkin 基因能够明显提高软骨细胞COL-2A、ACAN 等细胞外基质的水平,从而减轻关节软骨退变[6]。终板软骨与关节软骨同属透明软骨,但线粒体自噬对于应力刺激诱导终板软骨退变的影响及作用机制目前尚未见报道。本研究通过构建大鼠脊柱失稳模型,探讨力学失衡条件下PINK1/Parkin信号通路介导线粒体自噬对于终板软骨退变的影响,以期为椎间盘退变的相关机制研究提供参考。
1 材料与方法
1.1 实验动物
清洁级SD 大鼠18只(10周龄),雌雄不限,体质量(180±20)g,上海西普尔一必凯实验动物有限公司提供,许可证号:SCXK(沪)2022-0035。本研究获安徽医科大学附属六安医院实验动物伦理委员会批准(20220407001)。
1.2 试剂和仪器
DMEM/F12(美国Hyclone 公司);胎牛血清(美国Gibico 公司);胰蛋白酶、COL-2A 酶(美国Sigma-Aldrich 公司);TRIzol(美国Invitrogen 公司);PCR 反应体系和反转录试剂盒(美国Fermentas 公司);RT-PCR仪(德国Roche 公司,LightCycler 480);NanoDrop 2000(美国Thermo&Scientific 公司);Odyssey 红外成像系统(英国Li-Cor 公司);羰基氰化物3-氯苯腙(carbonyl cyanide 3-chlorophenylhydrazone,CCCP);兔抗鼠COL-2A抗体(美国Cell Signaling Technology 公司,1:1 000);兔抗鼠ACAN、Parkin 抗体(英国Abcam 公司,1:1 000),兔抗鼠PINK1、Tomm20、Timm23 抗体(美国Cell Signaling Technology 公司,1:1 000),山羊抗兔二抗(美国Bioworld Technology公司,1:5 000)。
1.3 大鼠脊柱失稳模型的构建及分组
取18只SD大鼠,分为正常组、退变组及CCCP组,每组6 只。正常组大鼠未做任何处理。退变组:大鼠以6.5%的水合氯醛0.055 mL/kg 腹腔注射麻醉后取俯卧位,固定四肢,腰背部剃毛备皮约4 cm×8 cm,消毒铺单,沿腰背部棘突作后正中切口,切开皮肤后,沿骨膜下剥离。用咬骨钳去除L2~L5的棘上韧带、棘间韧带以及两侧关节突,切断棘突两侧的棘旁肌,逐层缝合皮下筋膜、皮肤。术后连续3 d,每只SD大鼠给予青霉素5 万U 肌肉注射。CCCP 组:在退变组手术基础上,参考Kim 等[7]的方法沿纤维环紧贴终板软骨向椎间盘注射CCCP(10 μmol/L)5 μL。所有大鼠单笼饲养,连续饲养16周。
1.4 石蜡切片的制作及组织染色
包埋与切片:实验结束后处死各组大鼠,获取L2~L5椎间盘组织,4%多聚甲醛固定24 h,EDTA 液中脱钙1 个月,流水冲洗干净,常规脱水、透明、浸蜡、包埋。将蜡块置于切片机上切片,取5 μm 厚的切片,置于载玻片上,37 ℃烘片过夜,标记后进行组织染色。HE染色:切片脱蜡、脱水,苏木精染色3 min,自来水冲洗;1%盐酸酒精分化,流水冲洗10 min,伊红染色2~3 min,自来水冲洗,梯度脱水后中性树胶封片后观察。番红-固绿染色:石蜡切片常规脱蜡,自来水冲洗30 s,固绿染色5 min;自来水冲洗1 min,番红染色5 min。1%冰醋酸分色1 s,95%乙醇脱水2 次,每次1 min,100%乙醇脱水2 次,每次1 min,二甲苯透明2次,每次5 min,中性树胶封固后观察。
1.5 RNA提取及 RT-PCR
取各组大鼠的椎间盘组织,在4 倍解剖显微镜下用尖刀切开,刮除髓核及纤维环,获取终板软骨放入研钵中,研磨至粉末状。常规TRIzol 法提取终板软骨组织的RNA,NanoDrop 2000 检测RNA 的浓度与纯度;取1 μg 总RNA,采用一步法20 μL 体系[DEPC 水12.5 μL,Oligo dT(100 mol/L)1 μL,5×反转录缓冲液4 μL,200 U反转录酶0.5 μL,dNTP(浓度为10 mmol/L)2 μL]反转录合成cDNA,将cDNA 稀释5 倍,按20 μL体系在设定的程序下进行PCR 扩增,相关引物见表1。反应条件:95 ℃预变性30 s;95 ℃变性5 s,60 ℃退火延伸25 s,扩增50 个循环。以GAPDH 为内参,灭菌超纯水作为阴性对照,每个模板设置3个复孔,结果使用相对定量2-△△Ct法分析。

表1 RT-PCR 引物序列
1.6 蛋白提取及Western blot
分离获取终板软骨组织,尽量剪碎研磨至粉末状,采用RIPA 蛋白提取缓冲液和蛋白酶抑制剂PMSF提取总蛋白,BCA 试剂盒测定蛋白浓度,取20 μg 蛋白样品,十二烷基磺酸钠聚丙烯酰胺凝胶电泳,湿转法转膜,5%脱脂牛奶室温封闭1 h,加入一抗后4 ℃孵育过夜,洗涤后加入二抗室温孵育1 h,Odyssey 机器成像。采用Image J软件测定蛋白条带灰度值,目标蛋白灰度值与GAPDH 灰度值的比值为蛋白相对表达水平。
1.7 统计学处理
实验数据采用SPSS 22.0 软件进行分析,结果以均数±标准差()表示,多组间数据比较采用单因素方差分析,组间两两比较采用LSD-t检验。α=0.05 为检验水准,P<0.05为差异有统计学意义。
2 结果
2.1 力学失衡对大鼠椎间盘组织形态的影响
HE 染色显示,退变组大鼠脊柱失稳诱导的椎间盘组织破坏明显,终板软骨及纤维环结构也明显受损;而CCCP 组大鼠椎间盘与退变组比较,髓核较为完整,终板软骨及纤维环结构受损较轻,见图1。

图1 各组大鼠椎间盘组织HE染色
番红-固绿染色结果显示,退变组大鼠脊柱失稳诱导的髓核及终板软骨细胞外基质分泌均较正常组明显减少,CCCP 组大鼠中终板软骨细胞外基质分泌较退变组明显增多,见图2。

图2 各组大鼠椎间盘组织番红-固绿染色
2.2 力学失衡诱导大鼠终板软骨退变
RT-PCR 结果显示,与正常组比较,退变组大鼠脊柱失稳诱导的终板软骨组织中ACAN、COL-2A mRNA的表达明显下调(P<0.05);而CCCP 组大鼠终板软骨组织中ACAN、COL-2A mRNA 的表达明显增高(P<0.05),见图3a。Western blot 结果显示,退变组大鼠终板软骨中ACAN、COL-2A 蛋白水平较正常组明显降低(P<0.05),而CCCP 组大鼠终板软骨中ACAN、COL-2A 蛋白水平较退变组明显增高(P<0.05),见图3b、c。

图3 各组终板软骨组织中ACAN和COL-2A mRNA及蛋白表达
2.3 PINK1/Parkin 信号通路介导的线粒体自噬在终板软骨组织中的表达
RT-PCR 结果显示,与正常组比较,退变组大鼠终板软骨PINK1、Parkin mRNA 表达明显降低(P<0.05);CCCP 组大鼠终板软骨组织PINK1、Parkin mRNA 表达较退变组明显增高(P<0.05),见图4a。此外,Western blot 结果显示,退变组大鼠终板软骨PINK1 及Parkin蛋白水平较正常组均明显降低(P<0.05),而线粒体蛋白Tomm20 及Timm23 水平较正常组增高(P<0.05);CCCP 组大鼠终板软骨中PINK1 及Parkin 蛋白水平较退变组明显增高(P<0.05),线粒体蛋白Tomm20 及Timm23水平较退变组明显降低(P<0.05),见图4b、c。
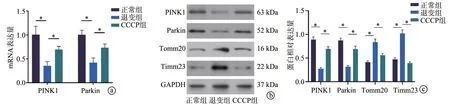
图4 各组终板软骨组织中线粒体自噬相关mRNA及蛋白表达量
3 讨论
椎间盘作为人体最大的无血管组织,其营养供应主要依赖椎体两端终板软骨以及外层纤维环的渗透作用。终板软骨是连接椎体与椎间盘的薄层透明软骨,其特殊结构能够维持椎间盘的形态及功能的稳定。一方面,终板软骨分泌的COL-2A、ACAN 等细胞外基质有利于营养物质向椎间盘的运输[8];另一方面,终板软骨对于椎体与椎间盘之间的力学传递具有重要的缓冲作用[8]。研究显示,终板退变是椎间盘退变的始动因素,维持终板软骨的正常功能对于椎间盘退变的防治起着决定性作用[9]。在肌肉及周围韧带的保护下,椎间盘通常处于静态平衡或者动态平衡,而一旦这种平衡被打破,脊柱稳定性丧失,就会导致椎间关节退变,椎间盘异常应力,进而引起椎间盘退变[10]。本研究通过破坏大鼠脊柱后方结构,构建脊柱失稳模型,结果显示,脊柱失稳能够明显诱导椎间盘退变,终板软骨细胞外基质的主要成分COL-2A 以及ACAN 的合成减少;因此,我们认为脊柱失稳引起的力学失衡导致终板软骨退变,终板营养途径障碍诱导椎间盘退变,改善终板软骨的功能可延缓椎间盘退变。
线粒体是真核动物细胞进行生物氧化和能量转换的主要场所,线粒体功能障碍与软骨细胞退变密切相关。线粒体受损以及功能障碍导致细胞内活性氧及氧化应激水平增高,进而影响软骨细胞合成及分解代谢,最终导致软骨细胞退变[11]。因此,适时清除老化和损伤的线粒体对于软骨细胞功能维持具有重要的作用。线粒体自噬能够选择性地清除细胞内受损或功能障碍的线粒体,降低细胞内活性氧水平,从而维持软骨细胞的内环境及功能稳定[12]。有研究表明,PINK1/Parkin 信号通路介导的线粒体自噬水平降低能够抑制关节软骨合成代谢,从而促进关节软骨退变[13]。然而,在力学失衡诱导的终板软骨退变过程中线粒体自噬水平的变化及作用机制目前仍不清楚。本研究结果显示,脊柱失稳模型中终板软骨细胞中PINK1/Parkin 信号通路被抑制,线粒体膜蛋白Tomm20 和Timm23水平增高,说明线粒体自噬水平降低导致线粒体蛋白降解减少;在脊柱失稳模型中给予线粒体自噬激动剂CCCP 激活线粒体自噬后,终板软骨中COL-2A及ACAN 表达升高,终板软骨及椎间盘的退变均明显减轻。由此可见,脊柱失稳导致的异常应力能够通过抑制线粒体自噬影响终板软骨细胞合成及分解代谢,进而诱导终板软骨退变,但异常应力刺激调控PINK1/Parkin 信号通路介导线粒体自噬的具体机制仍待进一步研究。
猜你喜欢
颈腰痛杂志(2023年2期)2023-05-05
海洋通报(2021年1期)2021-07-23
生物学通报(2021年4期)2021-03-16
云南医药(2019年3期)2019-07-25
中国骨与关节杂志(2019年6期)2019-01-03
中国中西医结合影像学杂志(2018年6期)2018-12-17
川北医学院学报(2015年5期)2015-12-05
精准医学杂志(2015年5期)2015-05-01
癌变·畸变·突变(2014年1期)2014-03-01
中国中西医结合外科杂志(2013年3期)2013-03-11
